Сексуальная мимикрия
Сексуальная мимикрия возникает, когда один пол имитирует противоположный пол в своем поведении, внешнем виде или химических сигналах.
Это чаще встречается у беспозвоночных, хотя сексуальная мимикрия также наблюдается у позвоночных, таких как пятнистые гиены.
Сексуальная мимикрия обычно используется в качестве стратегии спаривания для получения доступа к партнеру, защитного механизма, позволяющего избегать более доминирующих особей, или стратегии выживания. Это также может быть физическая характеристика, определяющая место человека в обществе. Сексуальная мимикрия используется по-разному у разных видов и является частью их стратегии выживания и размножения.
Примеры внутривидовой сексуальной мимикрии у животных включают пятнистую гиену , некоторые виды рыб, воробьинообразных птиц и некоторые виды насекомых.
Межвидовая сексуальная мимикрия может встречаться и у некоторых видов растений, особенно у орхидей. У растений, использующих сексуальную мимикрию, цветы имитируют брачные сигналы насекомых-опылителей. Эти насекомые привлекают цветы и опыляют их посредством псевдокопуляций или другого сексуального поведения, совершаемого с цветком.
Социальные системы
[ редактировать ]Сексуальная мимикрия может играть роль в развитии социальной системы вида. самый яркий пример этого можно увидеть у пятнистой гиены Возможно , Crocuta crocuta . [ 1 ] Самки гиен всех возрастов обладают псевдомаскулинизированными гениталиями, включая псевдопенис, образованный из клитора, и ложную мошонку, образованную из половых губ. Эти характеристики, вероятно, изначально возникли для снижения уровня внутриполой агрессии, получаемой детенышами и молодыми самками от взрослых самок. Однако эта черта вышла за рамки своего первоначального использования и стала очень актуальной для общения пятнистых гиен. Подчиненные гиены приветствуют доминирующих особей, поднимая пенис или псевдопенис и позволяя доминирующей особи лизать его. Вероятно, первоначально это возникло как средство определения пола подчиненного индивидуума, поскольку псевдопенис в эрегированном состоянии менее похож на настоящий пенис, а дегустация этой области может позволить обнаружить концентрации гормонов, специфичные для пола. Однако такое поведение продолжают проявлять взрослые рожавшие самки, которые больше не напоминают самцов ни визуально, ни обонятельно.
Системы спаривания
[ редактировать ]У пятнистых гиен единственный способ спариваться самцами с самками - это полное сотрудничество самки из-за пениформного клитора самки. Повышение статуса самцов дало им больше доступа к доминирующим самкам в клане. Недоминантные самки чаще совокупляются с самцами более низкого ранга. [ 2 ] Самкам гиен дорого обходится рождение ребенка через длинный пеновидный клитор. Пуповина имеет длину 12–18 см, длина пути от матки до конца клитора — 40 см. Пуповина часто рвется еще до появления детеныша, что приводит к смерти многих детенышей от гипоксии. Это путешествие вредно не только для детенышей, но и для матери. Ткань клитора иногда разрывается при первых родах, что может быть фатальным для матери. [ 3 ]
Некоторые виды также используют сексуальную мимикрию в качестве стратегии защиты партнера. Охрана партнера — это процесс, при котором представитель вида не позволяет другому представителю того же вида спариваться со своим партнером. Охрана партнера наблюдается у Cotesia Rubecula , паразитической осы из семейства Braconidae, чья система спаривания является полигинной . Самцов привлекают самки посредством феромонов, и они побуждают самок к спариванию посредством вибраций, на которые самка реагирует, принимая определенную позу. Когда самец, совокупившийся с самкой, видит, что другой самец пытается ухаживать за ней, он часто занимает восприимчивую позицию самки. Посткопуляционная мимикрия самки со стороны самца дает преимущество, действуя как механизм защиты партнера. Если второй самец прибудет достаточно скоро после совокупления самки с первым самцом, второй самец сможет вызвать второе совокупление, которое будет конкурировать с первым. Однако если первый самец, совокупившийся с ней, имитирует самку, это отвлекает второго самца на достаточно долгое время, и самка становится невосприимчивой. [ 4 ]
