Фитохром
| Фитохром | |||
|---|---|---|---|
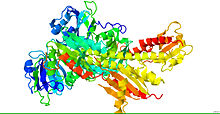 Кристаллическая структура фитохрома. [1] | |||
| Идентификаторы | |||
| Символ | Фитохром | ||
| Пфам | PF00360 | ||
| ИнтерПро | ИПР013515 | ||
| |||

Фитохромы представляют собой класс белков-фоторецепторов, обнаруженных в растениях , бактериях и грибах . Они реагируют на свет в красной и дальней красной областях видимого спектра и могут быть классифицированы как Тип I, который активируется дальним красным светом, или Тип II, который активируется красным светом. [2] Недавние достижения показали, что фитохромы также действуют как датчики температуры, поскольку более высокие температуры усиливают их дезактивацию. [3] Все эти факторы способствуют способности растения прорастать .
Фитохромы контролируют многие аспекты развития растений. Они регулируют прорастание семян , удлинение проростков , (фотобластия), синтез хлорофилла размер, форму, количество и движение листьев и сроки цветения взрослых растений. Фитохромы широко экспрессируются во многих тканях и на разных стадиях развития. [2]
Другие фоторецепторы растений включают криптохромы и фототропины , которые реагируют на синий и ультрафиолетовый свет-А, а также UVR8 , который чувствителен к ультрафиолетовому свету-В.
Структура
[ редактировать ]Фитохромы состоят из белка , ковалентно связанного со светочувствительным билина хромофором . [4] Белковая часть состоит из двух идентичных цепей (А и В). Каждая цепочка имеет домен PAS , домен GAF и домен PHY. Расположение доменов в фитохромах растений, бактерий и грибов сопоставимо, поскольку три N-концевых домена всегда представляют собой домены PAS, GAF и PHY. Однако С-концевые домены более расходятся. Домен PAS служит сенсором сигнала, а домен GAF отвечает за связывание с цГМФ, а также воспринимает световые сигналы. Вместе эти субъединицы образуют фитохромную область, которая регулирует физиологические изменения растений при изменении условий красного и дальнего красного света. У растений красный свет переводит фитохром в его биологически активную форму, а дальний красный свет переводит белок в его биологически неактивную форму.
Изоформы и состояния
[ редактировать ]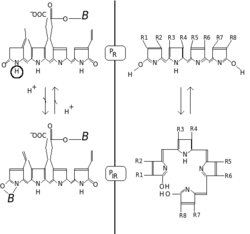
Фитохромы характеризуются красной/дальнекрасной фотохромностью . Фотохромные пигменты меняют свой «цвет» (свойства спектрального поглощения) при поглощении света. В случае фитохрома основным состоянием является Pr , причем буква r указывает на то, что он особенно сильно поглощает красный свет. Максимум поглощения представляет собой острый пик 650–670 нм, поэтому концентрированные растворы фитохромов кажутся человеческому глазу бирюзово-голубыми при просмотре в белом свете. Но как только красный фотон поглощается, пигмент претерпевает быстрое конформационное изменение, образуя состояние P fr . Здесь fr указывает на то, что теперь дифференциально поглощается не красный, а дальний красный (также называемый «ближним инфракрасным»; 705–740 нм). Этот сдвиг в поглощении виден человеческому глазу как немного более зеленоватый цвет. Когда P fr поглощает дальний красный свет, он преобразуется обратно в P r . Следовательно, красный свет создает P fr , дальний красный свет создает P r . У растений по крайней мере P fr представляет собой физиологически активное или «сигнальное» состояние.
