Нейронное кодирование
Нейронное кодирование (или нейронное представление ) — это область нейробиологии, занимающаяся характеристикой гипотетических взаимосвязей между стимулом и реакциями нейронов, а также взаимосвязей между электрической активностью нейронов в ансамбле . [1] [2] На основе теории, чтоСенсорная и другая информация представлена в мозге сетями нейронов . Считается, что нейроны могут кодировать как цифровую , так и аналоговую информацию. [3]
Обзор
[ редактировать ]Нейроны обладают необычной для клеток организма способностью быстро распространять сигналы на большие расстояния, генерируя характерные электрические импульсы, называемые потенциалами действия : скачки напряжения, которые могут перемещаться по аксонам. Сенсорные нейроны изменяют свою активность, запуская последовательности потенциалов действия в различных временных рамках в присутствии внешних сенсорных раздражителей, таких как свет , звук , вкус , запах и прикосновение . Информация о стимуле кодируется в этом образце потенциалов действия и передается в мозг и вокруг него. Помимо этого, специализированные нейроны, например нейроны сетчатки, могут передавать больше информации посредством градуированных потенциалов . Они отличаются от потенциалов действия, поскольку информация о силе стимула напрямую коррелирует с силой сигнала нейронов. Сигнал затухает гораздо быстрее для градуированных потенциалов, что требует коротких расстояний между нейронами и высокой плотности нейронов. Преимущество градуированных потенциалов заключается в более высоких скоростях передачи информации, способных кодировать больше состояний (т.е. более высокую точность), чем импульсные нейроны. [4]
Хотя потенциалы действия могут несколько различаться по продолжительности, амплитуде и форме, в исследованиях нейронного кодирования их обычно рассматривают как идентичные стереотипные события. Если игнорировать кратковременную продолжительность потенциала действия (около 1 мс), последовательность потенциалов действия, или серия спайков, может быть охарактеризована просто серией точечных событий во времени по принципу «все или ничего» . [5] Длины межспайковых интервалов ( ISI ) между двумя последовательными спайками в серии спайков часто различаются, по-видимому, случайным образом. [6] Изучение нейронного кодирования включает измерение и характеристику того, как атрибуты стимула, такие как интенсивность света или звука, или двигательные действия, такие как направление движения руки, представлены потенциалами действия или спайками нейронов. Для описания и анализа возбуждения нейронов статистические методы , методы теории вероятностей и стохастических точечных процессов широко применяются .
С развитием крупномасштабных технологий нейронной записи и декодирования исследователи начали взламывать нейронный код и уже впервые смогли взглянуть на нейронный код в реальном времени, когда память формируется и вызывается в гиппокампе, области мозга, которая, как известно, играть центральную роль в формировании памяти. [7] [8] [9] Нейробиологи инициировали несколько крупномасштабных проектов по расшифровке мозга. [10] [11]
Кодирование и декодирование
[ редактировать ]Связь между стимулом и реакцией можно изучать с двух противоположных точек зрения. Нейронное кодирование относится к карте от стимула к реакции. Основная задача — понять, как нейроны реагируют на широкий спектр стимулов, и построить модели, которые пытаются предсказать реакцию на другие стимулы. Нейронное декодирование относится к обратной карте ответа на стимул, и задача состоит в том, чтобы реконструировать стимул или определенные аспекты этого стимула на основе последовательностей спайков, которые он вызывает.
Гипотетические схемы кодирования
[ редактировать ]Последовательность или «последовательность» импульсов может содержать информацию, основанную на различных схемах кодирования. В некоторых нейронах сила реакции постсинаптического партнера может зависеть исключительно от «частоты срабатывания», среднего количества импульсов в единицу времени («код скорости»). С другой стороны, сложный « временной код » основан на точном времени одиночных всплесков. Они могут быть привязаны к внешнему стимулу, например зрительному. [12] и слуховой системой или генерироваться непосредственно нейронными цепями. [13]
Используют ли нейроны частотное или временное кодирование, это тема интенсивных споров в нейробиологическом сообществе, хотя четкого определения того, что означают эти термины, не существует. [14]
Традиционный вид: код тарифа
[ редактировать ]Модель кодирования скорости нейронной связи утверждает, что по мере увеличения интенсивности стимула частота или скорость потенциалов действия , или «спайковых импульсов», увеличивается. Кодирование скорости иногда называют частотным кодированием.
Кодирование скорости — это традиционная схема кодирования, предполагающая, что большая часть, если не вся, информация о стимуле содержится в частоте срабатывания нейрона. Поскольку последовательность потенциалов действия, генерируемых данным стимулом, варьируется от испытания к испытанию, реакции нейронов обычно обрабатываются статистически или вероятностно. Они могут характеризоваться скорострельностью, а не конкретными последовательностями всплесков. В большинстве сенсорных систем частота импульсов увеличивается, как правило, нелинейно с увеличением интенсивности стимула. [15] Согласно предположению о кодировании скорости, любая информация, возможно закодированная во временной структуре последовательности импульсов, игнорируется. Следовательно, кодирование скорости неэффективно, но очень устойчиво к « шуму » ISI. [6]
Во время кодирования скорости очень важен точный расчет скорострельности. Фактически, термин «скорость срабатывания» имеет несколько разных определений, которые относятся к различным процедурам усреднения, таким как среднее значение по времени (частота как количество спайков одного нейрона) или среднее значение по нескольким повторениям (частота PSTH) эксперимент.
При кодировании скорости обучение основано на зависимых от активности модификациях синаптического веса.
Кодирование скорости было первоначально показано Эдгаром Адрианом и Ингве Зоттерманом в 1926 году. [16] подвешивались разные веса В этом простом эксперименте к мышце . По мере увеличения веса стимула количество импульсов, регистрируемых сенсорными нервами, иннервирующими мышцу, также увеличивалось. На основании этих оригинальных экспериментов Адриан и Зоттерман пришли к выводу, что потенциалы действия представляют собой унитарные события и что основой большинства межнейронных коммуникаций является частота событий, а не величина отдельного события.
В последующие десятилетия измерение скорости стрельбы стало стандартным инструментом для описания свойств всех типов сенсорных или корковых нейронов, отчасти из-за относительной простоты экспериментального измерения скорости. Однако этот подход игнорирует всю информацию, которая может содержаться в точном времени всплесков. В последние годы появляется все больше и больше экспериментальных данных, свидетельствующих о том, что простая концепция скорострельности, основанная на временном усреднении, может быть слишком упрощенной для описания активности мозга. [6]
Скорость подсчета пиков (средняя за период времени)
[ редактировать ]Скорость подсчета пиков, также называемая средним временным значением, получается путем подсчета количества пиков, возникающих во время испытания, и деления его на продолжительность испытания. [14] Длина Т временного окна задается экспериментатором и зависит от типа нейрона, регистрируемого от и до стимула. На практике, чтобы получить разумные средние значения, в пределах временного окна должно произойти несколько всплесков. Типичные значения: T = 100 мс или T = 500 мс, но продолжительность также может быть больше или короче ( глава 1.5 в учебнике «Модели импульсных нейронов»). [14] ).