Подлый совокупление
[ редактировать ]

Скрытое совокупление — это стратегия, используемая многими водными организмами, изображающими сексуальную мимикрию. Несколько исследований показали, что маленькие самцы рыб будут выглядеть и вести себя как самки своего вида, чтобы получить доступ к территории самок и совокупиться с ними. [ 5 ] [ 6 ] В семействе рыб Blenniidae самка Salaria pavo демонстрирует особую окраску и движение, когда хочет приблизиться к самцу и совокупиться с ним. Самец охраняет территорию, а когда самка откладывает яйца, самец-родитель защищает эту территорию, пока из яиц не вылупятся яйца. Самцы второго типа, самцы-тапки, ведут паразитический образ жизни и напоминают самок бленийид своими небольшими размерами, цветом и характером движений. Это позволяет им проникнуть в гнездо, охраняемое родительскими самцами. Самцы-кеды подходят к гнездам с теми же окрасками и движениями, что и самки. В большинстве случаев самцы-кроссовки наблюдаются, когда в гнезде уже находится самка, хотя иногда рыба-кроссовка входит в гнездо вместе с самкой. Этот вид рыб выделяет сперму до того, как самка выпустит икру в воду. [ 7 ] позволяя рыбе-тапке оплодотворить икру, даже если самки нет в гнезде. [ 5 ]

В семействе Сепиина у каракатицы гигантской Sepia apama есть несколько крупных самцов, способных охранять гнездо самки, в то время как другие самцы маленькие и напоминают самок, чтобы красться во время спаривания. У гигантской каракатицы самец ухаживает за самкой и переносит ее сперму в мешочек под клювом самки. Во время этого процесса на теле самки появляются черные пятна на белом фоне. После откладки яиц самец охраняет гнездо от возможных претендентов и противников. «Вторую самку» иногда можно увидеть во время взаимодействия мужчины и женщины в непосредственной близости от пары. Эта каракатица, похожая на самку, имеет такие же черные пятна, как и настоящая самка. Если самец уходит сражаться с другими самцами, эта особь приближается к самке и совокупляется с ней, обычно успешно. Однако в отсутствие соперников эти самцы, «имитирующие самку», демонстрируют фенотип зрелого самца. [ 6 ]
Сексуальная мимикрия против агрессии
[ редактировать ]Аналогичное явление с самцами рыбы-тапки наблюдается у темнокрайного расщепленника Girardinichthys multiradiatus . Молодые особи напоминают беременных самок этого вида, имея темное пятно возле вентиляционного отверстия. Однако в этом случае имитирующие самцы обладают способностью напоминать самок или становиться морфологически зрелыми самцами на протяжении большей части своей взрослой жизни. Это темное пятно позволяет самцам, похожим на самок, избежать агрессии со стороны более доминирующих самцов, а также снижает вероятность того, что самка поблизости сбежит из-за настойчивых ухаживаний самцов. Взрослые самцы не нападают на подчиненную рыбу, и подчиненная рыба сама решает, когда начинать драку, что дает ей преимущество, поскольку половозрелый самец этого не ожидает. Темное пятно также обеспечивает доступ подчиненных самцов к самкам, что является преимуществом, поскольку яйца самок могут быть оплодотворены только в течение пятидневного периода оплодотворения. [ 8 ]


У птиц также наблюдается сексуальная мимикрия, позволяющая избежать агрессии. У некоторых видов птиц на втором году жизни самцы имеют окраску оперения, похожую на самку (самцы SY). Эти самцы SY являются половозрелыми и способны к размножению, но их морфология сильно отличается от более старых самцов после второго года жизни (ASY). Различные исследования изучали эту задержку созревания оперения (DPM) и обнаружили, что DPM у самцов SY снижает агрессию со стороны самцов ASY. [ 9 ] [ 10 ] [ 11 ] Самка мимикрии у птиц впервые была обнаружена у европейской мухоловки-пеструшки Ficedula hypoleuca . Когда поблизости находится самец тусклого цвета, взрослые самцы снижают свою агрессивность и ведут себя так, как будто злоумышленником является самка. Тусклое оперение наблюдается в основном у молодых самцов, вероятно, из-за того, что они родились позже предыдущей весной. Сходство с самками приносит пользу этим молодым самцам при попытке занять территорию, где уже присутствует много самцов, поскольку молодые самцы могут получить информацию и доступ к территории, которая в противном случае была бы для них недоступна. [ 10 ]
