Гипотеза родного языка




Гипотеза отцовского языка предполагает, что люди склонны говорить на языке своего отца. Гипотеза основана на предположении 1997 года о том, что языковая принадлежность более тесно коррелирует с вариациями Y-хромосомы, чем с вариациями митохондриальной ДНК . Первоначальная работа была проведена на африканских и европейских образцах группой популяционных генетиков под руководством Лорана Экскофье. На основе этих и аналогичных результатов других генетиков исторический лингвист Джордж ван Дрим в 2010 году разработал гипотезу о том, что обучение матерью языка своего супруга своим детям является механизмом, с помощью которого язык преимущественно распространяется с течением времени. . Сосредоточение внимания на доисторическом языковом сдвиге в уже заселенных районах, примеры по всему миру. [1] показывают, что всего лишь 10–20% доисторической мужской иммиграции могут (но не обязательно) вызвать смену языка, что указывает на навязывание элиты, подобное тому, которое могло произойти с появлением первых фермеров или рабочих-металлистов в эпоху неолита, бронзы и железного века. .
Раннее аутосомное исследование
[ редактировать ]До открытия вариаций мтДНК и вариаций Y-хромосомы в 1980-х и 1990-х годах соответственно [2] [3] в популяционной генетике невозможно было отличить мужские и женские эффекты. Вместо этого исследователям пришлось полагаться на аутосомную изменчивость, начиная с первого популяционно-генетического исследования с использованием групп крови , проведенного Людвиком Хиршфельдом в 1919 году. [4] Позже стали использоваться и другие генетические полиморфизмы , например полиморфизмы белков плазмы крови , полиморфизмы антигенов лимфоцитов человека или полиморфизмы иммуноглобулинов . [5] На этом основании иногда предлагались корреляции между языками и генетическими вариациями. [6] [7] но вопросы, специфичные для пола, не могли быть решены до 1990-х годов, когда для изучения стали доступны как мтДНК, так и вариации Y-хромосомы у людей.
Происхождение гипотезы
[ редактировать ]Y-хромосома наследуется по отцовской линии , то есть передается только среди мужчин, от отца к сыну. Митохондриальная ДНК, с другой стороны, наследуется по материнской линии , то есть она передается только от матери к ее детям и от ее дочерей к своим детям. В 1997 году Лоран Экскофье, его ученица Эстелла Полони и его команда сообщили, что они обнаружили сильную корреляцию между вариацией Y-хромосомной последовательности P49a,f/Taql и лингвистикой, но не смогли найти такого соответствия для вариации мтДНК. Полони и др. предложил возможные последствия такой корреляции, т.е. гипотезу «отцовского языка»:
- «Как следствие, женское разнообразие нашего генома будет хуже сочетаться с географией и лингвистикой, чем наш мужской компонент. [...] Если это окажется так, то общее убеждение, что мы говорить на родном языке следует пересмотреть в пользу концепции «отцовского языка». [8]
Эстелла Полони также представила гипотезу «отцовского языка» на международной конференции в Париже в апреле 2000 года. [9]
На основе этой популяционно-генетической работы исторический лингвист Джордж ван Дрим разработал гипотезу «отцовского языка» в своих этнолингвистических публикациях и публикациях по популяционной генетике, соавтором которых он является. [10] На конференции Индо-Тихоокеанской ассоциации предыстории в Тайбэе в 2002 году он предположил, что
- «Мать, обучающая своих детей языку отца, была повторяющейся, повсеместной и распространенной моделью на протяжении всей лингвистической истории, […] некоторые механизмы изменения языка с течением времени, вероятно, присущи динамике этого пути передачи. Такие корреляции наблюдаются во всем мире». [11]
Открытие Y-хромосомных маркеров языков
[ редактировать ]Следующим событием стало открытие специфических маркеров Y-хромосомы, связанных с языком. [12] [13] [14] Эти варианты Y-хромосомы не вызывают изменения языка, но оказались переносчиками исторических или доисторических носителей мужского пола, распространявших язык. Эти специфичные для языка маркеры Y-хромосомы создают корреляции, подобные тем, которые наблюдали Poloni et al. 1997 г., и, кроме того, позволяют определить географическую протяженность, временную глубину и уровень мужской иммиграции, лежащие в основе незарегистрированного (доисторического) языкового изменения.
