Нейронные корреляты сознания
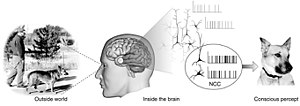
Нейронные корреляты сознания ( НКС ) — это минимальный набор нейрональных событий и механизмов, достаточный для возникновения психических состояний , с которыми они связаны. [2] Нейробиологи используют эмпирические подходы для обнаружения нейронных коррелятов субъективных явлений; то есть нейронные изменения, которые обязательно и регулярно коррелируют с конкретным опытом. [3] [4] Набор должен быть минимальным , поскольку, согласно материалистическому предположению, что мозг достаточен для того, чтобы вызвать любой данный сознательный опыт, вопрос заключается в том, какие из его компонентов необходимы для его создания.
Нейробиологический подход к сознанию
[ редактировать ]Наука и о сознании должна объяснить точную взаимосвязь между субъективными психическими состояниями и состояниями мозга, природу взаимосвязи между сознательным разумом электрохимическими проблема взаимодействиями в организме ( разума и тела ). Прогресс в нейропсихологии и нейрофилософии произошел благодаря сосредоточению внимания на теле, а не на разуме. В этом контексте нейрональные корреляты сознания можно рассматривать как его причины, а сознание можно рассматривать как зависящее от состояния свойство неопределенной сложной , адаптивной и тесно взаимосвязанной биологической системы. [5]
Обнаружение и характеристика нейронных коррелятов не предлагает причинной теории сознания, которая могла бы объяснить, как определенные системы что-либо воспринимают, так называемую сложную проблему сознания . [6] но понимание NCC может стать шагом к причинной теории. Большинство нейробиологов предполагают, что переменные, вызывающие сознание, следует искать на уровне нейронов, что регулируется классической физикой. Существуют теории квантового сознания , основанные на квантовой механике . [7]
В нейронных сетях существует очевидная избыточность и параллелизм, поэтому, хотя активность в одной группе нейронов может коррелировать с восприятием в одном случае, другая популяция может опосредовать связанное восприятие, если первая популяция потеряна или инактивирована. Возможно, каждое феноменальное, субъективное состояние имеет нейронный коррелят. Если NCC можно вызвать искусственно, субъект будет испытывать соответствующее восприятие, в то время как возмущение или инактивация области корреляции для конкретного восприятия повлияет на восприятие или заставит его исчезнуть, создавая причинно-следственную связь от нейронной области к природа восприятия.
Предложения, выдвигавшиеся на протяжении многих лет, включают: что характеризует NCC? Что общего между NCC для видения и слышания? Будет ли NCC задействовать все пирамидные нейроны коры в любой момент времени? Или это всего лишь подмножество проекционных клеток дальнего действия в лобных долях, которые проецируются на сенсорную кору спины? Нейроны, которые срабатывают ритмично? Нейроны, которые срабатывают синхронно ? [8]
Растущая способность нейробиологов манипулировать нейронами, используя методы молекулярной биологии в сочетании с оптическими инструментами (например, Адамантидис и др., 2007 ), зависит от одновременной разработки соответствующих поведенческих тестов и модельных организмов, поддающихся крупномасштабному геномному анализу и манипуляциям. Именно сочетание мелкозернистого нейронного анализа у животных со все более чувствительными методами психофизической визуализации и визуализации мозга у людей, дополненное разработкой надежной теоретической прогностической системы, приведет, как мы надеемся, к рациональному пониманию сознания, одного из центральных процессов. тайны жизни.
Уровень возбуждения и содержание сознания
[ редактировать ]Есть два общих, но различных измерения термина « сознание» . [9] один связан с возбуждением и состояниями сознания , а другой — с содержанием сознания и состояниями сознания . Чтобы осознавать что -либо, мозг должен находиться в относительно высоком состоянии возбуждения (иногда называемом бдительностью ), будь то бодрствование или быстрый сон , ярко переживаемый во сне, хотя обычно не запоминающийся. Уровень возбуждения мозга колеблется в соответствии с циркадным ритмом, но на него могут влиять недостаток сна, наркотики и алкоголь, физические нагрузки и т. д. Возбуждение можно измерить поведенчески по амплитуде сигнала, который запускает некоторую критериальную реакцию (например, уровень звука, необходимый для того, чтобы вызвать движение глаз или поворот головы в сторону источника звука). Клиницисты используют системы оценки, такие как шкала комы Глазго, для оценки уровня возбуждения у пациентов.
Состояния высокого возбуждения связаны с состояниями сознания, имеющими определенное содержание: видение, слушание, воспоминание, планирование или фантазирование о чем-то. Разные уровни или состояния сознания связаны с разными видами сознательного опыта. Состояние «бодрствования» сильно отличается от состояния «сновидения» (например, в последнем мало или совсем нет саморефлексии) и от состояния глубокого сна. Во всех трех случаях затрагивается основная физиология мозга, как и в измененных состояниях сознания , например, после приема наркотиков или во время медитации, когда сознательное восприятие и понимание могут быть усилены по сравнению с обычным состоянием бодрствования.
Клиницисты говорят о нарушениях сознания, таких как « коматозное состояние », « стойкое вегетативное состояние » (PVS) и « состояние минимального сознания » (MCS). Здесь «состояние» относится к разным «количествам» внешнего/физического сознания: от полного отсутствия в коме, стойкого вегетативного состояния и общей анестезии до колеблющейся и ограниченной формы сознательного ощущения в минимально сознательном состоянии, таком как ходьба во сне или во время сложного парциального эпилептического припадка. [10] Репертуар сознательных состояний или переживаний, доступных пациенту в минимально сознательном состоянии, сравнительно ограничен. При смерти мозга возбуждения нет, но неизвестно, прервалась ли субъективность переживания, а не его наблюдаемая связь с организмом. Функциональная нейровизуализация показала, что части коры все еще активны у вегетативных пациентов, которые предположительно находятся без сознания; [11] однако эти области, по-видимому, функционально оторваны от ассоциативных областей коры, активность которых необходима для осознания.
Потенциальное богатство сознательного опыта, по-видимому, увеличивается от глубокого сна к сонливости и к полному бодрствованию, что можно оценить количественно, используя понятия теории сложности, которые включают в себя как размерность, так и степень детализации сознательного опыта, чтобы дать интегрированную теоретическую информацию . сознание. [12] По мере увеличения поведенческого возбуждения увеличивается диапазон и сложность возможного поведения. Однако для быстрого сна характерна атония , низкое двигательное возбуждение и человека трудно разбудить, но сохраняется высокая метаболическая и электрическая активность мозга и живое восприятие.
Многие ядра с отчетливыми химическими характеристиками в таламусе , среднем мозге и мосту должны функционировать, чтобы субъект находился в состоянии возбуждения мозга, достаточном для того, чтобы вообще что-либо испытывать. Таким образом, эти ядра относятся к факторам, способствующим сознанию. И наоборот, вполне вероятно, что специфическое содержание любого конкретного сознательного ощущения опосредовано определенными нейронами коры и связанными с ними сателлитными структурами, включая миндалевидное тело , таламус , Клауструм и базальные ганглии .
Нейрональная основа восприятия
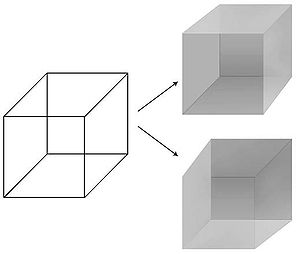
[ редактировать ]Возможность точного манипулирования визуальными восприятиями во времени и пространстве сделала зрение предпочтительным способом поиска NCC. Психологи усовершенствовали ряд техник – маскирование , бинокулярное соперничество , постоянное подавление вспышки , слепота, вызванная движением , слепота к изменениям , слепота по невнимательности – в которых кажущаяся простая и недвусмысленная связь между физическим стимулом в мире и связанным с ним восприятием в уединении человека разум субъекта нарушен. [13] В частности, стимул может быть перцептивно подавлен на секунды или даже минуты: изображение проецируется в один из глаз наблюдателя, но остается невидимым, не видимым. Таким образом, можно изолировать нервные механизмы, которые реагируют на субъективное восприятие, а не на физический стимул, что позволяет отслеживать зрительное сознание в мозгу. В перцептивной иллюзии физический стимул остается фиксированным, в то время как восприятие колеблется. Самый известный пример — куб Неккера , 12 линий которого можно воспринимать по глубине одним из двух способов.
Иллюзия восприятия, которой можно точно управлять, — это бинокулярное соперничество . Здесь небольшое изображение, например, горизонтальная решетка, представляется левому глазу, а другое изображение, например, вертикальная решетка, отображается в соответствующем месте в правом глазу. Несмотря на постоянный зрительный стимул, наблюдатели сознательно видят, как горизонтальная решетка каждые несколько секунд чередуется с вертикальной. Мозг не позволяет одновременно воспринимать оба изображения.
Логотетис и коллеги. [15] зафиксировали различные зрительные области коры у бодрствующих макак, выполняющих задачу бинокулярного соперничества. Обезьян-макак можно научить сообщать, видят ли они левое или правое изображение. Распределение времени переключения и то, как изменение контраста в одном глазу влияет на это, не оставляет сомнений в том, что обезьяны и люди испытывают одно и то же основное явление. В первичной зрительной коре (V1) только небольшая часть клеток слабо модулировала свой ответ в зависимости от восприятия обезьяны, в то время как большинство клеток реагировало на тот или иной стимул сетчатки, мало обращая внимания на то, что животное воспринимало в этот момент. Но в высокоуровневой области коры, такой как нижняя височная кора вдоль вентрального потока, почти все нейроны реагировали только на доминирующий в восприятии стимул, так что клетка «лица» активировалась только тогда, когда животное давало понять, что видит лицо, а не лицо. образец, представленный другому глазу. Это означает, что в NCC задействованы нейроны, активные в нижневисочной коре: вполне вероятно, что необходимы специфические реципрокные действия нейронов нижневисочной и частей префронтальной коры.