Влияние фитохромов на фототропизм
[ редактировать ]Фитохромы также обладают способностью чувствовать свет, что заставляет растение расти навстречу ему. Это называется фототропизмом . [7] Джаноуди и его коллеги хотели посмотреть, какой тип фитохрома отвечает за возникновение фототропизма, и провели серию экспериментов. Они обнаружили, что синий свет вызывает у растения Arabidopsis thaliana фототропную реакцию; эта кривизна усиливается с добавлением красного света. [7] Они также обнаружили, что в растении присутствовало пять различных фитохромов, в то время как у некоторых мутантов, которые не функционировали должным образом, наблюдался недостаток фитохромов. [7] Два из этих мутантных вариантов были очень важны для этого исследования: phyA-101 и phyB-1. [7] Это мутанты фитохрома А и В соответственно. Нормально функциональный фитохром А вызывает чувствительность к дальнему красному свету и регулирует выражение кривизны по отношению к свету, тогда как фитохром B более чувствителен к красному свету. [7]
Эксперимент включал форму дикого типа Arabidopsis, phyA-101 (нулевой мутант по фитохрому A (phyA)), phyB-1 (мутант с дефицитом фитохрома B). [7] Затем их подвергали воздействию белого света в качестве контрольного синего и красного света при различных плотностях света и измеряли кривизну. [7] Было установлено, что для достижения фенотипа , подобного фенотипу дикого типа, phyA-101 должен быть подвергнут воздействию на четыре порядка большей величины или около 100 мкмоль·м. −2 флюенс. [7] Однако плотность энергии, которая заставляет phyB-1 проявлять ту же кривизну, что и у дикого типа, идентична флюенсу дикого типа. [7] Было обнаружено, что фитохром, который экспрессировал больше, чем обычно, количества фитохрома А, по мере увеличения флюенса, кривизна также увеличивалась до 10 мкмоль-м. −2 кривизна была аналогична дикому типу. [7] Фитохром, экспрессирующий больше, чем обычно, количество фитохрома B, демонстрировал кривизну, аналогичную кривизне дикого типа, при различных флюенсах красного света вплоть до флюенса 100 мкмоль-м. −2 при флюенсах выше этой кривизны была намного выше, чем у дикого типа. [7]
Таким образом, в результате эксперимента было обнаружено, что на кривизну влияет не только фитохром А, но и другой фитохром, поскольку мутант не так уж далек от дикого типа, а phyA вообще не экспрессируется. [7] Это приводит к выводу, что за фототропизм должны отвечать две фазы. Они определили, что ответ происходит как при низких, так и при высоких флюенсах. [7] Это связано с тем, что для phyA-101 порог кривизны возникает при более высоких значениях флюенса, но искривление также возникает при низких значениях флюенса. [7] Поскольку порог мутанта возникает при высоких значениях флюенса, было установлено, что фитохром А не отвечает за искривление при высоких значениях флюенса. [7] Поскольку мутант по фитохрому B продемонстрировал реакцию, аналогичную реакции дикого типа, был сделан вывод, что фитохром B не необходим для усиления воздействия низкой или высокой плотности энергии. [7] Было предсказано, что мутанты, которые сверхэкспрессируют фитохромы А и В, будут более чувствительными. Однако показано, что чрезмерная экспрессия phy A на самом деле не влияет на кривизну, поэтому в диком типе фитохрома достаточно для достижения максимальной кривизны. [7] Для мутанта со сверхэкспрессией фитохрома B более высокая кривизна, чем обычно, при более высоких плотностях света указывает на то, что фитохром B контролирует кривизну при высоких плотностях света. [7] В целом они пришли к выводу, что фитохром А контролирует искривление при низкой плотности потока света. [7]
Влияние фитохрома на рост корней
[ редактировать ]Фитохромы также могут влиять на рост корней. Хорошо известно, что гравитропизм является основным тропизмом корней. Однако недавнее исследование показало, что фототропизм также играет роль. Положительный фототропизм, вызываемый красным светом, недавно был зарегистрирован в эксперименте, в котором использовался арабидопсис, чтобы проверить, какие участки растения оказывают наибольшее влияние на положительную фототропную реакцию. Экспериментаторы использовали аппарат, который позволял регулировать высоту вершины корня до нуля градусов, чтобы гравитропизм не мог быть конкурирующим фактором. При помещении на красный свет корни арабидопсиса искривились на 30–40 градусов. Это показало положительную фототропную реакцию на красный свет. Затем они хотели точно определить, где именно на растении поступает свет. Когда корни были закрыты, искривление корней под воздействием красного света практически не наблюдалось. Напротив, когда побеги были покрыты, наблюдалась положительная фототропная реакция на красный свет. Это доказывает, что в боковых корнях происходит светоощущение. Для дальнейшего сбора информации о фитохромах, участвующих в этой активности, мутанты фитохромов A, B, D и E, а также корни WT подвергались воздействию красного света. Мутанты фитохромов А и В были серьезно повреждены. Существенной разницы в ответе не было. phyD и phyE по сравнению с диким типом, доказывая, что phyA и phyB ответственны за положительный фототропизм в корнях.
