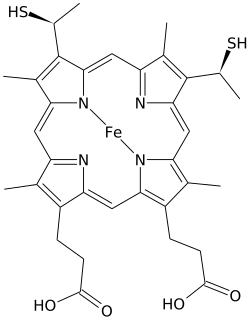
Гем С
 | |
| Идентификаторы | |
|---|---|
3D model ( JSmol ) | |
| ХимическийПаук | |
| МеШ | гем+С |
ПабХим CID | |
| Характеристики | |
| С 34 Н 36 О 4 Н 4 S 2 Fe | |
| Молярная масса | 684.64904 g/mol |
Если не указано иное, данные приведены для материалов в стандартном состоянии (при 25 °C [77 °F], 100 кПа). | |
Гем С (или гем С ) — важный вид гема .
История
[ редактировать ]Правильная структура гема С была опубликована в середине 20 века шведским биохимиком К.-Г. Пол. [1] Эта работа подтвердила структуру, впервые предложенную великим шведским биохимиком Хьюго Теореллом . Структура гема C на основе ЯМР и ИК-экспериментов восстановленной формы гема Fe(II) была подтверждена в 1975 году. [2] Структура гема С, включая абсолютную стереохимическую конфигурацию тиоэфирных связей, была впервые представлена для белка позвоночных - цитохрома с. [3] и теперь распространяется на многие другие белки, содержащие гем C.
Характеристики
[ редактировать ]Гем C отличается от гема B тем, что две винильные боковые цепи гема B заменены ковалентными тиоэфирными связями с апопротеином . Две тиоэфирные связи обычно образуются цистеиновыми остатками белка. Эти связи не позволяют гему C легко диссоциировать от голопротеина , цитохрома c , по сравнению с более легко диссоциируемым гемом B, который может диссоциировать от голопротеина, комплекса гем-белок, даже в мягких условиях. Это обеспечивает очень широкий диапазон структур и функций цитохрома c, при этом множество цитохромов типа c действуют в первую очередь как переносчики электронов. Окислительно-восстановительный потенциал цитохрома с также можно «тонко настроить» за счет небольших изменений в структуре белка и взаимодействии с растворителем. [4]
Количество единиц гема С, связанных с голопротеином , сильно варьируется. Для клеток позвоночных правилом является один гем C на белок, но для бактерий это число часто составляет 2, 4, 5, 6 или даже 16 групп гема C на голопротеин. Общепризнано, что количество и расположение групп гема C взаимосвязаны и даже необходимы для правильного функционирования голопротеина. Например, белки, содержащие несколько групп гема C, участвуют в реакциях множественного переноса электронов, особенно важным является восстановление шести электронов, необходимое для восстановления атмосферного азота до двух молекул органического аммиака. соотношение гема C и аминокислот обычно высокое Для бактериальных гемепротеинов , поэтому внутренняя часть некоторых белков цитохрома c кажется заполненной множеством групп гема C по сравнению с другими гемепротеинами. Некоторые гемопротеины, часто из одноклеточных организмов , могут содержать пять гемов C. [5] Комплекс bc 1 — еще один важный фермент , содержащий гем C-типа.
Тиоэфирные связи, по-видимому, предоставляют голопротеинам большую свободу функционирования. В целом, цитохромы типа c могут быть «тонко настроены» в более широком диапазоне окислительно-восстановительного потенциала, чем цитохромы b. Это может быть важной причиной того, что цитохром С практически повсеместно распространен на протяжении всей жизни. Гем С также играет важную роль в апоптозе, когда всего несколько молекул цитоплазматического цитохрома С, которые все еще должны содержать гем С, приводят к запрограммированной гибели клеток. [6] Цитохром С можно измерить в сыворотке человека и использовать в качестве маркера воспаления. [7]
В дополнение к этим экваториальным ковалентным связям железо гема также обычно аксиально координировано с боковыми цепями двух аминокислот , что делает железо гексакоординированным. Например, цитохром С млекопитающих и тунца содержит один гем C, который аксиально координирован с боковыми цепями как гистидина , так и метионина . [8] Возможно, из-за двух ковалентных связей, удерживающих гем с белком, железо гема С иногда аксиально лигируется с аминогруппой лизина или даже с водой.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Пол, КГ; Хёгфельдт, Эрик; Силлен, Ларс Гуннар; Кинелл, Пер-Улоф (1950). «Расщепление солями серебра цистеин-порфириновых связей в цитохроме с» . Acta Chemica Scandinavica . 4 : 239–244. doi : 10.3891/acta.chem.scand.04-0239 .
- ^ Коги, штат Вашингтон; Смайт, Джорджия; О'Киф, DH; Маскаски, Дж. Э.; Смит, М.Л. (1975). «Гем А цитохром с-оксидазы» . Журнал биологической химии . 250 (19): 7602–7622. дои : 10.1016/S0021-9258(19)40860-0 . ПМИД 170266 .
- ^ Такано Т.; Трус БЛ; Мандель Н.; Мандель Г.; Каллай О.Б.; Суонсон Р.; Дикерсон Р.Э. (1977). «Цитохром c тунца с разрешением 2,0 А. II. Анализ структуры ферроцитохрома» . Журнал биологической химии . 252 (2): 776–785. дои : 10.1016/S0021-9258(17)32784-9 . ПМИД 188826 .
- ^ Бергюс, AM; Брайер, Г.Д. (1992). «Конформационные изменения цитохрома с, зависящие от степени окисления». Дж. Мол. Биол . 223 (4): 959–976. дои : 10.1016/0022-2836(92)90255-i . ПМИД 1311391 .
- ^ Гвайер Джеймс Д., Ричардсон Дэвид Дж., Батт Джулиа Н. (2005). «Характеристики диода или туннельного диода? Разрешение каталитических последствий протон-связанного переноса электрона в многоцентровой оксидоредуктазе». Журнал Американского химического общества . 127 (43): 14964–14965. дои : 10.1021/ja054160s . ПМИД 16248601 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Боуман, SEJ, Брен, КЛ (2008). «Химия и биохимия гема С: функциональные основы ковалентного присоединения» . Нат. Прод. Представитель . 25 (6): 1118–1130. дои : 10.1039/b717196j . ПМЦ 2654777 . ПМИД 19030605 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Элефтериадис, Т.; Писсас, Г.; Лиакопулос, В.; Стафанидис, И. (2016). «Цитохром С как потенциально клинически полезный маркер митохондриального и клеточного повреждения» . Передний. Иммунол . 7 : 279. дои : 10.3389/fimmu.2016.00279 . ПМЦ 4951490 . ПМИД 27489552 .
- ^ Йе, С.Р., Хан, С., и Руссо, Д.Л. (1998). «Цитохром С сворачивается и разворачивается». Отчеты о химических исследованиях . 31 (11): 727–735. дои : 10.1021/ar970084p .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка )