Преоптическая область
| Преоптическая область | |
|---|---|
 Преоптическая область обозначена буквой «ПО» слева и выделена синим цветом. | |
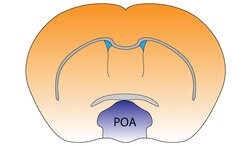 Преоптическая область мозга мыши | |
| Подробности | |
| Идентификаторы | |
| латинский | преоптическая область |
| Акроним(ы) | доверенность на доступ |
| МеШ | D011301 |
| Нейроимена | 377 |
| НейроЛекс ID | бирнлекс_1706 |
| ТА98 | A14.1.08.407 |
| ТА2 | 5710 |
| ФМА | 62313 |
| Анатомические термины нейроанатомии | |
Преоптическая область — это область гипоталамуса . MeSH классифицирует его как часть переднего гипоталамуса. TA перечисляет четыре ядра в этой области (медиальное, срединное, латеральное и перивентрикулярное).
Функции
[ редактировать ]Преоптическая область отвечает за терморегуляцию и получает нервную стимуляцию от терморецепторов кожи, слизистых оболочек и самого гипоталамуса. [1]
Ядра
[ редактировать ]Срединное преоптическое ядро
[ редактировать ]Срединное преоптическое ядро расположено вдоль средней линии, значительно дорсально по отношению к трем другим преоптическим ядрам, по крайней мере, в мозге крабоедающих макак . Он охватывает верхнюю (дорсальную), переднюю и нижнюю (вентральную) поверхности передней спайки .
Срединное преоптическое ядро генерирует жажду . Употребление алкоголя снижает высвобождение норадреналина в срединном преоптическом ядре. [2]
Медиальное преоптическое ядро
[ редактировать ]Медиальное преоптическое ядро ограничено латерально латеральным преоптическим ядром, а медиально — преоптическим перивентрикулярным ядром. Он высвобождает гонадотропин-рилизинг гормон (ГнРГ), контролирует совокупление у самцов и имеет больший размер у самцов, чем у самок.
Родительское поведение
[ редактировать ]Медиальная преоптическая область (mPOA) участвует в родительской заботе как у мужчин, так и у женщин. У крыс окситоцин и вазопрессин связаны с поддержанием материнской заботы посредством местного высвобождения в mPOA и прилежащем ядре ложа терминальной полоски (BNST). Связывание рецепторов окситоцина и вазопрессина V1a увеличивается как в mPOA, так и в BNST у кормящих крыс по сравнению с контролем. [3] mPOA также имеет высокую плотность рецепторов эстрадиола , которые при активации могут вызвать у крыс-самцов поведение материнского типа. [4] Кроме того, mPOA имеет решающее значение для возникновения и выражения родительского поведения, о чем свидетельствует увеличение количества непосредственно раннего гена c-fos у опытных матерей или отцов крыс по сравнению с контрольной группой. Исследования также показали, что у отцов, когда они получают ультразвуковые сигналы или сигналы феромонов от своих партнеров, экспрессия их c-fos в mPOA еще больше увеличивается, что позволяет предположить, что отцовское поведение крыс опосредовано через mPOA, но активируется прямым взаимодействием с партнером. [5] Крупные поражения mPOA нарушают начало материнского поведения, строительство гнезда и извлечение детенышей, при этом боковые проекции особенно важны. [6]
Сексуальное поведение
[ редактировать ]mPOA имеет половой диморфизм, то есть различается по функциям у мужчин и женщин. Исследования у женщин изучали влияние mPOA на прекопулятивное и аппетитное поведение. Прекопулятивное поведение затрагивает несколько областей мозга, включая mPOA, а также медиальную миндалину (MA) и BNST. Исследования с использованием самок сирийских хомяков показали, что mPOA важен для предпочтения сексуального запаха. В то время как самки контрольной группы будут исследовать мужские запахи больше, чем женские, у женщин с двусторонним поражением mPOA (MPOA-X) не будет выявлено различий в предпочтениях запахов, но вагинальная ароматическая маркировка и лордоз останутся незатронутыми. [7] Аппетитное поведение самок крыс, включая прыжки, дротики и домогательства, было связано с передачей дофамина (DA) в mPOA. Поскольку аскорбиновая кислота (витамин С) усиливает передачу ДА по мезолимбическим и нигростриарным путям, было показано, что введение аскорбиновой кислоты в mPOA увеличивает аппетитное поведение по сравнению с контрольной группой. [8]
У самцов крыс mPOA влияет на завершающую фазу сексуального поведения и, возможно, на мотивацию, при этом поражения вызывают полную потерю копулятивного поведения. [9] И наоборот, электрическая стимуляция этой области вызывает у мужчин копулятивное поведение, что измеряется уменьшением латентного периода до эякуляции. [10] Более того, тестостерон, имплантированный в мПОА кастрированных самцов, полностью восстанавливает спаривание, пока ароматаза . не подавляется [11]
Вентролатеральное преоптическое ядро
[ редактировать ]Вентролатеральное преоптическое ядро , или промежуточное ядро, примыкает к медиальному преоптическому ядру. Он также опосредует начало медленного сна.