Биохимия
[ редактировать ]Химически фитохром состоит из хромофора , одной молекулы билина, состоящей из открытой цепи из четырех пиррольных колец, ковалентно связанных с фрагментом белка через высококонсервативную аминокислоту цистеин. Именно хромофор поглощает свет и в результате меняет конформацию билина, а затем и присоединенного белка, переводя его из одного состояния или изоформы в другое.
Фитохромный хромофор обычно представляет собой фитохромомобилин и тесно связан с фикоцианобилином (хромофором фикобилипротеинов) используемых . цианобактериями (на структуру которого также и красными водорослями для захвата света для фотосинтеза ) и желчного пигмента билирубина влияет воздействие света, что используется при фототерапии новорожденных с желтухой ).Термин «били» во всех этих названиях относится к желчи. Билины образуются из закрытого тетрапиррольного кольца гема в результате окислительной реакции, катализируемой гем-оксигеназой, с образованием характерной открытой цепи. Хлорофилл и гем ( гем ) имеют общего предшественника в форме протопорфирина IX и имеют одинаковую характерную замкнутую тетрапиррольную кольцевую структуру. В отличие от билинов гем и хлорофилл несут в центре кольца атом металла — железа или магния соответственно. [8]
Состояние P fr передает сигнал другим биологическим системам клетки, например, механизмам, ответственным за экспрессию генов . Хотя этот механизм почти наверняка является биохимическим процессом, он до сих пор остается предметом многочисленных дискуссий. Известно, что хотя фитохромы синтезируются в цитозоле P r и там локализуется форма , форма P fr , генерируемая световым освещением, транслоцируется в ядро клетки . Это подразумевает роль фитохрома в контроле экспрессии генов, и известно, что многие гены регулируются фитохромом, но точный механизм еще предстоит полностью раскрыть. Было высказано предположение, что фитохром в форме P fr может действовать как киназа , и было продемонстрировано, что фитохром в форме P fr может напрямую взаимодействовать с факторами транскрипции . [9]
Открытие
[ редактировать ]Фитохромный пигмент был обнаружен Стерлингом Хендриксом и Гарри Бортвиком в Центре сельскохозяйственных исследований Министерства сельского хозяйства США-ARS в Белтсвилле в Мэриленде в период с конца 1940-х до начала 1960-х годов. Используя спектрограф, построенный из заимствованных и полученных в результате войны деталей, они обнаружили, что красный свет очень эффективен для стимулирования прорастания или запуска реакции цветения. Реакция на красный свет была обратима под действием дальнего красного света, что указывает на наличие фотообратимого пигмента.
Пигмент фитохром был идентифицирован с помощью спектрофотометра в 1959 году биофизиком Уорреном Батлером и биохимиком Гарольдом Сигельманом . Батлер также придумал название «фитохром».
В 1983 году лаборатории Питера Куэйла и Кларка Лагариаса сообщили о химической очистке интактной молекулы фитохрома, а в 1985 году первую последовательность гена Говард Херши и Питер Куэйл опубликовали фитохрома. К 1989 году молекулярная генетика и работа с моноклональными антителами установили, что существовало более одного типа фитохромов; например, было показано, что растение гороха имеет по крайней мере два типа фитохромов (тогда называемые типом I (обнаруженным преимущественно в темных сеянцах) и типом II (преобладающим в зеленых растениях)). теперь известно Благодаря секвенированию генома , что арабидопсис имеет пять генов фитохромов (PHYA-E), а у риса их только три (PHYA-C). Хотя это, вероятно, отражает состояние некоторых дву- и однодольных растений, многие растения являются полиплоидными . Следовательно, кукуруза , например, имеет шесть фитохромов — phyA1, phyA2, phyB1, phyB2, phyC1 и phyC2. Хотя все эти фитохромы имеют существенно разные белковые компоненты, все они используют фитохромомобилин в качестве светопоглощающего хромофора. Фитохром А или фиА быстро разлагается в форме Pfr — гораздо сильнее, чем другие члены семейства. В конце 1980-х годов лаборатория Виерстра показала, что phyA разлагается под действием убиквитиновой системы, первой естественной мишени системы, идентифицированной у эукариот.