Частоту подсчета спайков можно определить по результатам одного испытания, но за счет потери всего временного разрешения изменений нейронной реакции в ходе испытания. Временное усреднение может хорошо работать в тех случаях, когда стимул постоянен или медленно меняется и не требует быстрой реакции организма — и именно такая ситуация обычно встречается в протоколах экспериментов. Однако реальные данные вряд ли являются стационарными, а часто меняются в быстром временном масштабе. Например, даже при просмотре статического изображения люди совершают саккады — быстрые изменения направления взгляда. Таким образом, изображение, проецируемое на фоторецепторы сетчатки , меняется каждые несколько сотен миллисекунд ( глава 1.5 в [14] )
Несмотря на свои недостатки, концепция кода скорости счета импульсов широко используется не только в экспериментах, но и в моделях нейронных сетей . Это привело к идее, что нейрон преобразует информацию об одной входной переменной (силе стимула) в одну непрерывную выходную переменную (скорость срабатывания).
Появляется все больше доказательств того, что, по крайней мере, в нейронах Пуркинье информация кодируется не только при срабатывании, но также во времени и продолжительности периодов покоя и покоя. [17] [18] Исследования клеток сетчатки также показали, что информация кодируется не только в частоте импульсов, но и во времени импульсов. [19] В более общем плане, всякий раз, когда требуется быстрая реакция организма, скорость срабатывания, определяемая как количество импульсов в течение нескольких сотен миллисекунд, просто слишком медленна. [14]
Зависящая от времени скорострельность (в среднем по нескольким испытаниям)
[ редактировать ]Зависящая от времени скорость стрельбы определяется как среднее количество всплесков (усредненное по испытаниям), появляющихся в течение короткого интервала между моментами t и t+Δt, деленное на продолжительность интервала. [14] Он работает как для стационарных, так и для зависящих от времени стимулов. Чтобы экспериментально измерить скорость срабатывания, зависящую от времени, экспериментатор записывает сигналы нейрона, стимулируя его некоторой входной последовательностью. Одна и та же последовательность стимуляции повторяется несколько раз, и реакция нейрона отображается на гистограмме перистимул-время (PSTH). Время t измеряется относительно начала последовательности стимуляции. Δt должно быть достаточно большим (обычно в диапазоне одной или нескольких миллисекунд), чтобы в интервале было достаточное количество выбросов для получения надежной оценки среднего значения. Количество появлений спайков n K (t;t+Δt), суммированное по всем повторениям эксперимента, деленное на количество K повторений, является мерой типичной активности нейрона между временем t и t+Δt. Дальнейшее деление на длину интервала Δt дает зависящую от времени частоту импульсов r(t) нейрона, которая эквивалентна плотности спайков PSTH ( глава 1.5 в [14] ).
Для достаточно малого Δt r(t)Δt — это среднее количество всплесков, происходящих между моментами t и t+Δt за несколько испытаний. Если Δt мало, в каждом конкретном испытании никогда не будет более одного всплеска в интервале между t и t+Δt. Это означает, что r(t)Δt — это также доля испытаний, в которых между этими моментами произошел всплеск. Эквивалентно, r(t)Δt — это вероятность того, что всплеск произойдет в течение этого интервала времени.
В качестве экспериментальной процедуры измерение скорости срабатывания, зависящее от времени, является полезным методом оценки активности нейронов, особенно в случае стимулов, зависящих от времени. Очевидная проблема этого подхода заключается в том, что он не может быть схемой кодирования, используемой нейронами мозга. Нейроны не могут дождаться повторного появления одинаковых стимулов, прежде чем сгенерировать ответ. [14]
Тем не менее, экспериментальное измерение скорости срабатывания, зависящее от времени, может иметь смысл, если существуют большие популяции независимых нейронов, которые получают один и тот же стимул. Вместо записи популяции из N нейронов за один прогон экспериментально проще записать данные с одного нейрона и усреднить по N повторным прогонам. Таким образом, зависящее от времени кодирование скорости срабатывания основано на неявном предположении, что популяции нейронов существуют всегда.
Временное кодирование
[ редактировать ]Когда обнаруживается, что точное время всплесков или высокочастотные колебания скорости стрельбы несут информацию, нейронный код часто идентифицируется как временной код. [14] [20] Ряд исследований показал, что временное разрешение нейронного кода находится в миллисекундном масштабе времени, что указывает на то, что точное время всплеска является важным элементом нейронного кодирования. [3] [21] [19] Такие коды, которые обмениваются данными через время между импульсами, также называются кодами межимпульсных интервалов и были подтверждены недавними исследованиями. [22]
Нейроны демонстрируют высокочастотные колебания скорости стрельбы, которые могут быть шумом или нести информацию. Модели скоростного кодирования предполагают, что эти нарушения являются шумом, тогда как модели временного кодирования предполагают, что они кодируют информацию. Если бы нервная система использовала коды скорости только для передачи информации, более последовательная и регулярная скорость стрельбы была бы эволюционно выгодной, и нейроны использовали бы этот код по сравнению с другими, менее надежными вариантами. [23] Временное кодирование дает альтернативное объяснение «шуму», предполагая, что он на самом деле кодирует информацию и влияет на нейронную обработку. Чтобы смоделировать эту идею, для обозначения всплесков можно использовать двоичные символы: 1 — всплеск, 0 — отсутствие всплеска. Временное кодирование позволяет последовательности 000111000111 означать нечто иное, чем 001100110011, хотя средняя скорость стрельбы одинакова для обеих последовательностей и составляет 6 пиков/10 мс. [24] До недавнего времени ученые уделяли наибольшее внимание кодированию скорости как объяснению паттернов постсинаптического потенциала . Однако функции мозга более точны во времени, чем позволяет использование только кодирования скорости. [19] Другими словами, важная информация может быть потеряна из-за неспособности кода тарифа уловить всю доступную информацию о пакете всплесков. Кроме того, реакции на схожие (но не идентичные) стимулы достаточно различаются, что позволяет предположить, что отдельные образцы спайков содержат больший объем информации, чем можно включить в код скорости. [25]
Временные коды (также называемые кодами спайков) [14] ), используют те особенности пиковой активности, которые не могут быть описаны скоростью стрельбы. Например, время до первого всплеска после начала стимула, фаза срабатывания по отношению к фоновым колебаниям, характеристики, основанные на втором и более высоких статистических моментах ISI распределения вероятностей , случайность всплеска или точно рассчитанные по времени группы всплесков. ( временные шаблоны ) являются кандидатами на временные коды. [26] Поскольку в нервной системе нет абсолютной привязки ко времени, информация передается либо в терминах относительного времени возникновения спайков в популяции нейронов (временные паттерны), либо в отношении продолжающихся колебаний мозга (фаза возбуждения). [3] [6] Один из способов декодирования временных кодов при наличии нейронных колебаний заключается в том, что спайки, возникающие в определенных фазах колебательного цикла, более эффективно деполяризуют постсинаптический нейрон . [27]
Временная структура серии импульсов или частоты выстрелов, вызванная стимулом, определяется как динамикой стимула, так и природой процесса нейронного кодирования. Стимулы, которые быстро меняются, имеют тенденцию вызывать точно рассчитанные по времени всплески. [28] (и быстро меняющаяся скорость стрельбы в PSTH) независимо от того, какая стратегия нейронного кодирования используется. Временное кодирование в узком смысле относится к временной точности реакции, которая возникает не только из динамики стимула, но, тем не менее, связана со свойствами стимула. Взаимодействие между стимулом и динамикой кодирования затрудняет идентификацию временного кода.
При временном кодировании обучение можно объяснить зависимыми от активности модификациями синаптической задержки. [29] Модификации сами по себе могут зависеть не только от частоты всплесков (кодирование скорости), но также от шаблонов времени всплесков (временное кодирование), т. е. могут быть частным случаем пластичности, зависящей от времени всплеска . [30]
Проблема временного кодирования отличается и независима от проблемы независимого кодирования. Если каждый спайк не зависит от всех остальных импульсов в цепочке, временной характер нейронного кода определяется поведением зависящей от времени скорости срабатывания r(t). Если r(t) медленно меняется со временем, код обычно называют кодом скорости, а если он меняется быстро, код называют временным.
Временное кодирование в сенсорных системах
[ редактировать ]Для очень коротких стимулов максимальная скорость возбуждения нейрона может оказаться недостаточно высокой, чтобы произвести более одного импульса. Из-за плотности информации о сокращенном стимуле, содержащейся в этом единственном спайке, может показаться, что время самого спайка должно передавать больше информации, чем просто средняя частота потенциалов действия за определенный период времени. Эта модель особенно важна для локализации звука , которая происходит в мозгу за миллисекунды. Мозг должен получить большое количество информации на основе относительно короткого нейронного ответа. Кроме того, если следует отличать низкую скорость срабатывания порядка десяти импульсов в секунду от произвольно близкой скорости кодирования различных стимулов, то нейрону, пытающемуся различить эти два стимула, возможно, придется подождать секунду или больше, чтобы накопить достаточно информации. Это не согласуется с многочисленными организмами, которые способны различать стимулы за миллисекунды, что позволяет предположить, что код скорости - не единственная действующая модель. [24]
Чтобы объяснить быстрое кодирование зрительных стимулов, было высказано предположение, что нейроны сетчатки кодируют визуальную информацию в течение латентного времени между началом стимула и первым потенциалом действия, также называемого задержкой до первого импульса или временем до первого импульса. [31] Этот тип временного кодирования выявлен также в слуховой и соматосенсорной системе. Основным недостатком такой схемы кодирования является ее чувствительность к собственным нейронным флуктуациям. [32] Было обнаружено, что в первичной зрительной коре макак время появления первого спайка относительно начала стимула дает больше информации, чем интервал между спайками. Однако интервал между импульсами можно использовать для кодирования дополнительной информации, что особенно важно, когда частота импульсов достигает своего предела, как в ситуациях с высоким контрастом. По этой причине временное кодирование может играть роль в кодировании определенных краев, а не постепенных переходов. [33]
млекопитающих Вкусовая система полезна для изучения временного кодирования из-за ее довольно четких стимулов и легко различимых реакций организма. [34] Временно закодированная информация может помочь организму различать разные вкусовые вещества одной и той же категории (сладкий, горький, кислый, соленый, умами), которые вызывают очень похожие реакции с точки зрения количества пиков. Временной компонент паттерна, вызываемый каждым веществом вкуса, можно использовать для определения его идентичности (например, разницы между двумя веществами горького вкуса, такими как хинин и денатониум). Таким образом, в вкусовой системе можно использовать как скоростное, так и временное кодирование: скоростное для основного типа вкуса, временное для более конкретной дифференциации. [35] Исследования вкусовой системы млекопитающих показали, что во временных паттернах популяций нейронов присутствует обилие информации, и эта информация отличается от той, которая определяется схемами кодирования скорости. Группы нейронов могут синхронизироваться в ответ на стимул. В исследованиях, посвященных передней кортикальной части мозга у приматов, в небольших популяциях нейронов были обнаружены точные закономерности с короткими временными масштабами (всего несколько миллисекунд), которые коррелировали с определенным поведением обработки информации. Однако по закономерностям можно было получить мало информации; Одна из возможных теорий заключается в том, что они представляют собой обработку высшего порядка, происходящую в мозгу. [25]
Как и в случае со зрительной системой, в митральных/тафтинговых клетках мышей обонятельной луковицы латентность первого спайка относительно начала обнюхивания, по-видимому, кодирует большую часть информации о запахе. Эта стратегия использования задержки пика позволяет быстро идентифицировать запах и отреагировать на него. Кроме того, некоторые митральные/тафтинговые клетки имеют специфические модели активации определенных запахов. Дополнительная информация такого типа могла бы помочь распознать определенный запах, но она не является абсолютно необходимой, поскольку среднее количество шипов за время обнюхивания животным также было хорошим идентификатором. [36] В том же духе эксперименты, проведенные с обонятельной системой кроликов, показали различные закономерности, которые коррелировали с разными подгруппами запахов, и аналогичный результат был получен в экспериментах с обонятельной системой саранчи. [24]
Приложения временного кодирования
[ редактировать ]Специфика временного кодирования требует высокоточной технологии для измерения информативных, надежных экспериментальных данных. Достижения в области оптогенетики позволяют неврологам контролировать спайки в отдельных нейронах, обеспечивая электрическое и пространственное разрешение отдельных клеток. Например, синий свет вызывает открытие светозависимого ионного канала родопсина , деполяризуя клетку и вызывая спайк. Когда клетка не воспринимает синий свет, канал закрывается, и нейрон перестает генерировать импульсы. Рисунок шипов соответствует рисунку стимулов синего света. Встраивая последовательности гена каналородопсина в ДНК мыши, исследователи могут контролировать спайки и, следовательно, определенное поведение мыши (например, заставлять мышь поворачивать налево). [37] Исследователи с помощью оптогенетики имеют инструменты для воздействия на разные временные коды в нейроне, сохраняя при этом одну и ту же среднюю скорость срабатывания, и тем самым могут проверить, происходит ли временное кодирование в определенных нейронных цепях. [38]
Оптогенетическая технология также потенциально может позволить исправить аномалии спайков, лежащие в основе некоторых неврологических и психологических расстройств. [38] Если нейроны действительно кодируют информацию в виде отдельных временных паттернов импульсов, ключевые сигналы можно пропустить, пытаясь взломать код, глядя только на среднюю скорость срабатывания. [24] Понимание любых временно закодированных аспектов нейронного кода и репликация этих последовательностей в нейронах может позволить лучше контролировать и лечить неврологические расстройства, такие как депрессия , шизофрения и болезнь Паркинсона . Регулирование интервалов спайков в отдельных клетках более точно контролирует активность мозга, чем введение фармакологических агентов внутривенно. [37]
Код фазы стрельбы
[ редактировать ]Код фазы срабатывания — это схема нейронного кодирования, которая объединяет код подсчета всплесков с привязкой ко времени, основанной на колебаниях . Этот тип кода учитывает метку времени для каждого всплеска в соответствии с привязкой ко времени, основанной на фазе локальных продолжающихся колебаний при низком уровне. [39] или высокие частоты. [40]
Было показано, что нейроны в некоторых сенсорных областях коры кодируют богатые натуралистические стимулы с точки зрения времени их спайков относительно фазы продолжающихся колебательных колебаний сети, а не только с точки зрения количества их спайков. [39] [41] Сигналы потенциала локального поля отражают колебания популяции (сети). Код фазы срабатывания часто классифицируется как временной код, хотя временная метка, используемая для всплесков (т. е. фазы колебаний сети), представляет собой ссылку с низким разрешением (крупнозернистую) для времени. В результате часто всего четырех дискретных значений фазы достаточно, чтобы представить всю информативность такого рода кода относительно фазы колебаний на низких частотах. Код фазы срабатывания в общих чертах основан на явлении прецессии фазы , наблюдаемом в клетках гиппокампа . Другая особенность этого кода заключается в том, что нейроны придерживаются предпочтительного порядка импульсов между группой сенсорных нейронов, что приводит к последовательности срабатываний. [42]
Было показано, что фазовый код в зрительной коре включает также высокочастотные колебания . [42] В рамках цикла гамма-колебаний каждый нейрон имеет свое предпочтительное относительное время срабатывания. В результате вся популяция нейронов генерирует последовательность импульсов длительностью примерно до 15 мс. [42]
Кодирование населения
[ редактировать ]Популяционное кодирование — это метод представления стимулов с использованием совместной деятельности ряда нейронов. При популяционном кодировании каждый нейрон имеет распределение ответов по некоторому набору входных данных, и ответы многих нейронов могут быть объединены для определения некоторой ценности входных данных. С теоретической точки зрения популяционное кодирование — одна из немногих математически хорошо сформулированных задач нейробиологии. Он охватывает основные особенности нейронного кодирования, но при этом достаточно прост для теоретического анализа. [43] Экспериментальные исследования показали, что эта парадигма кодирования широко используется в сенсорных и моторных областях мозга.