Примеры родных языков
[ редактировать ]
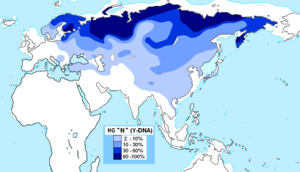
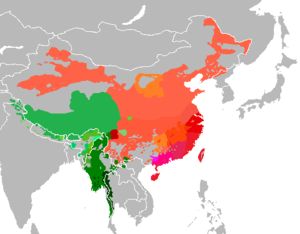

Есть несколько ярких примеров, когда доисторическое распространение языковой семьи сильно коррелирует с распространением гаплогрупп Y-хромосомы. [1]
- Предполагается, что расселение индоевропейцев с предполагаемой родины в Понтийско-Каспийской степи согласно курганской гипотезе связано с распространением субклада гаплогруппы R , R1a1, в Европу. [12] R1a1 также может отражать приход индоариев в северную Индию. [10] [15]
- Y-хромосомная линия L потенциально может отражать более раннее патрилингвальное распространение эламо-дравидийцев, происходящее из региона современного Ирана. [15] Однако эламо-дравидийское предложение по-прежнему отвергается ведущими лингвистами. [ нужна ссылка ]
- Австроазиатские носители демонстрируют высокую частоту субклада гаплогруппы O2a . Например, носители мунда на севере и северо-востоке Индии демонстрируют высокие частоты O2a, чего нет у их региональных соседей, говорящих на других языках, кроме австроазиатских, в то время как их гаплогруппы мтДНК, похоже, часто встречаются в их регионе, независимо от языковой принадлежности. [13]
- Популяционно-генетическое исследование 23 ханьских китайцев. популяций [16] показало, что ханьская экспансия на юг во время китаизации того, что сегодня является южным Китаем, была преимущественно ориентирована на мужчин и является бесспорным примером гипотезы отцовского языка. [10]
- Также было высказано предположение, что банту и другие нигеро-конголезские языки хорошо коррелируют с гаплогруппами Y-хромосомы. [17] [18]
- Распространение афроазиатских языков было связано с расширением гаплогруппы E1b1b . [19]
Подразумеваемое
[ редактировать ]Гипотеза «отцовского языка» имеет далеко идущие последствия для нескольких процессов в лингвистике, таких как изменение языка , овладение языком и социолингвистика . Гипотеза «отцовского языка» также имеет значение для овладения языком, поскольку эта гипотеза предлагает эволюционное объяснение того, почему женщины могут быть лучше в некоторых аспектах языковой деятельности и усвоения. [20] [21] [22] [23] [24] [25] [26] [27] [28]
Исторический лингвист Джордж ван Дрим интерпретировал корреляцию гаплогрупп Y-хромосомы и языковых семей как указание на то, что распространение языковых семей часто было опосредовано миграцией по признаку мужского пола, независимо от того, были ли эти вторжения военными или чем-то менее впечатляющим. Он предположил, что большинство языковых сообществ говорят на родных языках, а не на родных. [15]
Гипотеза «отцовского языка» имеет значение для понимания лингвистами языковых изменений. Надо полагать, что динамика языковых изменений, посредством которых матери передают язык своих супругов своим потомкам, отличается от динамики языковых изменений в одноязычном сообществе и даже от динамики изменения в двуязычном сообществе, где матери передают язык самостоятельно. язык своим детям. [29] Как следствие, такая динамика может привести к разрыву с прошлым. Например, было замечено, что мичиф , генетически являющийся алгонкинским языком (как и равнинный кри ), был релексифицирован женщинами -метисами с французским метисом , языком их мужей, и поэтому генетическое родство мичифа стало почти неопределяемым. [30] [31] [32] Если процесс релексификации вышел за рамки возможности лингвистической реконструкции , динамика такого процесса может затмить истинное лингвистическое наследие сообщества. [29]
Исключения