Преоптическое перивентрикулярное ядро
[ редактировать ]Преоптическое перивентрикулярное ядро расположено вдоль средней линии и медиальнее медиального преоптического ядра.
Ссылки
[ редактировать ]- ^ Мартин Дж. Х. (2003). Нейроанатомия: Текст и атлас . МакГроу-Хилл Профессионал.
- ^ Миякубо Х., Ямамото К., Хатакенака С., Хаяси Ю., Танака Дж. (октябрь 2003 г.). «Питьё снижает выброс норадреналина в срединной преоптической области, вызванный гиповолемией у крыс». Поведенческие исследования мозга . 145 (1–2): 1–5. дои : 10.1016/S0166-4328(03)00102-5 . ПМИД 14529799 . S2CID 46018164 .
- ^ Бош О.Дж., Пфёрч Дж., Байдербек Д.И., Ландграф Р., Нойманн И.Д. (май 2010 г.). «Материнское поведение связано с высвобождением вазопрессина в медиальной преоптической области и ядре ложа терминальной полоски у крыс». Журнал нейроэндокринологии . 22 (5): 420–9. дои : 10.1111/j.1365-2826.2010.01984.x . ПМИД 20163514 . S2CID 5286826 .
- ^ Розенблатт Дж. С., Сеус К. (февраль 1998 г.). «Эстрогеновые имплантаты в медиальной преоптической области стимулируют материнское поведение у самцов крыс». Гормоны и поведение . 33 (1): 23–30. дои : 10.1006/hbeh.1997.1430 . ПМИД 9571010 . S2CID 45305790 .
- ^ Чжун Дж., Лян М., Актер С., Хигасида С., Цудзи Т., Хигасида Х. (сентябрь 2014 г.). «Экспрессия c-Fos в мозге отцовской мыши, индуцированная коммуникативным взаимодействием с товарищами по материнской линии» . Молекулярный мозг . 7 (1): 66. дои : 10.1186/s13041-014-0066-x . ПМЦ 4172782 . ПМИД 25208928 .
- ^ Нуман М (1988). «Нейральные основы материнского поведения крысы». Психонейроэндокринология . 13 (1–2): 47–62. дои : 10.1016/0306-4530(88)90006-6 . ПМИД 2897700 . S2CID 5928669 .
- ^ Мартинес Л.А., Петрулис А. (апрель 2013 г.). «У самок сирийских хомяков медиальная преоптическая область необходима для предпочтения сексуальных запахов, но не для сексуального возбуждения» . Гормоны и поведение . 63 (4): 606–14. дои : 10.1016/j.yhbeh.2013.02.003 . ПМЦ 3633686 . ПМИД 23415835 .
- ^ Грэм, доктор медицинских наук, Пфаус Дж.Г. (октябрь 2013 г.). «Вливание аскорбиновой кислоты в медиальную преоптическую область способствует аппетитному сексуальному поведению самок крыс». Физиология и поведение . 122 (1): 140–6. дои : 10.1016/j.physbeh.2013.09.008 . ПМИД 24064109 . S2CID 9515707 .
- ^ Паредес Р.Г. (июль 2003 г.). «Медиальная преоптическая область/передний гипоталамус и сексуальная мотивация». Скандинавский журнал психологии . 44 (3): 203–12. дои : 10.1111/1467-9450.00337 . ПМИД 12914583 .
- ^ Малсбери CW (декабрь 1971 г.). «Облегчение копулятивного поведения самцов крыс путем электрической стимуляции медиальной преоптической области». Физиология и поведение . 7 (6): 797–805. дои : 10.1016/0031-9384(71)90042-4 . ПМИД 5134017 .
- ^ Кристенсен Л.В., Клеменс Л.Г. (декабрь 1975 г.). «Блокада тестостерон-индуцированного монтажного поведения у самцов крыс с помощью внутричерепного применения ингибитора ароматизации андрост-1,4,6-триен-3,17-диона». Эндокринология . 97 (6): 1545–51. дои : 10.1210/endo-97-6-1545 . ПМИД 1204576 .