Непохожесть на самца обходится дорого, когда дело доходит до защиты территории или привлечения партнера. Самки проявляют агрессию по отношению к самцам тусклой окраски, что затрудняет им спаривание. [ 10 ] Однако у DPM есть некоторые преимущества: как упоминалось выше, он снижает агрессию со стороны пожилых мужчин. [ 10 ] Кроме того, эти птицы, похожие на самок, могут получить доступ к территориям, партнерам и пище, которые в противном случае могли бы быть им недоступны. Еще одним преимуществом является то, что DPM обеспечивает птицам SY более длительную продолжительность жизни; поскольку им не приходится конкурировать с другими самцами, уровень их смертности ниже. Однако это преимущество приносит пользу только особям видов, которые имеют более длительную потенциальную продолжительность жизни, и, следовательно, DPM не принесет пользы короткоживущим видам. [ 9 ] Это известно как гипотеза порога размножения и гласит, что самцам SY следует откладывать размножение только в том случае, если существует большая разница в смертности между самцами SY, которые пытаются размножаться, и теми, кто этого не делает. [ 9 ]

Большинство исследований рассматривали ДПМ как разновидность сексуальной мимикрии. [ 9 ] [ 10 ] это делается путем обмана: самцы птиц ASY не должны различать самок и самцов SY. Однако Мухетер и др. (1997) обнаружили, что территориальные самцы воспринимают самцов тусклой окраски как самцов, но они проявляют меньшую агрессию, поскольку их тусклое оперение снижает конкурентоспособность. Они назвали это честной передачей сигналов , а не сексуальной мимикрией. [ 11 ]
Другой пример сексуальной мимикрии встречается у плоской ящерицы Бродли, Platysaurus Broadleyi , у которой некоторые самцы имитируют самок. Самцы плоских ящериц, как правило, территориальны и агрессивны по отношению к другим самцам. Поэтому некоторым самцам выгодно подражать самкам, чтобы избежать агрессивных встреч и свободно перемещаться по территории самца в поисках партнёров. В этой популяции есть два типа мужчин; самцы-самцы, имитирующие самок, и самцы-самцы, похожие на самцов. Самцы могут визуально обмануть самцов, заставив их поверить в то, что они женщины, из-за их женской морфологии. Однако самцы не могут обмануть самцов с помощью запаха, поскольку самцы могут обнаружить разницу. Поэтому наиболее успешными самцами являются те самцы, которые избегают тесного контакта с другими самцами, тем самым снижая шансы обнаружения по химическим сигналам. [ 12 ]
Молекулярный контроль над сексуальной мимикрией
[ редактировать ]У вида токущих куликов, ерша , или Philomachus pugnax , есть три отдельные морфы самцов: независимые самцы, первичная морфа с темными оперениями, которая устанавливает и агрессивно защищает территорию, на которой токует ток, самцы-спутники, морфа со светлым оперением, которая не защищает свою собственную территорию, но ищет выдающихся независимых самцов для показа рядом с ними, а самцы-федеры, у которых нет типичного самца для размножения оперения, вместо этого они более похожи на самок, обладают меньшим размером тела, промежуточным между типичным самцом и самкой, и которые не занимают последовательно одну территорию, но свободно перемещаются между различными независимыми территориями самцов, возможно, из-за своего женского оперения, и участвуют в тайных совокуплениях с самками. [ 13 ]
Когда рифам (самкам ершей) вводят тестостерон, у рифов выражено ухаживающее поведение самцов и окраска перьев самцов. Тестостерон в этом случае проявляет характеристики, ограниченные полом, воздействуя на единственный аутосомный ген. [ 14 ]