Ряд экспериментов с фМРТ , в которых использовалось бинокулярное соперничество и связанные с ним иллюзии для выявления гемодинамической активности, лежащей в основе зрительного сознания у людей, довольно убедительно демонстрируют, что активность на верхних стадиях вентрального пути (например, веретенообразной области лица и области парагиппокампального места ) а также в ранних областях, включая V1 и латеральное коленчатое ядро (LGN), следуют за восприятием, а не за стимулом сетчатки. [16] Кроме того, ряд фМРТ [17] [18] и эксперименты DTI [19] предполагают, что V1 необходим, но недостаточен для зрительного сознания. [20]
В родственном феномене восприятия, подавлении вспышки , восприятие, связанное с изображением, проецируемым в один глаз, подавляется путем проецирования другого изображения в другой глаз, в то время как исходное изображение остается. Его методологическое преимущество перед бинокулярным соперничеством состоит в том, что время перцептивного перехода определяется внешним триггером, а не внутренним событием. Большинство клеток нижней височной коры и верхней височной борозды обезьян, обученных сообщать о своем восприятии во время подавления вспышки, следуют восприятию животного: когда воспринимается предпочтительный стимул клетки, клетка реагирует. Если изображение все еще присутствует на сетчатке, но перцептивно подавлено, клетка замолкает, хотя первичные нейроны зрительной коры срабатывают. [21] [22] Записи одиночных нейронов в медиальной височной доле пациентов с эпилепсией во время подавления вспышки также демонстрируют отмену ответа, когда предпочтительный стимул присутствует, но перцептивно замаскирован. [23]
Глобальные расстройства сознания
[ редактировать ]Учитывая отсутствие какого-либо общепринятого критерия минимальных нейрональных коррелятов, необходимых для сознания, различие между пациентом с устойчивой вегетативной фазой, у которого наблюдаются регулярные переходы волн сна и который может двигаться или улыбаться, и пациентом с минимально сознательным состоянием, который может общаться (иногда ) осмысленным образом (например, дифференциальными движениями глаз) и проявляющим некоторые признаки сознания, часто бывает трудным. При глобальной анестезии пациент не должен испытывать психологической травмы, но уровень возбуждения должен быть совместим с клиническими потребностями.

зависящая от уровня кислорода в крови, ФМРТ, продемонстрировала нормальные закономерности мозговой активности у пациента в вегетативном состоянии после тяжелой черепно-мозговой травмы, когда его попросили представить игру в теннис или посещение комнат в его / ее доме. [25] Дифференциальная визуализация мозга пациентов с такими глобальными нарушениями сознания (включая акинетический мутизм ) показывает, что дисфункция широко распространенной кортикальной сети, включая медиальные и латеральные префронтальные и теменные ассоциативные области, связана с глобальной потерей сознания. [26] Нарушение сознания при эпилептических припадках височной доли также сопровождалось снижением мозгового кровотока в лобной и теменной ассоциативной коре и увеличением срединных структур, таких как медиодорсальный таламус . [27]
Относительно локальные двусторонние повреждения срединных (парамедианных) подкорковых структур также могут вызывать полную потерю сознания. [28] Таким образом, эти структуры обеспечивают и контролируют возбуждение мозга (определяемое метаболической или электрической активностью) и являются необходимыми нейронными коррелятами. Одним из таких примеров является гетерогенное скопление из более чем двух десятков ядер на каждой стороне верхнего отдела ствола мозга (мост, средний мозг и задний гипоталамус), которые в совокупности называются ретикулярной активирующей системой (РАС). Их аксоны широко распространены по всему мозгу. Эти ядра – трехмерные совокупности нейронов со своей цитоархитектурой и нейрохимической идентичностью – выделяют отдельные нейромодуляторы, такие как ацетилхолин, норадреналин/норадреналин, серотонин, гистамин и орексин/гипокретин, чтобы контролировать возбудимость таламуса и переднего мозга, опосредуя чередование между бодрствование и сон, а также общий уровень поведенческого и мозгового возбуждения. Однако после такой травмы со временем возбудимость таламуса и переднего мозга может восстановиться и сознание может вернуться. [29] Еще одним фактором, обеспечивающим сознание, являются пять или более внутриламинарных ядер (ILN) таламуса. Они получают сигналы от многих ядер ствола мозга и сильно проецируются непосредственно в базальные ганглии и, более распределенным образом, в слой I большей части неокортекса. Сравнительно небольшой (1 см. 3 или меньше) двусторонние поражения таламического ILN полностью отключают всякое сознание. [30]
Прогнозы вперед и обратной связи
[ редактировать ]Многие действия в ответ на сенсорные сигналы являются быстрыми, преходящими, стереотипными и бессознательными. [31] Их можно рассматривать как корковые рефлексы, и они характеризуются быстрыми и несколько стереотипными реакциями, которые могут принимать форму довольно сложного автоматизированного поведения, как это наблюдается, например, при сложных парциальных эпилептических припадках. Эти автоматические реакции, иногда называемые поведением зомби , [32] Его можно противопоставить более медленному, универсальному сознательному режиму, который медленнее обрабатывает более широкие, менее стереотипные аспекты сенсорной информации (или ее отражению, как в образах) и требует времени для принятия решения о соответствующих мыслях и реакциях. Без такого режима сознания потребовалось бы огромное количество различных режимов зомби, чтобы реагировать на необычные события.