[ редактировать ]Генетика не определяет язык, на котором говорит человек, и связь между гаплогруппами Y-хромосомы и языковым родством является наблюдаемой корреляцией, а не причинно-следственной связью . Хотя родные языки преобладают, в мире существуют исключения из них. Двумя очень известными исключениями являются жители Бэлць на севере Пакистана и венгры .Гаплогруппы мтДНК, наиболее часто встречающиеся в Бельцах, такие же, как и в соседних тибетских общинах, тогда как гаплогруппы Y-хромосомы, наиболее часто встречающиеся у мужчин Бэлць, по-видимому, проникли в Балтистан с запада с введением ислама. Бельцы говорят на одном из самых консервативных тибетских языков . [10] Язык бельц соответствует мтДНК, а не Y-хромосоме, и фактически является ярким примером родного языка. [15] Другим известным исключением является венгерский язык. [ сомнительно – обсудить ] . Гаплогруппа N1c Y-хромосомы, отличающаяся делецией Tat-C, часто встречается во всех уральских языковых сообществах, но практически отсутствует у венгерских мужчин. Таким образом, хотя вторжение мадьяр на территорию современной Венгрии исторически засвидетельствовано и оставило четкие лингвистические свидетельства, генетически мадьярское вторжение не оставило заметных генетических следов. Напротив, с генетической точки зрения венгры сильно напоминают западнославянскую языковую общность. [29]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б Форстер, Питер; Ренфрю С. (2011). «Родной язык и Y-хромосомы». Наука . 333 (6048): 1390–1391. Бибкод : 2011Sci...333.1390F . дои : 10.1126/science.1205331 . ПМИД 21903800 . S2CID 43916070 .
- ^ Феррис, SD; Браун В.М., Дэвидсон В.С., Уилсон AC (1981). «Обширный полиморфизм в митохондриальной ДНК обезьян» . Труды Национальной академии наук США . 78 (10): 6319–6323. Бибкод : 1981PNAS...78.6319F . дои : 10.1073/pnas.78.10.6319 . ПМК 349030 . ПМИД 6273863 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Андерхилл, Питер; Джин Л., Лин А.А., Мехди С.К., Дженкинс Т., Воллрат Д., Дэвис Р.В., Кавалли-Сфорца Л.Л., Офнер П.Дж. (1997). «Обнаружение многочисленных биаллельных полиморфизмов Y-хромосомы методом денатурирующей высокоэффективной жидкостной хроматографии» . Геномные исследования . 7 (10): 996–1005. дои : 10.1101/гр.7.10.996 . ПМК 310671 . ПМИД 9331370 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Хиршфельд, Л; Хиршфельд Х (1919). «Попытка применить серологические методы к проблеме рас». Антропология . 29 : 505–537.
- ^ Кавалли-Сфорца, Луиджи Лука; Меноцци П., Пьяцца А (1994). История и география генов человека . Принстон, Нью-Джерси: Издательство Принстонского университета.
- ^ Дарлингтон, компакт-диск (1947). «Генетическая составляющая языка» . Наследственность . 1 (3): 269–286. дои : 10.1038/hdy.1947.18 .
- ^ Кавалли-Сфорца, LL; Пьяцца А, Меноцци П, Маунтин Дж (1988). «Реконструкция эволюции человека: объединение генетических, археологических и лингвистических данных» . Труды Национальной академии наук США . 85 (16): 6002–6006. Бибкод : 1988PNAS...85.6002C . дои : 10.1073/pnas.85.16.6002 . ПМК 281893 . ПМИД 3166138 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Полони, Эстела С; Семино О, Пассарино Дж, Сантакьяра-Бенерекетти А.С., Дюпанлуп И, Лангани А, Экскофье Л (1997). «Генетическое сходство человека с гаплотипами Y-хромосомы p49a,f/TaqI демонстрирует сильное соответствие с лингвистическими данными» . Американский журнал генетики человека . 61 (5): 1015–1035. дои : 10.1086/301602 . ПМК 1716025 . ПМИД 9346874 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Полони, Эстелла Симоне; и др. (2000). «Языки и гены: способы передачи, наблюдаемые посредством анализа генов, специфичных для мужчин и женщин». В Жан-Луи Дессале и Лале Гадакпуре (ред.). Материалы: Эволюция языка, 3-я Международная конференция, 3-6 апреля 2000 г. Париж: Высшая национальная школа телекоммуникаций. стр. 185–186.