Другой пример можно наблюдать у взрослых самок плодовых мух Drosophila melanogaster , которые очень привлекательны, но уровень их привлекательности снижается вдвое или более после трех минут спаривания. [ 15 ] Во время ухаживания самцы выделяют в самку соединение 7-трикозен, которое снижает ее привлекательность. Однако исследователь обнаружил, что самки также выделяют это соединение через шесть часов после спаривания. Это соединение снижает уровень привлекательности самки как во время ухаживания самца, так и во время спаривания. Таким образом, самка имитирует самца и с помощью этого соединения снижает уровень своей привлекательности. [ 16 ]
Генетический контроль над сексуальной мимикрией
[ редактировать ]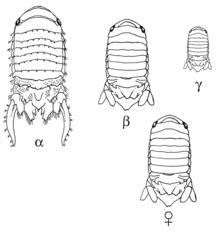
Сексуальная мимикрия некоторых организмов генетически детерминирована определенными аллелями. У морских изопод Paracerceisculpta есть три различные морфологии самцов: альфа-самец — самая крупная морфа, он созревает последним и именно он получает привилегированный доступ к самкам. Бета-самец среднего размера и имитирует самку, чтобы получить доступ к самкам. Наконец, гамма-самец — самая маленькая морфа, и он вторгается в гаремы, куда самки спариваются с альфа-самцами, чтобы получить возможность спариваться. Эта морфология связана с одним аутосомным геном и тремя различными аллелями. Бета является наиболее доминантным аллелем, за ним следует гамма, за которой следует альфа. Отбор по этим аллелям действует в соответствии с равновесием Харди-Вайнберга, и успех спаривания одинаков для всех трех морф. [ 17 ]
Альфа-самцы, гомозиготные по альфа-аллелю, спариваются со многими самками в гареме. Самки предпочитают объединяться с другими самками в гареме, что дает альфа-самцу больший выбор партнеров для спаривания. Шустер (1992) изучил поведение и отношения каждой морфы по отношению к гарему и обнаружил, что бета- и гамма-самцы могут находить гаремы, в которых есть сексуально восприимчивые самки. Они также смогли отличить гарем с сексуально восприимчивой самкой, т. е. той, которая способна к спариванию, и несексуально восприимчивой самкой, т. е. той, которая уже поместила эмбрион в свою сумку и больше не может спариваться. Хотя до сих пор неясно, как бета-самцы делают это или как работают их стратегии спаривания, альфа-самцы не беспокоят их из-за их подражания самкам: бета-самцы могут привлекать в гарем других самок, поскольку самки любят ходить туда, где другие самки. есть, и это дает альфа-самцам больше партнеров. [ 18 ]
Другой отряд организмов, на сексуальную мимику которых влияет их ДНК, — это Odonata , плотоядные насекомые, известные как стрекозы и стрекозы. У этих видов самка иногда имитирует самца. Внутри вида группы самок различаются по цвету: одна группа имитирует цвет самцов, и они известны как андрохромы. Другие группы будут иметь свою собственную женскую окраску и известны как гинохромы. У Ischnura elegans андрохромы составляют 6-30% женской популяции и их цвет обычно синий, как и у самцов; в некоторых популяциях андрохромы крупнее гинохромов. Этот полиморфизм контролируется аутосомным аллелем, и в некоторых исследованиях изучались причины сохранения полиморфизма. [ 19 ]


Наиболее вероятной теорией сохранения полиморфизма у Odonata является теория зависимости от плотности. [ 20 ] в нем говорится, что при высокой плотности самцов андрохромы не беспокоят самцы и их существованию не угрожает мужское преследование. Эта гипотеза также предполагает, что мужчины не могут отличить андрохромов от других мужчин. Однако этому преимуществу противодействует тот факт, что у них не будет много возможностей для спаривания (если таковые имеются), а их воспроизводство ограничено. Эта теория является наиболее вероятным объяснением сохранения полиморфизма, поскольку исследования показали, что андрохромы имеют преимущество в популяциях с высокой плотностью мужчин. [ 21 ]