Например, в медиально-височной зоне (МТ) нейроны настроены на направление движения объекта. [44] В ответ на объект, движущийся в определенном направлении, многие нейроны в МТ срабатывают с шумовой и колоколообразной активностью по всей популяции. Направление движения объекта определяется на основе активности популяции, чтобы быть невосприимчивым к колебаниям, существующим в сигнале одного нейрона. Когда обезьян обучают перемещать джойстик по направлению к освещенной цели, один нейрон будет срабатывать в нескольких направлениях. Однако он срабатывает быстрее всего в одном направлении и медленнее в зависимости от того, насколько близко цель находится к «предпочтительному» направлению нейрона. [45] [46] Если каждый нейрон представляет движение в предпочтительном направлении и вычисляется векторная сумма всех нейронов (каждый нейрон имеет скорость срабатывания и предпочтительное направление), сумма указывает в направлении движения. Таким образом, популяция нейронов кодирует сигнал движения. [ нужна ссылка ] Этот конкретный код совокупности называется кодированием вектора совокупности .
Коды совокупности места и времени, называемые кодом усредненного локализованного синхронизированного ответа (ALSR), были получены для нейронного представления слуховых акустических стимулов. При этом используется как расположение или настройка внутри слухового нерва, так и фазовая синхронизация внутри каждого нервного волокна слухового нерва. Первое представление ALSR предназначалось для устойчивых гласных; [47] Представления ALSR высоты тона и формантных частот в сложных, нестационарных стимулах были позже продемонстрированы для звонкой высоты звука, [48] и формантные представления в согласных-гласных слогах. [49] Преимущество таких представлений заключается в том, что глобальные характеристики, такие как профили перехода высоты тона или формант, могут быть представлены как глобальные характеристики по всему нерву одновременно посредством кодирования скорости и места.
Популяционное кодирование имеет также ряд других преимуществ, включая снижение неопределенности из-за изменчивости нейронов и способность одновременно представлять ряд различных атрибутов стимула. Популяционное кодирование также намного быстрее, чем кодирование скорости, и может почти мгновенно отражать изменения в условиях стимула. [50] Отдельные нейроны в такой популяции обычно обладают разной, но перекрывающейся избирательностью, поэтому многие нейроны, но не обязательно все, реагируют на данный стимул.
Обычно функция кодирования имеет такое пиковое значение, что активность нейрона максимальна, если перцептивное значение близко к пиковому значению, и соответственно снижается для значений, менее близких к пиковому значению. [ нужна ссылка ] Отсюда следует, что фактическое воспринимаемое значение можно реконструировать по общей картине активности набора нейронов. Векторное кодирование является примером простого усреднения. Более сложным математическим методом проведения такой реконструкции является метод максимального правдоподобия , основанный на многомерном распределении ответов нейронов. Эти модели могут предполагать независимость, корреляции второго порядка, [51] или даже более подробные зависимости, такие как модели максимальной энтропии более высокого порядка , [52] или копулы . [53]
Корреляционное кодирование
[ редактировать ]Модель корреляционного кодирования возбуждения нейронов утверждает, что корреляции между потенциалами действия или «спайками» внутри серии спайков могут нести дополнительную информацию, выходящую за рамки простой синхронизации спайков. Ранние работы показали, что корреляция между сериями спайков может только уменьшить, но никогда не увеличить общую взаимную информацию, содержащуюся в двух сериях спайков, о признаке стимула. [54] Однако позже было показано, что это неверно. Корреляционная структура может повысить информативность, если корреляции шума и сигнала имеют противоположный знак. [55] Корреляции также могут нести информацию, отсутствующую в средней частоте срабатывания двух пар нейронов. Хороший пример этого существует в слуховой коре мартышки, анестезированной пентобарбиталом, в которой чистый тон вызывает увеличение количества коррелирующих спайков, но не увеличение средней частоты импульсов пар нейронов. [56]
Независимое кодирование шипов
[ редактировать ]Модель кодирования независимых спайков возбуждения нейронов утверждает, что каждый отдельный потенциал действия , или «спайк», независим от каждого другого спайка в последовательности спайков . [57] [58]
Кодирование позиции
[ редактировать ]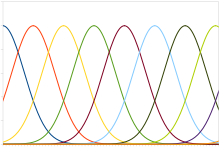
Типичный популяционный код включает в себя нейроны с кривой настройки Гаусса, средние значения которой изменяются линейно в зависимости от интенсивности стимула, а это означает, что нейрон наиболее сильно (с точки зрения импульсов в секунду) реагирует на стимул, близкий к среднему значению. Фактическую интенсивность можно определить как уровень стимула, соответствующий среднему значению нейрона с наибольшим ответом. Однако шум, присущий нейронным реакциям, означает, что функция оценки максимального правдоподобия является более точной.

Этот тип кода используется для кодирования непрерывных переменных, таких как положение суставов, положение глаз, цвет или частота звука. Любой отдельный нейрон слишком зашумлен, чтобы точно закодировать переменную с использованием скоростного кодирования, но целая популяция обеспечивает большую точность и достоверность. Для совокупности унимодальных кривых настройки, т. е. с одним пиком, точность обычно линейно масштабируется в зависимости от количества нейронов. Следовательно, для половины точности требуется вдвое меньше нейронов. Напротив, когда кривые настройки имеют несколько пиков, как в ячейках сетки , представляющих пространство, точность совокупности может экспоненциально масштабироваться в зависимости от количества нейронов. Это значительно уменьшает количество нейронов, необходимых для той же точности. [59]
Разреженное кодирование
[ редактировать ]Разреженный код — это когда каждый элемент кодируется сильной активацией относительно небольшого набора нейронов. Для каждого элемента, который нужно закодировать, это отдельное подмножество всех доступных нейронов. В отличие от кодирования с разрежением датчиков, кодирование с высокой плотностью датчиков подразумевает, что известна вся информация от возможных местоположений датчиков.