В 1996 году Дэвид Кехо и Артур Гроссман из Института Карнеги при Стэнфордском университете идентифицировали белки в нитчатой цианобактерии Fremyella diplosiphon, названные RcaE, с фитохромом, аналогичным растительному, который контролирует красно-зеленый фотообратимый ответ, называемый хроматической акклимацией, и идентифицировали ген в секвенированной опубликовали геном цианобактерии Synechocystis , более похожий на геном растительного фитохрома. Это было первое свидетельство существования фитохромов за пределами царства растений. Джон Хьюз из Берлина и Кларк Лагариас из Калифорнийского университета в Дэвисе впоследствии показали, что этот ген Synechocystis действительно кодирует настоящий фитохром (названный Cph1) в том смысле, что это красный/дальнекрасный обратимый хромопротеин. Предположительно, фитохромы растений произошли от предкового фитохрома цианобактерий, возможно, путем миграции генов из хлоропласта в ядро. Впоследствии фитохромы были обнаружены у других прокариот, включая Deinococcus radiodurans и Agrobacterium tumefaciens. . У Deinococcus фитохром регулирует выработку светозащитных пигментов, однако у Synechocystis и Agrobacterium биологическая функция этих пигментов пока неизвестна.
В 2005 году лаборатории Виерстра и Форест в Университете Висконсина опубликовали трехмерную структуру усеченного фитохрома Deinococcus (домены PAS/GAF). Эта статья показала, что белковая цепь образует узел — весьма необычную структуру для белка. В 2008 году две группы из Эссена и Хьюза в Германии и Янга и Моффата в США опубликовали трехмерные структуры всей фотосенсорной области. Одна из структур относилась к Synechocystis sp. (штамм PCC 6803) фитохрома в Pr и другой для фитохрома Pseudomonas aeruginosa в состоянии P fr . Структуры показали, что консервативная часть домена PHY, так называемый язык PHY, принимает различные складки. В 2014 году Такала и др. подтвердили, что рефолдинг происходит даже для одного и того же фитохрома (из Deinococcus ) в зависимости от условий освещения.
Генная инженерия
[ редактировать ]Примерно в 1989 году нескольким лабораториям удалось получить трансгенные растения , которые производили повышенное количество различных фитохромов ( сверхэкспрессия ). Во всех случаях полученные растения имели заметно короткие стебли и темно-зеленые листья. Гарри Смит и его коллеги из Лестерского университета в Англии показали, что, повышая уровень экспрессии фитохрома А (который реагирует на дальний красный свет), избегания тени . можно изменить реакцию [10] В результате растения могут тратить меньше энергии на то, чтобы вырасти как можно выше, и иметь больше ресурсов для выращивания семян и расширения своей корневой системы. Это может иметь много практических преимуществ: например, травинки, которые будут расти медленнее, чем обычная трава, не будут требовать столь частого скашивания, или сельскохозяйственные растения могут передавать больше энергии зерну вместо того, чтобы расти выше.
В 2002 году светоиндуцированное взаимодействие между растительным фитохромом и фактором взаимодействия с фитохромами (PIF) было использовано для контроля транскрипции генов у дрожжей. Это был первый пример использования фотобелков другого организма для управления биохимическим путем. [11]
Ссылки
[ редактировать ]- ^ PDB : 3G6O ; Ян Икс, Кук Дж, Моффат К (2009). «Кристаллическая структура фитохрома бактериального фитохрома PaBphP P. aeruginosa, мутанта фотосенсорного основного домена Q188L» . Учеб. Натл. акад. наук. США . 106 (37): 15639–15644. дои : 10.1073/pnas.0902178106 . ПМЦ 2747172 . ПМИД 19720999 .