У меченосца Xiphophorus birchmanni у 40% самцов развивается «ложное пятно беременности», признак, который позволяет самцам имитировать « пятно беременности », обнаруженное у самок. [ 22 ] Ложное пятно беременности вызвано структурными изменениями , которые усиливают экспрессию близлежащего гена -лиганда . [ 22 ] Самцы, пережившие пятно, снизили агрессию со стороны других самцов; однако женщины их презирают, но мужчины уделяют им больше внимания. [ 22 ]
Самоконтроль над сексуальной мимикрией
[ редактировать ]Хотя, как было замечено ранее, большинство организмов, изображающих сексуальную мимикрию, рождаются с такой морфологией/поведением, это не всегда так. Гигантская каракатица Sepia apama , упомянутая выше в разделе «подлые спаривания», рождается со способностью выбирать, изменять ли свою морфологию, чтобы выглядеть как самка или взрослый самец. Когда поблизости не видно конкурентов, каракатица будет выглядеть как взрослый самец и спариваться с самкой. Однако, когда половозрелый самец и самка совокупляются, гигантская каракатица будет напоминать самку и держаться на близком расстоянии от пары, надеясь на шанс спариться с самкой, если взрослый самец уйдет, чтобы сражаться с другими самцами. [ 6 ] Другим примером организма, который способен оставаться маленьким и выглядеть как самка или становиться морфологически зрелым самцом, является расщепленный плавник с темными краями, Girardinichthys multiradiatus . Цель их женской мимикрии была видна раньше, в разделе «сексуальная мимикрия против агрессии», где самцы, выглядящие как женщины, будут избегать агрессии со стороны доминирующих самцов и не позволять самкам покидать их компанию из-за настойчивых ухаживаний самцов. [ 8 ]
Межвидовая обманчивая мимикрия
[ редактировать ]
Межвидовая сексуальная мимикрия также может встречаться у некоторых видов растений. Самый распространенный пример этого известен как опыление с сексуальным обманом и встречается среди некоторых орхидей. [ 23 ] Орхидея имитирует самок своего опылителя, обычно перепончатокрылых, таких как осы и пчелы, привлекая к цветку самцов. Цветы орхидеи имитируют половые феромоны и в некоторой степени внешний вид самок насекомых своего вида-опылителей. Примат обонятельных сигналов над визуальными был продемонстрирован во многих случаях, например, в европейском роде орхидей Ophrys, а также во многих австралийских орхидеях, вводящих в заблуждение сексуально. В некоторых других случаях, таких как южноафриканская маргаритка Gorteria diffusa , зрительные сигналы имеют первостепенное значение. [ 24 ] Визуальные сигналы также повышают привлекательность цветков некоторых видов Ophrys для их опылителей. [ 25 ] [ 26 ] Некоторых самцов ос-сколиид, таких как Campsoscolia ciliata, больше привлекают запахи цветков Ophrys , чем запахи самок ос, хотя оба они привлекают самцов одними и теми же соединениями. Скорее всего, это результат более сильного аромата, исходящего от цветов орхидеи; Самки ос, как правило, издают меньше запаха, чтобы не привлекать хищников. [ 27 ] Независимо от того, используют ли орхидеи внешний вид, аромат или и то, и другое, они имитируют самку-опылителя ради собственной выгоды.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Мюллер, Мартин Н; Рэнгем, Ричард (2002). «Сексуальная мимикрия у гиен» . Ежеквартальный обзор биологии . 77 (1): 3–16. дои : 10.1086/339199 . ISSN 0033-5770 .
- ^ Ист М.Л., Берк Т., Вильгельм К., Крейг С., Хофер Х. (2003). «Сексуальные конфликты у пятнистых гиен: тактика спаривания самцов и самок и их репродуктивный результат в зависимости от возраста, социального статуса и срока владения» . Труды Королевского общества Б. 270 (1521): 1247–54. дои : 10.1098/rspb.2003.2363 . ПМК 1691369 . ПМИД 12816637 .
- ^ Фрэнк Л.Г. (февраль 1997 г.). «Эволюция генитальной маскулинизации: почему у самок гиен такой большой «пенис»?». Тенденции в экологии и эволюции . 12 (2): 58–62. дои : 10.1016/S0169-5347(96)10063-X . ПМИД 21237973 .
- ^ Филд С.А., Келлер М.А. (декабрь 1993 г.). «Альтернативная тактика спаривания и мимикрия самок как посткопулятивное поведение по охране партнера у паразитической осы Cotesia Rubecula». Поведение животных . 46 (6): 1183–1189. дои : 10.1006/anbe.1993.1308 . S2CID 54325074 .