Особенностью, отличающей человека от большинства животных, является то, что мы не рождаемся с обширным репертуаром поведенческих программ, которые позволили бы нам выжить самостоятельно (« физиологическая недоношенность »). Чтобы компенсировать это, у нас есть непревзойденная способность учиться, то есть сознательно приобретать такие программы путем подражания или исследования. Когда эти программы осознанно усвоены и достаточно реализованы, они могут стать автоматизированными до такой степени, что их выполнение происходит за пределами нашего сознания. Возьмем, к примеру, невероятную мелкую моторику, развиваемую при игре сонаты Бетховена для фортепиано, или сенсомоторную координацию, необходимую для езды на мотоцикле по извилистой горной дороге. Такое сложное поведение возможно только потому, что достаточное количество задействованных подпрограмм может выполняться с минимальным или даже приостановленным сознательным контролем. Фактически, сознательная система может в некоторой степени вмешиваться в работу этих автоматизированных программ. [33]
С эволюционной точки зрения очевидно имеет смысл иметь как автоматизированные поведенческие программы, которые можно быстро выполнять стереотипным и автоматизированным образом, так и немного более медленную систему, дающую время для обдумывания и планирования более сложного поведения. Этот последний аспект может быть одной из основных функций сознания. Другие философы, однако, предположили, что сознание не является необходимым для какого-либо функционального преимущества в эволюционных процессах. [34] [35] Они утверждают, что никто не дал причинно-следственного объяснения того, почему функционально эквивалентный бессознательный организм (то есть философский зомби ) не может достичь тех же преимуществ выживания, что и сознательный организм. Если эволюционные процессы не видят разницы между функцией F, выполняемой сознательным организмом O и бессознательным организмом O* , то неясно, какое адаптивное преимущество может обеспечить сознание. [36] В результате эксаптивное объяснение сознания завоевало поддержку некоторых теоретиков, которые утверждают, что сознание развилось не как адаптация, а как экзаптация, возникшая в результате других событий, таких как увеличение размера мозга или перестройка коры. [37] Сознание в этом смысле сравнивают со слепым пятном на сетчатке, где оно не является адаптацией сетчатки, а всего лишь побочным продуктом того, как устроены аксоны сетчатки. [38] Некоторые ученые, в том числе Пинкер , Хомский , Эдельман и Лурия, указали на важность появления человеческого языка как важного регулирующего механизма обучения и памяти в контексте развития сознания высшего порядка.
Кажется возможным, что зрительные режимы зомби в коре преимущественно используют дорсальный поток в теменной области. [31] Однако теменная активность может влиять на сознание, оказывая внимание на вентральный поток, по крайней мере, при некоторых обстоятельствах. Сознательный режим зрения во многом зависит от ранних зрительных областей (за пределами V1) и особенно от вентрального потока.
На первый взгляд сложная визуальная обработка (например, обнаружение животных в естественных, загроможденных сценах) может быть выполнена корой головного мозга человека за 130–150 мс. [39] [40] слишком короткий для того, чтобы произошли движения глаз и сознательное восприятие. Более того, такие рефлексы, как окуловестибулярный рефлекс, происходят еще быстрее. Вполне вероятно, что такое поведение опосредуется чисто прямой движущейся волной пиковой активности, которая проходит от сетчатки через V1 в V4, IT и префронтальную кору, пока не воздействует на мотонейроны в спинном мозге, которые контролируют нажатие пальца ( как в типичном лабораторном эксперименте). Гипотеза о том, что основная обработка информации осуществляется с прямой связью, наиболее непосредственно подтверждается коротким временем (около 100 мс), необходимым для появления избирательного ответа в ИТ-клетках.
И наоборот, считается, что сознательное восприятие требует более устойчивой, реверберационной нейронной активности, скорее всего, через глобальную обратную связь от лобных областей неокортекса обратно к сенсорным областям коры. [20] оно накапливается с течением времени, пока не превысит критический порог. В этот момент устойчивая нервная активность быстро распространяется на теменные, префронтальные и передние поясные области коры, таламус, клауструм и связанные с ними структуры, которые поддерживают кратковременную память, мультимодальную интеграцию, планирование, речь и другие процессы, тесно связанные с сознанием. Конкуренция препятствует одновременному и активному представлению более чем одного или очень небольшого числа восприятий. Это основная гипотеза глобального рабочего пространства . теории сознания [41] [42]
Короче говоря, хотя быстрая, но преходящая нейронная активность в таламо-кортикальной системе может опосредовать сложное поведение без осознанных ощущений, предполагается, что сознание требует устойчивой, но хорошо организованной нейронной активности, зависящей от дальней кортико-кортикальной обратной связи.
История
[ редактировать ]Нейробиолог Кристфрид Якоб (1866–1956) утверждал, что единственные состояния, которые должны иметь нейронные корреляты, — это прямые ощущения и реакции; это называются «интонации». [ нужна ссылка ]
Нейрофизиологические исследования на животных дали некоторое представление о нейронных коррелятах сознательного поведения. Вернон Маунткасл в начале 1960-х годов приступил к изучению этого набора проблем, который он назвал «проблемой разума/мозга», путем изучения нейронной основы восприятия в соматической сенсорной системе . Его лаборатории в Университете Джонса Хопкинса, наряду с Эдвардом В. Эвартсом из НИЗ, были одними из первых, кто зафиксировал нервную активность поведение обезьян. Пораженная элегантностью подхода С. С. Стивенса к оценке величины, группа Маунткасла обнаружила три различных модальности соматических ощущений, имеющих один общий когнитивный признак: во всех случаях частота импульсов периферических нейронов была линейно связана с силой вызванного восприятия. Совсем недавно Кен Х. Бриттен, Уильям Т. Ньюсом и К. Дэниел Зальцман показали, что в области MT у обезьян нейроны реагируют с изменчивостью, что позволяет предположить, что они являются основой принятия решений о направлении движения. Сначала они показали, что частота нейронов предсказывает решения, используя теорию обнаружения сигналов, а затем, что стимуляция этих нейронов может предсказуемо исказить решение. За такими исследованиями соматической сенсорной системы последовал Ранульфо Ромо, чтобы подтвердить, используя другое восприятие и область мозга, что небольшое количество нейронов в одной области мозга лежит в основе перцептивных решений.
Другие лабораторные группы последовали плодотворной работе Маунткасла, связавшей когнитивные переменные с активностью нейронов и решая более сложные когнитивные задачи. Хотя обезьяны не могут говорить о своем восприятии, были созданы поведенческие задачи, в которых животные сообщали невербально, например, производя движения рук. Многие из этих исследований используют перцептивные иллюзии как способ отделить ощущения ( т. е . сенсорную информацию, которую получает мозг) от восприятий ( т. е. того, как сознание их интерпретирует). Нейронные паттерны, которые представляют восприятие, а не просто сенсорную информацию, интерпретируются как отражающие нейронный коррелят сознания.
Используя такой дизайн, Никос Логотетис и его коллеги обнаружили нейроны, отражающие восприятие, в височной доле. Они создали экспериментальную ситуацию, в которой противоречивые изображения предъявлялись разным глазам ( т. е . бинокулярное соперничество ). В таких условиях люди сообщают о бистабильном восприятии: они попеременно воспринимают то одно, то другое изображение. Логотетис и его коллеги научили обезьян движениями рук сообщать, какой образ они воспринимают. Нейроны височной доли в экспериментах с Логотетисом часто отражали то, что воспринимали обезьяны. Нейроны с такими свойствами реже наблюдались в первичной зрительной коре, что соответствует относительно ранним стадиям зрительной обработки. Другая серия экспериментов с использованием бинокулярного соперничества на людях показала, что определенные слои коры могут быть исключены как кандидаты на нейронный коррелят сознания. Логотетис и его коллеги переключали изображения между глазами во время восприятия одного из изображений. Удивительно, но восприятие оставалось стабильным. Это означает, что сознательное восприятие оставалось стабильным, и в то же время основной входной сигнал в слой 4, который является входным слоем, в зрительной коре изменился. Следовательно, слой 4 не может быть частью нейронного коррелята сознания. Михаил Лебедев и его коллеги наблюдали аналогичный феномен в префронтальной коре обезьян. В своих экспериментах обезьяны сообщали о воспринимаемом направлении движения зрительного стимула (которое могло быть иллюзией), совершая движения глаз. Некоторые нейроны префронтальной коры представляли собой действительные, а некоторые — воспринимаемые смещения стимула. Наблюдение за нейронами, связанными с восприятием, в префронтальной коре согласуется с теорией Кристофа Коха и Фрэнсиса Крика , которые постулировали, что нейронный коррелят сознания находится в префронтальной коре. Сторонники распределенной нейрональной обработки, вероятно, могут оспорить точку зрения о том, что сознание имеет точную локализацию в мозге.
