Пинеалоцит
| Пинеалоцит | |
|---|---|
 Поперечное сечение шишковидной железы, на котором видны пинеалоциты и другие клетки. | |
| Подробности | |
| Система | Эндокринная система |
| Расположение | Шишковидная железа |
| Идентификаторы | |
| латинский | пинеалоцит, эндокриноцит шишковидной железы |
| ТД | Х3.08.02.3.00002 |
| ФМА | 83417 |
| Анатомические термины микроанатомии | |
Пинеалоциты являются основными клетками шишковидной железы , расположенной за третьим желудочком и между двумя полушариями головного мозга . Основная функция пинеалоцитов — секреция гормона мелатонина , играющего важную роль в регуляции циркадных ритмов . [1] У людей супрахиазматическое ядро гипоталамуса . передает сообщение темноты пинеалоцитам и, как следствие, контролирует дневной и ночной цикл [2] Было высказано предположение, что пинеалоциты происходят из фоторецепторных клеток . [3] [4] Исследования также показали снижение количества пинеалоцитов путем апоптоза по мере увеличения возраста организма. [5] Существует два разных типа пинеалоцитов, тип I и тип II, которые классифицируются на основе определенных свойств, включая форму, наличие или отсутствие свертки ядерной оболочки и состав цитоплазмы .
Виды пинеалоцитов
[ редактировать ]Пинеалоциты 1 типа
[ редактировать ]Пинеалоциты типа 1 также известны как светлые пинеалоциты, поскольку при просмотре под световым микроскопом они окрашиваются с низкой плотностью и кажутся человеческому глазу светлее. В ходе исследований было выявлено, что эти клетки типа 1 имеют круглую или овальную форму и диаметр в пределах 7–11 микрометров. [6] Пинеалоциты 1-го типа обычно более многочисленны как у детей, так и у взрослых, чем пинеалоциты 2-го типа. [6] Они также считаются более активными клетками из-за наличия определенного клеточного содержимого, включая высокую концентрацию митохондрий . [7] Еще одним фактом, согласующимся с пинеалоцитами 1-го типа, является увеличение количества лизосом и плотных гранул, присутствующих в клетках, по мере увеличения возраста организма, что, возможно, указывает на важность аутофагоцитоза в этих клетках. [6] Исследования также показали, что пинеалоциты 1-го типа содержат нейромедиатор серотонин , который позже превращается в мелатонин , основной гормон, секретируемый шишковидной железой . [8]
Пинеалоциты 2 типа
[ редактировать ]Пинеалоциты 2-го типа также известны как темные пинеалоциты, поскольку при просмотре под световым микроскопом они окрашиваются с высокой плотностью и кажутся более темными для человеческого глаза. Как показали исследования и микроскопия, они представляют собой круглые, овальные или удлиненные клетки диаметром около 7–11,2 микрометров. [6] Ядро шероховатой пинеалоцита 2 типа содержит множество складок, которые содержат большое количество эндоплазматической сети и рибосом . [6] обилие ресничек и центриолей также обнаружено В клетках 2-го типа шишковидной железы . [7] Уникальным для типа 2 является наличие вакуолей, содержащих 2 слоя мембраны. [7] Поскольку клетки типа 1 содержат серотонин , клетки типа 2 содержат мелатонин и, как полагают, имеют сходные характеристики с эндокринными и нейрональными клетками. [8]
Синаптические ленты
[ редактировать ]Синаптические ленты — это органеллы , видимые в пинеалоцитах с помощью электронной микроскопии . Синаптические ленты обнаруживаются в пинеалоцитах как у детей, так и у взрослых, но не обнаруживаются у плодов человека. [6] Исследования на крысах позволили получить больше информации об этих органеллах. Характерным белком синаптических лент является RIBEYE, выявленный методами световой и электронной микроскопии. [9] У низших позвоночных синаптические ленты служат фоторецепторным органом, а у высших позвоночных они выполняют секреторные функции внутри клетки. Присутствие таких белков, как Munc13-1, указывает на то, что они важны для высвобождения нейромедиаторов . [9] Ночью синаптические ленты крыс кажутся крупнее и слегка изогнутыми, но днем они кажутся меньшими и палочковидными. [9]
Эволюция пинеалоцитов