Как следствие, разреженность может быть сосредоточена на временной разреженности («активно относительно небольшое количество периодов времени») или на разреженности активированной популяции нейронов. В последнем случае это можно определить за один период времени как количество активированных нейронов по отношению к общему количеству нейронов в популяции. Похоже, это отличительная черта нейронных вычислений, поскольку по сравнению с традиционными компьютерами информация массово распределяется по нейронам. Разреженное кодирование естественных изображений создает вейвлет -ориентированные фильтры, которые напоминают рецептивные поля простых клеток зрительной коры. [60] Емкость разреженных кодов может быть увеличена за счет одновременного использования временного кодирования, как это происходит в обонятельной системе саранчи. [61]
Учитывая потенциально большой набор входных шаблонов, алгоритмы разреженного кодирования (например, разреженный автокодировщик ) пытаются автоматически найти небольшое количество репрезентативных шаблонов, которые при объединении в правильных пропорциях воспроизводят исходные входные шаблоны. Тогда разреженное кодирование входных данных состоит из этих репрезентативных шаблонов. Например, очень большой набор английских предложений может быть закодирован небольшим количеством символов (т.е. букв, цифр, знаков препинания и пробелов), объединенных в определенном порядке для конкретного предложения, и поэтому разреженное кодирование для английского языка будет таким: символы.
Линейная генеративная модель
[ редактировать ]Большинство моделей разреженного кодирования основаны на линейной генеративной модели. [62] В этой модели символы комбинируются линейным образом для аппроксимации входных данных.
Более формально, учитывая k-мерный набор входных векторов с действительными номерами , цель разреженного кодирования — определить n k-мерных базисных векторов , соответствующий рецептивным полям нейронов, вместе с разреженным n-мерным вектором весов или коэффициентов для каждого входного вектора, так что линейная комбинация базисных векторов с пропорциями, заданными коэффициентами, приводит к близкому приближению к входному вектору: . [63]




Кодировки, генерируемые алгоритмами, реализующими линейную генеративную модель, можно разделить на кодировки с мягкой разреженностью и кодировки с жесткой разреженностью . [62] Они относятся к распределению коэффициентов базисного вектора для типичных входных данных. Кодирование с мягкой разреженностью имеет гладкое распределение, подобное гауссовскому , но более пиковое, чем у гауссовского, со множеством нулевых значений, некоторыми небольшими абсолютными значениями, меньшим количеством больших абсолютных значений и очень немногими очень большими абсолютными значениями. Таким образом, многие базисные векторы активны. Жесткая разреженность, с другой стороны, указывает на то, что существует много нулевых значений, отсутствуют или практически отсутствуют малые абсолютные значения, меньше больших абсолютных значений и очень мало очень больших абсолютных значений, и, следовательно, лишь немногие из базисных векторов активны. Это привлекательно с метаболической точки зрения: меньше энергии используется, когда активируется меньше нейронов. [62]
Другой мерой кодирования является его критическая полнота или чрезмерная полнота . Если число базисных векторов n равно размерности k входного набора, кодирование называется критически полным. В этом случае плавные изменения входного вектора приводят к резким изменениям коэффициентов, и кодирование не может корректно обрабатывать небольшие масштабы, небольшие перемещения или шум во входных данных. Однако если количество базисных векторов превышает размерность входного набора, кодирование является чрезмерным . Сверхполное кодирование плавно интерполирует входные векторы и устойчиво к входному шуму. [64] По оценкам, первичная зрительная кора человека перегружена в 500 раз, так что, например, участок входных данных размером 14 x 14 (196-мерное пространство) кодируется примерно 100 000 нейронами. [62]
Другие модели основаны на поиске соответствия — алгоритме разреженной аппроксимации , который находит «наилучшие совпадающие» проекции многомерных данных, и обучении по словарю — методе обучения представлению, целью которого является поиск разреженного матричного представления входных данных в виде линейного сочетание базовых элементов, а также сами эти базовые элементы. [65] [66] [67]
Биологические доказательства
[ редактировать ]Разреженное кодирование может быть общей стратегией нейронных систем для увеличения объема памяти. Чтобы адаптироваться к окружающей среде, животные должны узнать, какие стимулы связаны с поощрениями или наказаниями, и отличать эти усиленные стимулы от похожих, но нерелевантных. Такие задачи требуют реализации ассоциативной памяти , специфичной для стимула , в которой только несколько нейронов из популяции реагируют на любой данный стимул, а каждый нейрон реагирует только на несколько стимулов из всех возможных стимулов.
Теоретические работы по разреженной распределенной памяти показали, что разреженное кодирование увеличивает емкость ассоциативной памяти за счет уменьшения перекрытия между представлениями. [68] Экспериментально скудное представление сенсорной информации наблюдалось во многих системах, включая зрение, [69] прослушивание, [70] трогать, [71] и обоняние. [72] Однако, несмотря на накопление доказательств широко распространенного разреженного кодирования и теоретических аргументов в пользу его важности, было трудно доказать, что разреженное кодирование улучшает стимульную специфичность ассоциативной памяти.
Считается, что в дрозофилы обонятельной системе редкое кодирование запаха клетками Кеньона тела гриба создает большое количество точно адресуемых мест для хранения воспоминаний, специфичных для запаха. [73] Разреженность контролируется цепью отрицательной обратной связи между клетками Кеньона и ГАМКергическими передними парными боковыми (APL) нейронами. Систематическая активация и блокада каждой ветви этой цепи обратной связи показывает, что клетки Кеньона активируют нейроны APL, а нейроны APL ингибируют клетки Кеньона. Нарушение петли обратной связи между клетками Кеньона и APL уменьшает разреженность реакций клеток Кеньона на запах, увеличивает корреляции между запахами и не позволяет мухам научиться различать похожие, но не разные запахи. Эти результаты позволяют предположить, что ингибирование обратной связи подавляет активность клеток Кеньона, поддерживая разреженное, декоррелированное кодирование запаха и, следовательно, запаховую специфичность воспоминаний. [74]
См. также
[ редактировать ]- Искусственная нейронная сеть
- Автоэнкодер
- Модель биологического нейрона
- Проблема привязки
- Когнитивная карта
- Глубокое обучение
- Теория интеграции функций
- Бабушка клетка
- Модели нейронных вычислений
- Нейронный коррелят
- Нейронное декодирование
- Нейронные колебания
- Рецептивное поле
- Разреженная распределенная память
- Векторное квантование
Ссылки
[ редактировать ]- ^ Браун Э.Н., Касс Р.Э., Митра П.П. (май 2004 г.). «Анализ данных множественных нейронных импульсов: современное состояние и проблемы будущего». Нат. Нейроски . 7 (5): 456–61. дои : 10.1038/nn1228 . ПМИД 15114358 . S2CID 562815 .
- ^ Джонсон, нокаут (июнь 2000 г.). «Нейронное кодирование» . Нейрон . 26 (3): 563–566. дои : 10.1016/S0896-6273(00)81193-9 . ISSN 0896-6273 . ПМИД 10896153 .
- ^ Jump up to: а б с Торп, С.Дж. (1990). «Время прибытия пиков: высокоэффективная схема кодирования для нейронных сетей» . В Экмиллере, Р.; Хартманн, Г.; Хауске, Г. (ред.). Параллельная обработка в нейронных системах и компьютерах (PDF) . Северная Голландия. стр. 91–94. ISBN 978-0-444-88390-2 .
- ^ Сенгупта Б., Лафлин С.Б., Нивен Дж.Э. (2014)Последствия преобразования в потенциалы действия при кодировании нейронной информации и энергоэффективности. PLOS Вычислительная биология 10 (1): e1003439. https://doi.org/10.1371/journal.pcbi.1003439
- ^ Герстнер, Вульфрам ; Кистлер, Вернер М. (2002). Модели импульсных нейронов: одиночные нейроны, популяции, пластичность . Издательство Кембриджского университета. ISBN 978-0-521-89079-3 .