Фрэнсис Крик написал популярную книгу « Удивительная гипотеза », в которой утверждается, что нейронный коррелят сознания находится в наших нервных клетках и связанных с ними молекулах. Крик и его коллега Кристоф Кох [43] стремились избежать философских дебатов, связанных с изучением сознания, подчеркивая поиск «корреляции», а не «причинности». [ нужно обновить ]
Существует много разногласий относительно природы этого коррелята ( например , требует ли он синхронных импульсов нейронов в разных областях мозга? Необходима ли совместная активация лобных или теменных областей?). Философ Дэвид Чалмерс утверждает, что нейронный коррелят сознания, в отличие от других коррелятов, таких как память, не сможет дать удовлетворительного объяснения этого явления; он называет это трудной проблемой сознания . [44] [45]
См. также
[ редактировать ]- Сознание животных
- Искусственное сознание
- Проблема привязки
- Локус моста
- Когнитивная карта
- Концептуальное пространство
- Теория глобального рабочего пространства
- Сложная проблема сознания
- Теории сознания высшего порядка
- Схема изображения
- Теоретико-информационная смерть
- Интегрированная теория информации
- ЛИДА (когнитивная архитектура)
- Модели нейронных вычислений
- Модель с несколькими черновиками
- Трилемма Мюнхгаузена
- Нейронное кодирование
- Нейронное декодирование
- Нейронный субстрат
- Философия разума
- Квантовое познание
- Квантовый разум
Примечания
[ редактировать ]- ^ Кох 2004 , Рисунок 1.1 Нейрональные корреляты сознания с. 16.
- ^ Кох 2004 , с. 304.
- ^ см . здесь . Архивировано 13 марта 2013 г. на Wayback Machine . Глоссарий связанных терминов
- ^ Чалмерс, Дэвид Дж. (июнь 1998 г.), «Что такое нейронный коррелят сознания?», Метцингер, Томас (редактор), «Нейронные корреляты сознания: эмпирические и концептуальные вопросы» , MIT Press (опубликовано в сентябре 2000 г.), ISBN 978-0-262-13370-8
- ^ Сквайр 2008 , с. 1223.
- ^ Кандель 2007 , с. 382.
- ^ Шварц, Джеффри М .; Стэпп, Генри П .; Борегар, Марио . «Квантовая физика в нейробиологии и психологии: нейрофизиологическая модель взаимодействия разума и мозга» (PDF) .
- ^ См . Чалмерс 1998 , доступно в Интернете.
- ^ Земан 2001
- ^ Шифф 2004 г.
- ^ Лорейс, Trends Cogn Sci, 2005, 9:556–559.
- ^ Тонони и др. 2016 год
- ^ Ким и Блейк 2004 г.
- ^ Кох 2004 , Рисунок 16.1 Бистабильный куб Неккера , стр. 16.1. 270.
- ^ Логотетис 1998 г.
- ^ Рис и Фрит 2007
- ^ Хейнс и Рис, 2005 г.
- ^ Ли и др. 2007 год
- ^ Симоно и Ники, 2013 г.
- ^ Jump up to: а б Крик и Кох, 1995 г.
- ^ Леопольд и Логотетис, 1996 г.
- ^ Шейнберг и Логотетис 1997.
- ^ Крейман и др. 2002 г.
- ^ Koch 2004 , Рисунок 5.1. Система холинергической поддержки, с. 92. См. главу 5, доступную в Интернете.
- ^ Оуэн и др. 2006 г.
- ^ Лорейс 2005
- ^ Блюменфельд и др. 2004 г.
- ^ Кох 2004 , с. 92
- ^ Виллабланка 2004
- ^ Книга 1995 г.
- ^ Jump up to: а б Милнер и Гудейл, 1995 г.
- ^ Кох и Крик 2001
- ^ Бейлок и др. 2002 г.
- ^ Фланаган, Оуэн ; Полгер, Том В. (1995). «Зомби и функция сознания». Журнал исследований сознания . 2 : 313–321.
- ^ Розенталь, Дэвид (2008). «Сознание и его функции». Нейропсихология . 46 (3): 829–840. doi : 10.1016/j.neuropsychologia.2007.11.012 . ПМИД 18164042 . S2CID 7791431 .
- ^ Харнад, Стеван (2002). «Тьюринговая неразличимость и слепой часовщик» . В Фетцере, Джеймс Х. (ред.). Сознание развивается . Джон Бенджаминс . Проверено 26 октября 2011 г.
- ^ Фейнберг, Т.Э.; Маллатт, Дж. (2013). «Эволюционное и генетическое происхождение сознания в кембрийский период более 500 миллионов лет назад» . Фронт Психол . 4 : 667. doi : 10.3389/fpsyg.2013.00667 . ПМК 3790330 . ПМИД 24109460 .