- ^ Jump up to: а б Гонсалвеш Э.Дж., Алмада ВК, Оливейра Р.Ф., Сантос А.Дж. (11 мая 2009 г.). «Женская мимикрия как тактика спаривания у самцов собачки Salaria Pavo». Журнал Морской биологической ассоциации Соединенного Королевства . 76 (2): 529. doi : 10.1017/S0025315400030721 . S2CID 86751010 .
- ^ Jump up to: а б с Норман, доктор медицинских наук, Финн Дж., Трегенза Т. (7 июля 1999 г.). «Женское олицетворение как альтернативная репродуктивная стратегия гигантских каракатиц» . Труды Королевского общества Б. 266 (1426): 1347–1349. дои : 10.1098/rspb.1999.0786 . ПМК 1690068 .
- ^ Патцнер Р.А. (1984). «Размножение Blennius pavo (Teleostei Bleniidae). II. Строение поверхности созревшего яйца». Зоологический обзор . 213 : 44–50.
- ^ Jump up to: а б Масиас-Гарсия К, Валеро А (19 мая 2010 г.). «Контекстно-зависимая сексуальная мимикрия у живородящих рыб». Этология Экология и эволюция . 13 (4): 331–339. дои : 10.1080/08927014.2001.9522764 . S2CID 84161470 .
- ^ Jump up to: а б с д Стадд М.В., Робертсон Р.Дж. (1985). «Продолжительность жизни, конкуренция и задержка созревания оперения у самцов воробьинообразных: гипотеза порога размножения». Американский натуралист . 126 (1): 101–115. дои : 10.1086/284399 . JSTOR 2461565 . S2CID 85348250 .
- ^ Jump up to: а б с д и Слагсволд Т., Саэтре ГП (июнь 1991 г.). «Эволюция цвета оперения самцов мухоловки-пеструшки ( Ficedula Hyopleuca ): свидетельства женской мимикрии» . Эволюция . 45 (4): 910–917. дои : 10.1111/j.1558-5646.1991.tb04359.x . ПМИД 28564056 .
- ^ Jump up to: а б Мютер В.Р., Грин Э., Рэтклифф Л. (27 октября 1997 г.). «Замедленное созревание оперения у лазуритовых овсянок: проверка гипотез женской мимикрии и сигнализации статуса». Поведенческая экология и социобиология . 41 (4): 281–290. дои : 10.1007/s002650050389 . S2CID 22253297 .
- ^ Уайтинг М.Дж., Уэбб Дж.К., Кио Дж.С. (25 февраля 2009 г.). «Женщины-имитаторы плоских ящериц используют сексуальный обман с помощью визуальных, а не химических сигналов» . Труды Королевского общества Б. 276 (1662): 1585–1591. дои : 10.1098/рспб.2008.1822 . ПМК 2660994 . ПМИД 19324828 .
- ^ Куппер, Клеменс; Стокс, Майкл; Риссе, Джудит Э; дос Ремедиос, Натали; Фаррелл, Линдси Л; Макрей, Сьюзен Б; Морган, Тауна С; Карлионова, Наталья; Пинчук Павел; Веркуил, Ивонн I; Китайский, Александр С; Вингфилд, Джон С; Пирсма, Теунис; Цзэн, Кай; Слейт, Джон (январь 2016 г.). «Суперген определяет сильно расходящиеся мужские репродуктивные морфы ерша» . Природная генетика . 48 (1): 79–83. дои : 10.1038/ng.3443 . ISSN 1061-4036 . ПМК 5218575 . ПМИД 26569125 .
- ^ Lank DB, Coupe M, Wynne-Edwards KE (22 ноября 1999 г.). «Вызванные тестостероном мужские признаки у самок ершей (Philomachus pugnax): аутосомное наследование и гендерная дифференциация» . Труды Королевского общества Б. 266 (1435): 2323–2330. дои : 10.1098/rspb.1999.0926 . ПМК 1690456 .
- ^ Томпкинс Л., Холл Дж.К. (январь 1981 г.). «Различное влияние летучих соединений на ухаживание спаренных и девственных самок дрозофилы». Журнал физиологии насекомых . 27 (1): 17–21. дои : 10.1016/0022-1910(81)90026-3 .