[ редактировать ]Распространенная теория эволюции пинеалоцитов состоит в том, что они произошли от фоторецепторных клеток . Предполагается, что у предков позвоночных пинеалоциты выполняли ту же функцию, что и фоторецепторные клетки, такие как клетки сетчатки; у многих позвоночных, не являющихся млекопитающими, шишковидные клетки сетчатки все еще активно фоторецепторны, хотя эти клетки не вносят вклад в формирование зрительного изображения. [10] [11] Между этими двумя типами клеток существует структурное, функциональное и генетическое сходство. Структурно оба развиваются из области мозга, называемой промежуточным мозгом , а также области, содержащей таламус и гипоталамус , во время эмбриологического развития. [3] Оба типа клеток имеют схожие особенности, включая реснички , складчатые мембраны и полярность . [4] Функциональные доказательства этой теории эволюции можно увидеть на примере позвоночных, не относящихся к млекопитающим. Сохранение фоточувствительности пинеалоцитов миног, рыб, амфибий, рептилий и птиц, а также секреция мелатонина некоторыми из этих низших позвоночных позволяют предположить, что пинеалоциты млекопитающих, возможно, когда-то служили фоторецепторными клетками. [3] [4] Исследователи также указали на наличие нескольких белков-фоторецепторов, обнаруженных в сетчатке в пинеалоцитах кур и рыб. [3] Генетические данные показывают, что гены фототрансдукции, экспрессируемые в фоторецепторах сетчатки, также присутствуют в пинеалоцитах. [4]
Еще одним доказательством эволюции пинеалоцитов из фоторецепторных клеток является сходство ленточных комплексов в клетках двух типов. Присутствие белка RIBEYE и других белков как в пинеалоцитах, так и в сенсорных клетках (как в фоторецепторах, так и в волосковых клетках) предполагает, что эти две клетки эволюционно связаны друг с другом. [9] Различия между двумя синаптическими лентами существуют в наличии определенных белков, таких как ERC2 /CAST1, и в распределении белков внутри комплексов каждой клетки. [9]
Мелатонин
[ редактировать ]
Регулирование
[ редактировать ]Регуляция синтеза мелатонина важна для основной функции мелатонина в циркадных ритмах . Основным механизмом молекулярного контроля секреции мелатонина у позвоночных является фермент AANAT (арилалкиламин-N-ацетилтрансфераза). Экспрессия гена AANAT контролируется транскрипционным фактором pCREB, и это становится очевидным, когда клетки, обработанные эпиталоном, пептидом, который влияет на транскрипцию pCREB, приводят к увеличению синтеза мелатонина. [8] AANAT активируется через систему протеинкиназы А , в которой циклический АМФ (цАМФ). участвует [4] Активация AANAT приводит к увеличению выработки мелатонина. [4] Хотя существуют некоторые различия, специфичные для определенных видов позвоночных, влияние цАМФ на AANAT и AANAT на синтез мелатонина остается довольно постоянным. [4]
Синтез мелатонина также регулируется нервной системой. Нервные волокна ретиногипоталамического тракта соединяют сетчатку с супрахиазматическим ядром (SCN). SCN стимулирует высвобождение норадреналина из симпатических нервных волокон верхних шейных ганглиев , которые синапсируют с пинеалоцитами. [1] [4] Норадреналин вызывает выработку мелатонина в пинеалоцитах, стимулируя выработку цАМФ. Поскольку высвобождение норадреналина из нервных волокон происходит ночью, эта система регуляции поддерживает циркадные ритмы организма. [1]
Синтез
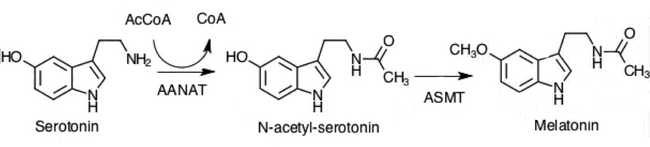
[ редактировать ]Пинеалоциты синтезируют гормон мелатонин, сначала превращая аминокислоту триптофан в серотонин. Затем серотонин ацетилируется ферментом AANAT и превращается в N-ацетилсеротонин . N-ацетилсеротонин превращается в мелатонин с помощью фермента гидроксииндол-О-метилтрансферазы (HIOMT), также известного как ацетилсеротонин-O-метилтрансфераза (ASMT). [1] Активность этих ферментов высока в ночное время и регулируется ранее обсуждавшимися механизмами с участием норадреналина. [1]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и Панди-Перумал С.Р., Шринивасан В., Маэстрони Г.Дж., Кардинали Д.П., Поггелер Б., Харделанд Р. (июль 2006 г.). «Мелатонин: самый универсальный биологический сигнал природы?» . Журнал ФЭБС . 273 (13): 2813–38. дои : 10.1111/j.1742-4658.2006.05322.x . ПМИД 16817850 .