- ^ Jump up to: а б с д Штейн Р.Б., Госсен Э.Р., Джонс К.Е. (май 2005 г.). «Нейрональная изменчивость: шум или часть сигнала?». Нат. Преподобный Нейроски . 6 (5): 389–97. дои : 10.1038/nrn1668 . ПМИД 15861181 . S2CID 205500218 .
- ^ Код памяти. http://www.scientificamerican.com/article/the-memory-code/
- ^ Чен, Г; Ван, LP; Цянь, JZ (2009). «Следы памяти на уровне нейронной популяции в гиппокампе мыши» . ПЛОС ОДИН . 4 (12): е8256. Бибкод : 2009PLoSO...4.8256C . дои : 10.1371/journal.pone.0008256 . ПМЦ 2788416 . ПМИД 20016843 .
- ^ Чжан, Х; Чен, Г; Куанг, Х; Цянь, JZ (ноябрь 2013 г.). «Картирование и расшифровка нейронных кодов NMDA-рецептор-зависимых энграмм памяти страха в гиппокампе» . ПЛОС ОДИН . 8 (11): e79454. Бибкод : 2013PLoSO...879454Z . дои : 10.1371/journal.pone.0079454 . ПМЦ 3841182 . ПМИД 24302990 .
- ^ Проект декодирования мозга. http://braindecodingproject.org/
- ^ Сотрудничество Саймонса в области глобального мозга. https://www.simonsfoundation.org/life-sciences/simons-collaboration-global-brain/
- ^ Буркас Г.Т. и Олбрайт Т.Д. Оценка сенсорных представлений в мозге. http://www.vcl.salk.edu/Publications/PDF/Buracas_Albright_1999_TINS.pdf
- ^ Герстнер В., Крайтер А.К., Маркрам Х., Герц А.В. (ноябрь 1997 г.). «Нейронные коды: скорострельность и не только» . Учеб. Натл. акад. наук. США . 94 (24): 12740–1. Бибкод : 1997PNAS...9412740G . дои : 10.1073/pnas.94.24.12740 . ПМК 34168 . ПМИД 9398065 .
- ^ Jump up to: а б с д и ж г час я дж Герстнер, Вульфрам. (2002). Модели спайковых нейронов: отдельные нейроны, популяции, пластичность . Кистлер, Вернер М., 1969-. Кембридж, Великобритания: Издательство Кембриджского университета. ISBN 0-511-07817-Х . OCLC 57417395 .
- ^ Кандель, Э.; Шварц, Дж.; Джессел, ТМ (1991). Принципы нейронауки (3-е изд.). Эльзевир. ISBN 978-0444015624 .
- ^ Адриан ЭД, Зоттерман Ю (1926). «Импульсы, производимые окончаниями сенсорных нервов: Часть II: Реакция одного конечного органа» . Дж Физиол . 61 (2): 151–171. дои : 10.1113/jphysicalol.1926.sp002281 . ПМЦ 1514782 . ПМИД 16993780 .
- ^ Форрест, доктор медицины (2014). «Внутриклеточная динамика кальция позволяет модели нейронов Пуркинье выполнять переключение и получать результаты вычислений на своих входных данных» . Границы вычислительной нейронауки . 8 : 86. дои : 10.3389/fncom.2014.00086 . ПМЦ 4138505 . ПМИД 25191262 .
- ^ Форрест, доктор медицины (декабрь 2014 г.). «Натриево-калиевый насос — это элемент обработки информации в вычислениях мозга» . Границы в физиологии . 5 (472): 472. doi : 10.3389/fphys.2014.00472 . ПМЦ 4274886 . ПМИД 25566080 .
- ^ Jump up to: а б с Голлиш, Т.; Мейстер, М. (22 февраля 2008 г.). «Быстрое нейронное кодирование в сетчатке с относительными пиковыми задержками» . Наука . 319 (5866): 1108–1111. Бибкод : 2008Sci...319.1108G . дои : 10.1126/science.1149639 . ISSN 0036-8075 . ПМИД 18292344 . S2CID 1032537 .
- ^ Даян, Питер; Эбботт, LF (2001). Теоретическая нейронаука: вычислительное и математическое моделирование нейронных систем . Издательство Массачусетского технологического института. ISBN 978-0-262-04199-7 .
- ^ Баттс Д.А., Венг С., Джин Дж. и др. (сентябрь 2007 г.). «Временная точность нейронного кода и временные рамки естественного зрения». Природа . 449 (7158): 92–5. Бибкод : 2007Natur.449...92B . дои : 10.1038/nature06105 . ПМИД 17805296 . S2CID 4402057 .
- ^ Сингх и Леви, «Пирамидальный нейрон консенсусного слоя V может поддерживать межимпульсно-интервальное кодирование» , PLoS ONE , 2017
- ^ Дж. Лео ван Хеммен, Т. Дж. Сейновски. 23 Проблемы системной нейронауки. Оксфордский университет. Пресс, 2006. с.143-158.
- ^ Jump up to: а б с д Теуниссен, Ф; Миллер, JP (1995). «Временное кодирование в нервных системах: строгое определение». Журнал вычислительной нейронауки . 2 (2): 149–162. дои : 10.1007/bf00961885 . ПМИД 8521284 . S2CID 206786736 .
- ^ Jump up to: а б Задор, Стивенс, Чарльз, Энтони. «Загадка мозга» . © Current Biology 1995, Том 5, № 12 . Проверено 4 августа 2012 г.
{{cite web}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Костал Л., Лански П., Роспарс Дж.П. (ноябрь 2007 г.). «Нейронное кодирование и всплеск случайности». Евро. Дж. Нейроски . 26 (10): 2693–701. дои : 10.1111/j.1460-9568.2007.05880.x . ПМИД 18001270 . S2CID 15367988 .
- ^ Гупта, Нитин; Сингх, Свикрити Саран; Стопфер, Марк (15 декабря 2016 г.). «Осцилляционные окна интеграции в нейронах» . Природные коммуникации . 7 : 13808. Бибкод : 2016NatCo...713808G . дои : 10.1038/ncomms13808 . ISSN 2041-1723 . ПМК 5171764 . ПМИД 27976720 .
- ^ Жоливе, Рено; Раух, Александр; Люшер, Ганс-Рудольф; Герстнер, Вульфрам (1 августа 2006 г.). «Прогнозирование времени спайков неокортикальных пирамидных нейронов с помощью простых пороговых моделей» . Журнал вычислительной нейронауки . 21 (1): 35–49. дои : 10.1007/s10827-006-7074-5 . ISSN 1573-6873 . ПМИД 16633938 . S2CID 8911457 .
- ^ Жоффруа, Э.; Эделин, Дж. М.; Виберт, Дж. Ф. (1994). «Обучение с помощью модификаций задержки» . В Экмане, Фрэнк Х. (ред.). Вычисления в нейронах и нейронных системах . Спрингер. стр. 133–8. ISBN 978-0-7923-9465-5 .
- ^ Сьёстрем, Йеспер и Вульфрам Герстнер. «Пластичность, зависящая от времени всплеска». Пластичность, зависящая от времени скачка 35 (2010).
- ^ Голлиш, Т.; Мейстер, М. (22 февраля 2008 г.). «Быстрое нейронное кодирование в сетчатке с относительными пиковыми задержками». Наука . 319 (5866): 1108–1111. Бибкод : 2008Sci...319.1108G . дои : 10.1126/science.1149639 . ПМИД 18292344 . S2CID 1032537 .