- ^ Jump up to: а б Ли Дж, Ли Джи, Ван Х, Ван Дэн Х (2011). «Фитохромные сигнальные механизмы» . Книга «Арабидопсис» . 9 : e0148. дои : 10.1199/tab.0148 . ПМК 3268501 . ПМИД 22303272 .
- ^ Холлидей, Карен Дж.; Дэвис, Сет Дж. (2016). «Светочувствительные фитохромы чувствуют тепло» (PDF) . Наука . 354 (6314): 832–833. Бибкод : 2016Sci...354..832H . дои : 10.1126/science.aaj1918 . ПМИД 27856866 . S2CID 42594849 .
- ^ Шаррок РА (2008). Суперсемейство фоторецепторов фитохрома красного/дальнего красного цвета. Геномная биология, 9(8), 230. doi:10.1186/gb-2008-9-8-230 PMC 2575506.
- ^ Бритц С.Дж., Галстон А.В. (февраль 1983 г.). «Физиология движений в стеблях проростков Pisum sativum L. cv Alaska: III. Фототропизм по отношению к гравитропизму, нутации и росту» . Физиол растений . 71 (2): 313–318. дои : 10.1104/стр.71.2.313 . ПМЦ 1066031 . ПМИД 16662824 .
- ^ Уокер Т.С., Бэйли Дж.Л. (апрель 1968 г.). «Две спектрально разные формы фитохромного хромофора, извлеченные из этиолированных проростков овса» . Биохим Дж . 107 (4): 603–605. дои : 10.1042/bj1070603 . ПМК 1198706 . ПМИД 5660640 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с Абдул-Кадер, Джаноуди (1977). «Множественные фитохромы участвуют в индуцированном красным светом усилении первого положительного фототропизма у Arabidopsis thaliana» (PDF) . www.plantpsyol.org .
- ^ Маусет, Джеймс Д. (2003). Ботаника: Введение в биологию растений (3-е изд.). Садбери, Массачусетс: Джонс и Бартлетт Обучение. стр. 422–427. ISBN 978-0-7637-2134-3 .
- ^ Шин, А-Янг; Хан, Юн-Джонг; Пэк, Аён; Ан, Тэхо; Ким, Су Ён; Нгуен, Тай Сон; Сын Минки; Ли, Гын У; Шен, Ю (13 мая 2016 г.). «Доказательства того, что фитохром действует как протеинкиназа в передаче световых сигналов растениям» . Природные коммуникации . 7 (1): 11545. Бибкод : 2016NatCo...711545S . дои : 10.1038/ncomms11545 . ISSN 2041-1723 . ПМК 4869175 . ПМИД 27173885 .
- ^ Робсон, П.Р.Х., МакКормак, А.С., Ирвин, А.С. и Смит, Х. Генная инженерия индекса урожая табака посредством сверхэкспрессии гена фитохрома. Природные биотехнологии. 14, 995–998 (1996).
- ^ Симидзу-Сато С., Хук Э., Тепперман Дж.М., Куэйл П.Х. (октябрь 2002 г.). «Система промотора генов с переключателем света». Природная биотехнология . 20 (10): 1041–4. дои : 10.1038/nbt734 . ПМИД 12219076 . S2CID 24914960 .
Источники
[ редактировать ]- Лия Х, Чжанб Дж, Виерстра Р.Д., Лия Х (2010). «Четвертичная организация димера фитохрома, выявленная методом криоэлектронной микроскопии» . ПНАС . 107 (24): 10872–10877. Бибкод : 2010PNAS..10710872L . дои : 10.1073/pnas.1001908107 . ПМК 2890762 . ПМИД 20534495 .
- «Фантастическое срабатывание выключателя света» , Джим Де Кватро, 1991 год.
- «Хронометраж природы» , Кит Смит, 2004 г.
- Терри и Джерри Одезирк. Биология: Жизнь на Земле.
- Линда Сейдж. Пигмент воображения: история исследования фитохрома. Академическая Пресса 1992. ISBN 0-12-614445-1
- Гурурани, Маянк Ананд, Марккандан Ганесан и Сонг Пилл-Сун. «Фотобиотехнология как инструмент улучшения агрономических качеств сельскохозяйственных культур». Достижения биотехнологии (2014).
| Базы данных органов управления : Национальные |
|---|