- ^ Скотт Д. (ноябрь 1986 г.). «Сексуальная мимикрия регулирует привлекательность спаривающихся самок Drosophila melanogaster» (PDF) . Труды Национальной академии наук Соединенных Штатов Америки . 83 (21): 8429–33. Бибкод : 1986PNAS...83.8429S . дои : 10.1073/pnas.83.21.8429 . ПМЦ 386942 . ПМИД 3095835 .
- ^ Шустер С.М., Уэйд М.Дж. (18 апреля 1991 г.). «Равный успех спаривания среди мужских репродуктивных стратегий морских изопод». Природа . 350 (6319): 608–610. Бибкод : 1991Natur.350..608S . дои : 10.1038/350608a0 . S2CID 4354417 .
- ^ Шустер С.М. (1992). «Репродуктивное поведение α-, β- и γ-самцов морф Paracerceisculpta, морского изоподного ракообразного» (PDF) . Поведение . 121 (3/4): 231–258. дои : 10.1163/156853992X00381 . JSTOR 4535029 .
- ^ Кордеро А., Карбоне С.С., Уцери К. (январь 1998 г.). «Возможности спаривания и затраты на спаривание снижаются у андрохромных самок стрекоз Ischnura elegans (Odonata)». Поведение животных . 55 (1): 185–197. дои : 10.1006/anbe.1997.0603 . ПМИД 9480685 . S2CID 2096830 .
- ^ Хиннекинт Б.О. 1987. Динамика численности населения Ишнуры э. Elegans (Vnader Linden) (Insecta:Odonata) с особым акцентом на морфологические изменения цвета, женский полиморфизм, многолетние циклы и их влияние на поведение . Гидобиология. 146:3-31.
- ^ Кордеро А., Андрес Х.А. (1996). «Цветовой полиморфизм у зубатых: самки, имитирующие самцов?». Журнал Британского общества стрекоз . 12 (2): 50–60.
- ^ Jump up to: а б с Додж, Тристрам О.; Ким, Бернард Ю.; Баченас, Джон Дж.; Банерджи, Шрея М.; Ганн, Тереза Р.; Донни, Алекс Э.; Учитывая, Лайл А.; Райс, Андреас Р.; Кокс, София К. Хаазе (14 мая 2024 г.), Сложные структурные вариации и поведенческие взаимодействия лежат в основе сбалансированного полиморфизма сексуальной мимикрии , doi : 10.1101/2024.05.13.594052 , получено 30 мая 2024 г.
- ^ Шистль Ф.П. (1 июня 2005 г.). «Об успехе аферы: опыление обманом у орхидей» . Naturwissenschaften . 92 (6): 255–264. Бибкод : 2005NW.....92..255S . дои : 10.1007/s00114-005-0636-y . hdl : 20.500.11850/32223 . ПМИД 15931514 . S2CID 24626163 .
- ^ Эллис А.Г., Джонсон С.Д. (ноябрь 2010 г.). «Цветочная мимикрия усиливает экспорт пыльцы: эволюция опыления путем сексуального обмана за пределами орхидных». Американский натуралист . 176 (5): E143–E151. дои : 10.1086/656487 . ПМИД 20843263 . S2CID 45076899 .
- ^ Гаскетт AC, Герберштейн ME (2 октября 2009 г.). «Цветовая мимикрия и сексуальный обман орхидей Язык (Cryptostylis)» . Naturwissenschaften . 97 (1): 97–102. дои : 10.1007/s00114-009-0611-0 . ПМИД 19798479 . S2CID 1729573 .
- ^ Сингер Р.Б., Флах А., Келер С., Марсайоли А.Дж., Амарал Мдо С. (июнь 2004 г.). «Сексуальная мимикрия у Mormolyca Ringens (Lindl.) Schltr. (Orchidaceae: Maxillariinae)» . Анналы ботаники . 93 (6): 755–62. дои : 10.1093/aob/mch091 . ПМЦ 4242296 . ПМИД 15051623 .
- ^ Аяс М., Шистль Ф.П., Паулюс Х.Ф., Ибарра Ф., Франке В. (7 марта 2003 г.). «Привлечение опылителей к сексуально обманчивой орхидее с помощью нетрадиционных химикатов» . Труды Королевского общества Б. 270 (1514): 517–522. дои : 10.1098/rspb.2002.2271 . ПМЦ 1691269 . ПМИД 12641907 .