- ^ Перейти обратно: а б с д ван Дрим, Джордж (2007). «Австроазиатская филогения и австроазиатская родина в свете недавних популяционно-генетических исследований». Мон-кхмерские исследования . 37 : 1–14.
- ^ ван Дрим, Джордж (2012). «Этнолингвистическая идентичность одомашнителей азиатского риса» (PDF) . Comptes Рендус Палевол . 11 (2): 117–132. Бибкод : 2012CRPal..11..117V . дои : 10.1016/j.crpv.2011.07.004 .
- ^ Перейти обратно: а б Зержал, Татьяна; Пандия А., Сантос Ф.Р., Адхикари Р., Таразона Э., Кайзер М., Евграфов О., Сингх Л., Тангарадж К., Дестро-Бисол Дж., Томас М.Г., Камар Р., Мехди С.К., Россер З.Х., Херлз М.Э., Джоблинг М.А., Тайлер-Смит С (1999). «Использование вариаций Y-хромосомной ДНК для изучения истории населения: недавнее распространение самцов в Азии и Европе». В Суриндер С. Папиха, Ранджан Дека и Ранаджит Чакраборти (ред.). Геномное разнообразие. Применение в генетических исследованиях человеческой популяции. Вильямсбург, Вирджиния, 26 июля — 1 августа 1998 г. Нью-Йорк: Издательство Kluwer Academic/Plenum. стр. 91–101.
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Перейти обратно: а б Чаубей, Гьянешвер; и др. (2010). «Генетическая структура населения у индийских австроазиатов: роль ландшафтных барьеров и примеси по признаку пола» . Молекулярная биология и эволюция . 28 (2): 1013–1024. дои : 10.1093/molbev/msq288 . ПМЦ 3355372 . ПМИД 20978040 .
- ^ Рай, Нирадж; и др. (2012). «Филогеография гаплогруппы Y-хромосомы H1a1a-M82 показывает вероятное индийское происхождение европейского цыганского населения» . ПЛОС ОДИН . 7 (11): е48477. Бибкод : 2012PLoSO...748477R . дои : 10.1371/journal.pone.0048477 . ПМК 3509117 . ПМИД 23209554 .
- ^ Перейти обратно: а б с д ван Дрим, Джордж (2012). «Этима, тесла с плечами и молекулярные варианты». Андреа Эндер, Адриан Лиманн и Бернхард Вельхли (ред.). Методы в современной лингвистике . Берлин: Мутон де Грюйтер.
- ^ Вэнь, Бо; и др. (2004). «Генетические данные подтверждают демическое распространение ханьской культуры». Природа . 431 (7006): 302–305. Бибкод : 2004Natur.431..302W . дои : 10.1038/nature02878 . ПМИД 15372031 . S2CID 4301581 .
- ^ Вуд, Элизабет Т; и др. (2005). «Контрастные закономерности вариаций Y-хромосомы и мтДНК в Африке: свидетельства демографических процессов с учетом пола» . Европейский журнал генетики человека . 13 (7): 867–876. дои : 10.1038/sj.ejhg.5201408 . ПМИД 15856073 .
- ^ де Филиппо, Чезаре; и др. (2011). «Y-хромосомная вариация в Африке к югу от Сахары: взгляд на историю нигер-конголезских групп» . Молекулярная биология и эволюция . 28 (3): 1255–1269. дои : 10.1093/molbev/msq312 . ПМЦ 3561512 . ПМИД 21109585 .
- ^ Гебремескель, Эйоав I; Ибрагим, Мунтасер Э (декабрь 2014 г.). «Гаплогруппы Y-хромосомы E: их распространение и значение для происхождения афро-азиатских языков и скотоводства» . Европейский журнал генетики человека . 22 (12): 1387–1392. дои : 10.1038/ejhg.2014.41 . ISSN 1476-5438 . ПМК 4231410 . ПМИД 24667790 .
- ^ Шайвиц, Б.А.; и др. (1995). «Половые различия в функциональной организации мозга для языка». Природа . 373 (6515): 607–609. Бибкод : 1995Natur.373..607S . дои : 10.1038/373607a0 . ПМИД 7854416 . S2CID 4315684 .
- ^ Пью, КР; и др. (1996). «Мозговая организация составных процессов при чтении» . Мозг . 119 (4): 1221–1238. дои : 10.1093/мозг/119.4.1221 . ПМИД 8813285 .
- ^ Пью, КР; и др. (1997). «Прогнозирование эффективности чтения на основе профилей нейровизуализации: церебральная основа фонологических эффектов при идентификации печатного слова». Журнал экспериментальной психологии: человеческое восприятие и деятельность . 23 (2): 299–318. дои : 10.1037/0096-1523.23.2.299 . ПМИД 9103996 .
- ^ Йегер; и др. (1998). «Половые различия в областях мозга, активируемые грамматическими задачами и задачами по чтению». НейроОтчет . 9 (12): 2803–2807. дои : 10.1097/00001756-199808240-00022 . ПМИД 9760124 . S2CID 41971376 .
- ^ Кансаку, К; и др. (2000). «Половые различия в латерализации, выявленные в задних областях языка» . Кора головного мозга . 10 (9): 866–872. дои : 10.1093/cercor/10.9.866 . ПМИД 10982747 .
- ^ Росселл, С.Л.; и др. (2002). «Половые различия в функциональной активации мозга во время задачи на лексическое зрительное поле». Мозг и язык . 80 (1): 97–105. дои : 10.1006/brln.2000.2449 . ПМИД 11817892 . S2CID 34669770 .
- ^ Бакстер, LC; и др. (2003). «Половые различия в обработке семантического языка: функциональное МРТ-исследование». Мозг и язык . 84 (2): 264–272. дои : 10.1016/s0093-934x(02)00549-7 . ПМИД 12590915 . S2CID 12658733 .
- ^ Клементс, AM; и др. (2006). «Половые различия в церебральной латерализации языка и зрительно-пространственной обработки». Мозг и язык . 98 (2): 150–158. дои : 10.1016/j.bandl.2006.04.007 . ПМИД 16716389 . S2CID 32525196 .
- ^ Берман, Дуглас Д.; и др. (2008). «Половые различия в нейронной обработке речи у детей» . Нейропсихология . 46 (5): 1349–1362. doi : 10.1016/j.neuropsychologia.2007.12.021 . ПМЦ 2478638 . ПМИД 18262207 .
- ^ Перейти обратно: а б с ван Дрим, Джордж (2008). «Размышления об этнолингвистической предыстории Большого Гималайского региона». В Бриджит Хубер, Марианне Волкарт и Поле Видмере (ред.). Джомолунгма, Демавенд и Казбек: Фестиваль Роланда Бильмайера к его 65-летию (2 тома) . Галле: Международный институт тибетских и буддийских исследований. стр. 39–59.
- ^ Баккер, Питер Ян (1992). Наш собственный язык: происхождение мичифа, смешанного кри-французского языка канадских метисов . Амстердам: Университет Амстердама: докторская диссертация.
- ^ Баккер, Питер Ян (1994). «Мичиф, кри-французский смешанный язык охотников на буйволов-метисов в Канаде». В Питере Баккере и Маартене Моусе (ред.). Смешанные языки, 15 практических примеров переплетения языков (Исследования языка и использования языка, 13) . Амстердам: Институт фундаментальных исследований языка и его использования. стр. 13–33.
- ^ ван Дрим, Джордж (2001). Языки Гималаев . Лейден: Брилл.