- ^ Уэйнриб, Жиль; Мишель, Тьёллен; Хашаяр, Пакдаман (7 апреля 2010 г.). «Внутренняя изменчивость задержки до первого всплеска». Биологическая кибернетика . 103 (1): 43–56. дои : 10.1007/s00422-010-0384-8 . ПМИД 20372920 . S2CID 7121609 .
- ^ Виктор, Джонатан Д. (2005). «Показатели шипованного поезда» . Современное мнение в нейробиологии . 15 (5): 585–592. дои : 10.1016/j.conb.2005.08.002 . ПМЦ 2713191 . ПМИД 16140522 .
- ^ Халлок, Роберт М.; Ди Лоренцо, Патрисия М. (2006). «Временное кодирование во вкусовой системе». Неврологические и биоповеденческие обзоры . 30 (8): 1145–1160. doi : 10.1016/j.neubiorev.2006.07.005 . ПМИД 16979239 . S2CID 14739301 .
- ^ Карлтон, Алан; Акколла, Риккардо; Саймон, Сидни А. (2010). «Кодирование вкусовой системы млекопитающих» . Тенденции в нейронауках . 33 (7): 326–334. дои : 10.1016/j.tins.2010.04.002 . ПМК 2902637 . ПМИД 20493563 .
- ^ Уилсон, Рэйчел I (2008). «Нейральные и поведенческие механизмы обонятельного восприятия» . Современное мнение в нейробиологии . 18 (4): 408–412. дои : 10.1016/j.conb.2008.08.015 . ПМК 2596880 . ПМИД 18809492 .
- ^ Jump up to: а б Карл Дисерот, Лекция. «Серия личностного роста: Карл Диссерот о взломе нейронного кода». Google Tech Talks. 21 ноября 2008 г. https://www.youtube.com/watch?v=5SLdSbp6VjM .
- ^ Jump up to: а б Хан Х, Цянь Х, Стерн П., Чуонг А.С., Бойден Э.С. «Информационные повреждения: оптические нарушения времени спайков и нейронной синхронности посредством слияния генов микробного опсина». Кембридж, Массачусетс: MIT Media Lad, 2009.
- ^ Jump up to: а б Монтемурро, Марсело А.; Раш, Мальте Дж.; Мураяма, Юсуке; Логотетис, Никос К.; Панцери, Стефано (2008). «Фазовое кодирование естественных зрительных стимулов в первичной зрительной коре» . Современная биология . 18 (5): 375–380. дои : 10.1016/j.cub.2008.02.023 . ПМИД 18328702 .
- ^ Фрис П., Николич Д., Сингер В. (июль 2007 г.). «Гамма-цикл». Тенденции нейробиологии . 30 (7): 309–16. дои : 10.1016/j.tins.2007.05.005 . ПМИД 17555828 . S2CID 3070167 .
- ^ Время прибытия всплесков: высокоэффективная схема кодирования для нейронных сетей. Архивировано 15 февраля 2012 г. в Wayback Machine , С. Дж. Торп - Параллельная обработка в нейронных системах, 1990 г.
- ^ Jump up to: а б с Хавенит М.Н., Ю.С., Бидерлак Дж., Чен Н.Х., Сингер В., Николич Д. (июнь 2011 г.). «Синхронность заставляет нейроны срабатывать последовательно, а свойства стимула определяют, кто впереди» . Дж. Нейроски . 31 (23): 8570–84. doi : 10.1523/JNEUROSCI.2817-10.2011 . ПМК 6623348 . ПМИД 21653861 .
- ^ Ву С., Амари С., Накахара Х. (май 2002 г.). «Кодирование и декодирование населения в нейронном поле: вычислительное исследование». Нейронный компьютер . 14 (5): 999–1026. дои : 10.1162/089976602753633367 . ПМИД 11972905 . S2CID 1122223 .
- ^ Маунселл Дж. Х., Ван Эссен, округ Колумбия (май 1983 г.). «Функциональные свойства нейронов средневисочной зрительной области обезьяны-макаки. I. Избирательность по направлению, скорости и ориентации стимула». Дж. Нейрофизиология . 49 (5): 1127–47. дои : 10.1152/jn.1983.49.5.1127 . ПМИД 6864242 . S2CID 8708245 .
- ^ «Введение в сенсорно-моторные системы, глава 38, страница 766» (PDF) . Архивировано из оригинала (PDF) 11 мая 2012 г. Проверено 3 февраля 2014 г.
- ^ Наука. 1986, 26 сентября; 233 (4771): 1416-9.
- ^ Сакс, Мюррей Б.; Янг, Эрик Д. (ноябрь 1979 г.). «Представление устойчивых гласных во временных аспектах паттернов разрядки популяций волокон слухового нерва». Журнал Акустического общества Америки . 66 (5): 1381–1403. Бибкод : 1979ASAJ...66.1381Y . дои : 10.1121/1.383532 . ПМИД 500976 .
- ^ Миллер, Мичиган; Сакс, МБ (июнь 1984 г.). «Представление высоты голоса в паттернах разрядки волокон слухового нерва». Исследование слуха . 14 (3): 257–279. дои : 10.1016/0378-5955(84)90054-6 . ПМИД 6480513 . S2CID 4704044 .
- ^ Миллер, Мичиган; Сакс, МБ (1983). «Представление стоповых согласных в паттернах разрядки волокон слухового нерва». Журнал Акустического общества Америки . 74 (2): 502–517. Бибкод : 1983ASAJ...74..502M . дои : 10.1121/1.389816 . ПМИД 6619427 .
- ^ Хьюбель Д.Х., Визель Т.Н. (октябрь 1959 г.). «Рецептивные поля отдельных нейронов полосатой коры головного мозга кошки» . Дж. Физиол . 148 (3): 574–91. doi : 10.1113/jphysicalol.1959.sp006308 . ПМЦ 1363130 . ПМИД 14403679 .
- ^ Шнайдман, Э; Берри, MJ; Сегев, Р; Бялек, В. (2006), «Слабые парные корреляции подразумевают сильно коррелированные состояния сети в нейронной популяции», Nature , 440 (7087): 1007–1012, arXiv : q-bio/0512013 , Bibcode : 2006Natur.440.1007S , doi : 10.1038/nature04701 , PMC 1785327 , PMID 16625187
- ^ Амари, С.Л. (2001), «Информационная геометрия в иерархии вероятностных распределений», IEEE Transactions on Information Theory , 47 (5): 1701–1711, CiteSeerX 10.1.1.46.5226 , doi : 10.1109/18.930911
- ^ Онкен, А; Грюневельдер, С; Мунк, MHJ; Обермайер, К. (2009), «Анализ кратковременных шумовых зависимостей количества спайков в префронтальной коре макак с использованием копул и преобразования фонарика», PLOS Comput Biol , 5 (11): e1000577, Bibcode : 2009PLSCB...5E0577O , doi : 10.1371/journal.pcbi.1000577 , PMC 2776173 , PMID 19956759
- ^ Джонсон, нокаут (июнь 1980 г.). «Сенсорная дискриминация: нервные процессы, предшествующие решению о различении». J Нейрофизиология . 43 (6): 1793–815. дои : 10.1152/Jn.1980.43.6.1793 . ПМИД 7411183 .