- ^ Робинсон, Зак; Мэйли, Кори Дж.; Пиччинини, Гуальтьеро (2015). «Является ли сознание спандрелом?». Журнал Американской философской ассоциации . 1 (2): 365–383. дои : 10.1017/apa.2014.10 . S2CID 170892645 .
- ^ Торп и др. 1996 год
- ^ ВанРуллен и Кох, 2003 г.
- ^ Баарс 1988
- ^ Деэн и др. 2003 г.
- ^ Кох, Кристоф (2004). В поисках сознания: нейробиологический подход . Энглвуд, США, Колорадо: Издательство Roberts & Company. ISBN 978-0-9747077-0-9 .
- ↑ См. предисловие Куни к переизданию статьи Чалмерса: Брайан Куни, изд. (1999). «Глава 27: Столкновение с проблемой сознания». Место ума . Cengage Обучение. стр. 382 и далее . ISBN 978-0534528256 .
- ^ Чалмерс, Дэвид (1995). «К проблеме сознания» . Журнал исследований сознания . 2 (3): 200–219. Архивировано из оригинала 8 марта 2005 г. Проверено 18 января 2019 г. См. также эту ссылку
Ссылки
[ редактировать ]- Адамантидис А.Р., Чжан Ф., Араванис А.М., Дейсерот К. и де Лесеа Л. (2007) Нейральные субстраты пробуждения, исследованные с помощью оптогенетического контроля гипокретиновых нейронов . Природа. продвинутое онлайн-издание.
- Баарс Б.Дж. (1988) Когнитивная теория сознания. Издательство Кембриджского университета: Нью-Йорк, штат Нью-Йорк.
- Сиан Бейлок , Карр Т.Х., МакМахон С. и Старкс Дж.Л. (2002) Когда внимание становится контрпродуктивным: влияние разделенного внимания по сравнению с вниманием, сфокусированным на навыках, на новичка и опытного выполнения сенсомоторных навыков . Дж. Эксп. Психол. Прил. 8:6–16.
- Блюменфельд Х., МакНелли К.А., Вандерхилл С.Д., Пейдж А.Л., Чанг Р., Дэвис К., Норден А.Д., Стоккинг Р., Стадхолм К., Новотный Э.Дж. младший, Зубал И.Г. и Спенсер С.С. (2004) Положительные и отрицательные сетевые корреляции при височной эпилепсии . Цереб. Корт. 14: 892–902.
- Боген Дж. Э. (1995) О нейрофизиологии сознания: I. Обзор. Сознание и познание 4: 52–62.
- Чалмерс, Дэвид Дж. (июнь 1998 г.), «Что такое нейронный коррелят сознания?», Метцингер, Томас (редактор), «Нейронные корреляты сознания: эмпирические и концептуальные вопросы» , MIT Press (опубликовано в сентябре 2000 г.), ISBN 978-0-262-13370-8
- Крик Ф. и Кох К. (1990) На пути к нейробиологической теории сознания. Семинары по неврологии, том 2, 263–275.
- Крик Ф.К. и Кох К. (1995) Знаем ли мы о нейронной активности в первичной зрительной коре? Природа 375: 121–3.
- Деэн С. , Серджент К. и Чанже Дж.П. (2003)Модель нейронной сети, связывающая субъективные отчеты и объективные физиологические данные во время сознательного восприятия. Учеб. Натл. акад. наук. США 100: 8520–5.
- Хейнс Дж. Д. и Рис Г. (2005) Предсказание ориентации невидимых стимулов на основе активности первичной зрительной коры человека . Нат. Неврология. 8: 686–91.
- Кандел Эрик Р. (2007). В поисках памяти: Возникновение новой науки о разуме . WW Нортон и компания. ISBN 978-0393329377 .
- Ким Сай и Блейк Р. (2004) Психофизическая магия: превращение видимого в «невидимое». Тенденции Когн. наук. 9: 381–8.
- Кох, Кристоф (2004). В поисках сознания: нейробиологический подход . Энглвуд, США, Колорадо: Издательство Roberts & Company. ISBN 978-0-9747077-0-9 .
- Кох К. и Крик ФК (2001) О зомби внутри. Природа 411: 893.
- Крейман Г., Фрид И. и Кох К. (2002) Однонейронные корреляты субъективного зрения в медиальной височной доле человека . Проц Натл. акад. наук. США 99: 8378–83.
- Лорейс С. (2005) Нейронный коррелят (не)осознанности: уроки вегетативного состояния . Тенденции Когн. наук. 9: 556–9.
- Ли Ш., Блейк Р. и Хигер DJ (2007) Иерархия корковых реакций, лежащих в основе бинокулярного соперничества . Нат. Неврология. 10: 1048–54.