- ^ Маронде Э., Штеле Дж.Х. (2007). «Шишковидная железа млекопитающих: известные факты, неизвестные аспекты». Тенденции в эндокринологии и обмене веществ . 18 (4): 142–9. дои : 10.1016/j.tem.2007.03.001 . ПМИД 17374488 . S2CID 20907798 .
- ^ Jump up to: а б с д Мано Х., Фукада Ю. (2006). «Срединный третий глаз: шишковидная железа прослеживает эволюцию фоторецепторных органов позвоночных» . Фотохимия и фотобиология . 83 (1): 11–8. doi : 10.1562/2006-02-24-IR-813 . ПМИД 16771606 . S2CID 13037403 .
- ^ Jump up to: а б с д и ж г час Кляйн, округ Колумбия (2006). «Эволюция шишковидной железы позвоночных: гипотеза AANAT». Хронобиология Интернэшнл . 23 (1–2): 5–20. дои : 10.1080/07420520500545839 . ПМИД 16687276 . S2CID 29845507 .
- ^ Полякова В.О., Линькова Н.С., Пичугин С.А. (февраль 2011 г.). «Изменения апоптоза и пролиферации клеток шишковидной железы человека при старении». Бюллетень экспериментальной биологии и медицины . 150 (4): 468–70. дои : 10.1007/s10517-011-1170-x . ПМИД 22268045 . S2CID 19539906 .
- ^ Jump up to: а б с д и ж Аль-Хусейн С.М. (август 2006 г.). «Пинеалоциты шишковидной железы человека: исследование световой и электронной микроскопии». Фолиа Морфологическая . 65 (3): 181–7. ПМИД 16988913 .
- ^ Jump up to: а б с Кальво Дж., Бойя Дж. (май 1984 г.). «Ультраструктура шишковидной железы взрослой крысы» . Журнал анатомии . 138 (Часть 3) (3): 405–9. ПМЦ 1164325 . ПМИД 6735903 .
- ^ Jump up to: а б с Хавинсон, В.Х.; Линькова, Н.С.; Кветной Игорь Михайлович ; Кветная, ТВ; Полякова, В.О.; Корф, HW (2012). «Молекулярно-клеточные механизмы пептидной регуляции синтеза мелатонина в культуре пинеалоцитов». Бюллетень экспериментальной биологии и медицины . 153 (2): 255–58. дои : 10.1007/s10517-012-1689-5 . ПМИД 22816096 . S2CID 22047170 .
- ^ Jump up to: а б с д и Спивокс-Беккер И., Маус С., Том Дик С., Фейтова А., Энгель Л., Воллочек Т., Вольфрум Ю., Воллрат Л., Шпессерт Р. (август 2008 г.). «Белки активной зоны динамически связаны с синаптическими лентами в пинеалоцитах крысы» . Исследования клеток и тканей . 333 (2): 185–95. дои : 10.1007/s00441-008-0627-3 . ПМЦ 2757586 . ПМИД 18523806 .
- ^ Пу Г.А., Даулинг Дж.Э. (ноябрь 1981 г.). «Анатомо-физиологические характеристики фоторецепторных клеток шишковидной железы личинки миноги Petromyzon marinus». Журнал нейрофизиологии . 46 (5): 1018–38. дои : 10.1152/jn.1981.46.5.1018 . ПМИД 7299444 .
- ^ Кавано-Ямасита Э., Коянаги М., Ситида Ю., Оиси Т., Тамоцу С., Теракита А. (январь 2011 г.). Барнс С. (ред.). «β-аррестин функционально регулирует неотбеливающий пигмент парапинопсин в шишковидной железе миноги» . ПЛОС ОДИН . 6 (1): e16402. Бибкод : 2011PLoSO...616402K . дои : 10.1371/journal.pone.0016402 . ПМК 3031554 . ПМИД 21305016 .
Внешние ссылки
[ редактировать ]- Гистологическое изображение: 14402loa - Система обучения гистологии в Бостонском университете.