- ^ танковые; Шульц; Тревес; Роллс (1999). «Корреляции и кодирование информации в нервной системе» . Учебник по биологическим наукам . 266 (1423): 1001–12. дои : 10.1098/rspb.1999.0736 . ПМК 1689940 . ПМИД 10610508 .
- ^ Мерцених, ММ (июнь 1996 г.). «Первичное корковое представление звуков путем координации времени потенциала действия». Природа . 381 (6583): 610–3. Бибкод : 1996Natur.381..610D . дои : 10.1038/381610a0 . ПМИД 8637597 . S2CID 4258853 .
- ^ Даян П. и Эбботт LF. Теоретическая нейронаука: вычислительное и математическое моделирование нейронных систем . Кембридж, Массачусетс: MIT Press; 2001. ISBN 0-262-04199-5
- ^ Рике Ф., Варланд Д., де Рюйтер ван Стивенинк Р., Биалек В. Спайкс: изучение нейронного кода . Кембридж, Массачусетс: MIT Press; 1999. ISBN 0-262-68108-0
- ^ Матис А., Герц А.В., Стеммлер М.Б. (июль 2012 г.). «Разрешение вложенных нейронных представлений может быть экспоненциальным по количеству нейронов» . Физ. Преподобный Летт . 109 (1): 018103. Бибкод : 2012PhRvL.109a8103M . doi : 10.1103/PhysRevLett.109.018103 . ПМИД 23031134 .
- ^ Ольсхаузен, Бруно А; Филд, Дэвид Дж (1996). «Появление свойств рецептивного поля простых клеток путем изучения разреженного кода естественных изображений» (PDF) . Природа . 381 (6583): 607–609. Бибкод : 1996Natur.381..607O . дои : 10.1038/381607a0 . ПМИД 8637596 . S2CID 4358477 . Архивировано из оригинала (PDF) 23 ноября 2015 г. Проверено 29 марта 2016 г.
- ^ Гупта, Н; Стопфер, М. (6 октября 2014 г.). «Временный канал для информации в разреженном сенсорном кодировании» . Современная биология . 24 (19): 2247–56. дои : 10.1016/j.cub.2014.08.021 . ПМК 4189991 . ПМИД 25264257 .
- ^ Jump up to: а б с д Рен, Мартин; Зоммер, Фридрих Т. (2007). «Сеть, которая использует несколько активных нейронов для кодирования зрительной информации, предсказывает разнообразные формы корковых рецептивных полей» (PDF) . Журнал вычислительной нейронауки . 22 (2): 135–146. дои : 10.1007/s10827-006-0003-9 . ПМИД 17053994 . S2CID 294586 .
- ^ Ли, Хонглак; Битва, Алексис; Райна, Раджат; Нг, Эндрю Ю. (2006). «Эффективные алгоритмы разреженного кодирования» (PDF) . Достижения в области нейронных систем обработки информации .
- ^ Ольсхаузен, Бруно А.; Филд, Дэвид Дж. (1997). «Разреженное кодирование с чрезмерно полным базисным набором: стратегия, используемая V1?» . Исследование зрения . 37 (23): 3311–3325. дои : 10.1016/s0042-6989(97)00169-7 . ПМИД 9425546 .
- ^ Чжан, Чжифэн; Маллат, Стефан Г.; Дэвис, Джеффри М. (июль 1994 г.). «Адаптивные частотно-временные разложения». Оптическая инженерия . 33 (7): 2183–2192. Бибкод : 1994OptEn..33.2183D . дои : 10.1117/12.173207 . ISSN 1560-2303 .
- ^ Пати, ЮК; Резайфар, Р.; Кришнапрасад, П.С. (ноябрь 1993 г.). «Погоня за ортогональным сопоставлением: аппроксимация рекурсивной функции с применением к вейвлет-разложению». Материалы 27-й Асиломарской конференции по сигналам, системам и компьютерам . С. 40–44 т.1. CiteSeerX 10.1.1.348.5735 . дои : 10.1109/ACSSC.1993.342465 . ISBN 978-0-8186-4120-6 . S2CID 16513805 .
- ^ Ниделл, Д.; Тропп, Дж. А. (1 мая 2009 г.). «CoSaMP: итеративное восстановление сигнала из неполных и неточных выборок». Прикладной и вычислительный гармонический анализ . 26 (3): 301–321. arXiv : 0803.2392 . дои : 10.1016/j.acha.2008.07.002 . ISSN 1063-5203 . S2CID 1642637 .
- ^ Хизер, Пентти. Разреженная распределенная память. Пресс МТИ, 1988 г.
- ^ Винье, МЫ; Галлант, Дж.Л. (2000). «Редкое кодирование и декорреляция в первичной зрительной коре при естественном зрении». Наука . 287 (5456): 1273–1276. Бибкод : 2000Sci...287.1273V . CiteSeerX 10.1.1.456.2467 . дои : 10.1126/science.287.5456.1273 . ПМИД 10678835 .
- ^ Громадка, Т; Дьюиз, MR; Задор, AM (2008). «Скудное представление звуков в неанестезированной слуховой коре» . ПЛОС Биол . 6 (1): е16. дои : 10.1371/journal.pbio.0060016 . ПМК 2214813 . ПМИД 18232737 .
- ^ Крючком, С; Пуле, JFA; Кремер, Ю; Петерсен, CCH (2011). «Синаптические механизмы, лежащие в основе редкого кодирования активного прикосновения» . Нейрон . 69 (6): 1160–1175. дои : 10.1016/j.neuron.2011.02.022 . ПМИД 21435560 .
- ^ Ито, я; Онг, RCY; Раман, Б; Стопфер, М. (2008). «Редкое представление запахов и обонятельное обучение» . Нат Нейроски . 11 (10): 1177–1184. дои : 10.1038/nn.2192 . ПМК 3124899 . ПМИД 18794840 .
- ^ Разреженная память — это точная память. Блог Oxford Science. 28 февраля 2014 г. http://www.ox.ac.uk/news/science-blog/sparse-memory-precision-memory.
- ^ Лин, Эндрю С. и др. « Редкая, декоррелированная кодировка запахов в теле гриба улучшает усвоенное распознавание запахов ». Природная неврология 17.4 (2014): 559-568.
Дальнейшее чтение
[ редактировать ]- Фёлдиак П., Эндрес Д., Разреженное кодирование , Scholarpedia , 3(1):2984, 2008.
- Даян П. и Эбботт Л.Ф. Теоретическая нейронаука: вычислительное и математическое моделирование нейронных систем . Кембридж, Массачусетс: MIT Press; 2001. ISBN 0-262-04199-5
- Рике Ф., Варланд Д., де Рюйтер ван Стивенинк Р., Биалек В. Спайкс: исследование нейронного кода . Кембридж, Массачусетс: MIT Press; 1999. ISBN 0-262-68108-0
- Ольсхаузен, бакалавр; Филд, диджей (1996). «Появление свойств рецептивного поля простых клеток путем изучения разреженного кода естественных изображений». Природа . 381 (6583): 607–9. Бибкод : 1996Natur.381..607O . дои : 10.1038/381607a0 . ПМИД 8637596 . S2CID 4358477 .
- Цянь, JZ.; и др. (2014). «О первоначальном картировании мозговой активности кода эпизодической и семантической памяти в гиппокампе» . Нейробиология обучения и памяти . 105 : 200–210. дои : 10.1016/j.nlm.2013.06.019 . ПМЦ 3769419 . ПМИД 23838072 .