- Леопольд Д.А. и Логотетис Н.К. (1996) Изменения активности в ранней зрительной коре отражают восприятие обезьян во время бинокулярного соперничества. Природа 379: 549–53.
- Логотетис Н. (1998) Отдельные единицы и сознательное видение. Филос. Пер. Р. Сок. Лонд. Б, 353: 1801–18.
- Милнер А.Д. и Гудейл М.А. (1995)Визуальный мозг в действии. Издательство Оксфордского университета, Оксфорд, Великобритания.
- Оуэн А.М., Клеман М.Р., Боли М., Дэвис М.Х., Лорейс С. и Пикард Дж.Д. (2006) Обнаружение осознания в вегетативном состоянии . Наука 313: 1402.
- Рис Г. и Фрит К. (2007) Методологии выявления нейронных коррелятов сознания . В: Блэквеллский спутник сознания. Велманс М. и Шнайдер С., ред., стр. 553–66. Блэквелл: Оксфорд, Великобритания.
- Шейнберг Д.Л. и Логотетис Н.К. (1997) Роль височных областей коры в организации восприятия . Учеб. Натл. акад. наук. США 94: 3408–13.
- Шифф, Николас Д. (ноябрь 2004 г.), «Неврология нарушенного сознания: проблемы когнитивной нейробиологии», в Газзаниге, Майкл С. (ред.), Когнитивные нейронауки (3-е изд.), MIT Press, ISBN 978-0-262-07254-0
- Шимоно М. и Ники К. (2013) Глобальное картирование сети всего мозга, подчеркивающее бинокулярное соперничество. Связность мозга 3: 212–221.
- Торп С., Файз Д. и Марлот К. (1996) Скорость обработки данных в зрительной системе человека . Природа 381: 520–2.
- Сквайр, Ларри Р. (2008). Фундаментальная нейронаука (3-е изд.). Академическая пресса . п. 1256 . ISBN 978-0-12-374019-9 .
- Тонони Г. (2004) Теория информационной интеграции сознания . БМК Нейронаука. 5: 42–72.
- Тонони, Джулио и Боли, Мелани и Массимини, Марчелло и Кох, Кристоф (2016). «Интегрированная теория информации: от сознания к его физическому субстрату». Обзоры природы Неврология . 17 (5). Издательская группа «Природа»: 450–461. дои : 10.1038/nrn.2016.44 . ПМИД 27225071 . S2CID 21347087 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ВанРуллен Р. и Кох К. (2003) Визуальное избирательное поведение может быть вызвано процессом прямой связи. Дж. Конг. Неврология. 15: 209–17.
- Виллабланка Дж.Р. (2004) Противопоставление функциональной роли переднего мозга и ствола мозга в контроле системы сна и бодрствования . Дж. Сон Рез. 13: 179–208.
- Земан А. (2001) Сознание . Мозг. 7: 1263–1289.
Дальнейшее чтение
[ редактировать ]- Аткинсон А. и др. «Сознание: картирование теоретического ландшафта»
- Чалмерс, Д. (1995). Сознательный разум: в поисках фундаментальной теории . Философия разума. Оксфорд: Издательство Оксфордского университета. ISBN 9780195117899 .
- Крик, Фрэнсис (1994). Удивительная гипотеза: научный поиск души . Справочник Macmillan США. ISBN 978-0-684-19431-8 .
- Докинз, М.С. (1993). Только нашими глазами? Поиски сознания животных . Оксфорд: Издательство Оксфордского университета. ISBN 9780198503200 .
- Эдельман, генеральный директор ; Тонони, Дж. (2000). Сознание: как материя становится воображением . Нью-Йорк: Основные книги. ISBN 9780465013777 .
- Гудейл, Массачусетс ; Милнер, AD (2004). Невидимое зрение: исследование сознательного и бессознательного видения . Оксфорд: Издательство Оксфордского университета. ISBN 978-0-19-856807-0 .
- Кох К. и Хепп К. (2006)Квантовая механика и высшие функции мозга: уроки квантовых вычислений и нейробиологии. Природа 440: 611–2. (Свободно доступен по адресу http://www.theswartzfoundation.org/papers/caltech/koch-hepp-07-final.pdf (2007))
- Логотетис, Северная Каролина; Гуггенбергер, Хайнц; Пелед, Шарон; Паулс, Джон (1999). «Функциональная визуализация мозга обезьяны». Природная неврология . 2 (6): 555–562. дои : 10.1038/9210 . ПМИД 10448221 . S2CID 21627829 .
- Шалл, Дж. « О построении моста между мозгом и поведением ». Ежегодные обзоры по психологии. Том 55. Февраль 2004 г., стр. 23–50.
- Ваас, Рюдигер (1999): «Почему нейронные корреляты сознания хороши, но недостаточны». Антропология и философия Том. 3, стр. 121–141. https://web.archive.org/web/20120205025719/http://www.swif.uniba.it/lei/mind/texts/t0000009.htm