субтеломера
Эта статья нуждается в дополнительных цитатах для проверки . ( апрель 2016 г. ) |
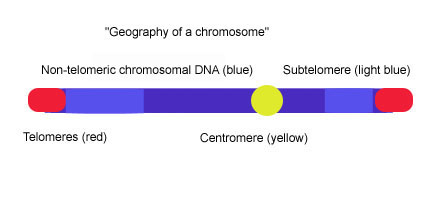
Субтеломеры — это сегменты ДНК между теломерными кэпами и хроматином.
Структура
[ редактировать ]Теломеры — это специализированные белки — конструкции ДНК , присутствующие на концах эукариотических хромосом, которые предотвращают их деградацию и сквозное слияние хромосом . большинства позвоночных состоит из длинных ( TTAGGG Теломерная ДНК n ) т.п.н. 3-20 повторов переменной длины, часто около Субтеломеры — это сегменты ДНК между теломерными шапками и хроматином . У позвоночных каждая хромосома имеет две субтеломеры, непосредственно прилегающие к длинным повторам (TTAGGG)n. Субтеломеры считаются наиболее дистальной (наиболее удаленной от центромеры ) областью уникальной ДНК хромосомы и представляют собой необычайно динамичную и изменчивую мозаику мультихромосомных блоков последовательности. Субтеломеры таких разнообразных видов, как человек, Plasmodium falciparum , Drosophila melanogaster и Saccharomyces cerevisiae структурно схожи, поскольку состоят из различных повторяющихся элементов, но размер субтеломеров и последовательность элементов сильно различаются у разных организмов. [ 1 ] У дрожжей ( S. cerevisiae ) субтеломеры состоят из двух доменов: проксимального и дистального (теломерного) доменов. Эти два домена различаются по содержанию последовательностей и степени гомологии с концами других хромосом, и они часто разделены участком вырожденных теломерных повторов (TTAGGG) и элементом, называемым «ядро X», который обнаруживается на всех концах хромосомы и содержит автономно реплицирующаяся последовательность (ARS) и сайт связывания ABF1. [ 2 ] [ 3 ] Проксимальный домен состоит из вариабельных межхромосомных дупликаций (<1–30 т.п.н. ); эта область может содержать такие гены, как Pho , Mel и Mal . [ 4 ] Дистальный домен состоит из 0–4 тандемных копий высококонсервативного элемента Y'; количество и хромосомное распределение элементов Y 'различно среди штаммов дрожжей. [ 5 ] Между ядром X и элементом Y' или ядром X и последовательностью TTAGGG часто существует набор из 4 элементов субтеломерных повторов (STR): STR-A, STR-B, STR-C и STR-D, который состоит из множества копий. теломерного мотива TTAGGG позвоночных. [ 6 ] Эта двухдоменная структура удивительно похожа на субтеломерную структуру хромосом 20p, 4q и 18p человека, в которой проксимальные и дистальные субтеломерные домены разделены участком вырожденных повторов TTAGGG, но картина, возникающая в результате изучения субтеломер других человеческих хромосом, хромосом указывает на то, что двухдоменная модель не применима повсеместно. [ 1 ]
Характеристики
[ редактировать ]Эта структура с повторяющимися последовательностями отвечает за частые события дупликации, которые создают новые гены, и события рекомбинации, лежащие в основе разнообразия комбинаций. Эти свойства создают разнообразие в индивидуальном масштабе и, следовательно, способствуют адаптации организмов к окружающей среде. Например, у Plasmodium falciparum во время интерфазы эритроцитарной стадии хромосомные окончания собираются на периферии ядра клетки, где они подвергаются частому делеции теломер и эффекту положения (TPE). Это событие, в дополнение к расширению и удалению субтеломерных повторов, приводит к полиморфизму размера хромосом и, таким образом, субтеломеры подвергаются эпигенетическому и генетическому контролю. Благодаря свойствам субтеломер Plasmodium falciparum уклоняется от иммунитета хозяина, изменяя антигенный и адгезивный характер инфицированных эритроцитов (см. Субтеломерные транскрипты). [ 7 ] [ 8 ]
Вариации
[ редактировать ]Вариации субтеломерных областей в основном представляют собой вариации STR из-за рекомбинации крупномасштабных участков, ограниченных (TTAGGG)n-подобными повторяющимися последовательностями, которые играют важную роль в рекомбинации и транскрипции. гаплотипы Таким образом, между людьми наблюдаются (варианты последовательности ДНК) и различия в длине.
Субтеломерные транскрипты
[ редактировать ]Субтеломерные транскрипты в основном состоят либо из псевдогенов (транскрибируемых генов, продуцирующих последовательности РНК, не транслируемые в белок), либо из семейств генов . У людей они кодируют обонятельные рецепторы , тяжелые цепи иммуноглобулинов и белки цинковых пальцев . У других видов некоторые паразиты, такие как Plasmodium и Trypanosoma brucei, развили сложные механизмы уклонения, позволяющие адаптироваться к враждебной среде, создаваемой хозяином, например, подвергая воздействию переменных поверхностных антигенов , чтобы избежать иммунной системы. Гены, кодирующие поверхностные антигены у этих организмов, расположены в субтеломерных областях, и было высказано предположение, что это предпочтительное расположение облегчает переключение и экспрессию генов, а также генерацию новых вариантов. [ 9 ] [ 10 ] Например, гены семейства var у Plasmodium falciparum (возбудителя малярии) преимущественно локализованы в субтеломерных областях. Антигенные вариации управляются эпигенетическими факторами, включая моноаллельную транскрипцию var в отдельных пространственных доменах на ядерной периферии ( ядерная пора ), дифференциальные метки гистонов на идентичных в остальном генах var и подавление var, опосредованное теломерным гетерохроматином . Другие факторы, такие как некодирующая РНК, продуцируемая в субтеломерных областях, прилегающих к генам var или внутри них , также могут способствовать антигенной изменчивости . [ 11 ] [ 12 ] У Trypanosoma brucei (возбудителя сонной болезни) вариация антигена вариабельного поверхностного гликопротеина (VSG) является важным механизмом, используемым паразитом для уклонения от иммунной системы хозяина. Экспрессия VSG является исключительно субтеломерной и происходит либо за счет активации молчащего гена VSG in situ, либо за счет перестройки ДНК, которая вставляет внутреннюю молчащую копию гена VSG в активный сайт теломерной экспрессии. В отличие от Plasmodium falciparum , у Trypanosoma brucei антигенные вариации регулируются эпигенетическими и генетическими факторами. [ 13 ] [ 14 ]
У Pneumocystis jirovecii семейство генов главного поверхностного гликопротеина (MSG) вызывает антигенные вариации. только ген MSG в уникальном локусе UCS (консервативная последовательность выше) Гены MSG представляют собой подобие коробочек на концах хромосом, и транскрибируется . Различные гены MSG могут занимать сайт экспрессии (UCS), что позволяет предположить, что рекомбинация может взять ген из пула молчащих доноров и установить его в сайт экспрессии, возможно, посредством кроссинговеров , активируя транскрипцию нового гена MSG и изменяя поверхностный антиген. Pneumocystis jirovecii . Переключению места экспрессии, вероятно, способствует субтеломерное расположение экспрессируемых и молчащих генов MSG. Второе семейство субтеломерных генов, MSR, не регулируется строго на уровне транскрипции, но может способствовать фенотипическому разнообразию. В антигенной изменчивости P. jirovecii доминирует генетическая регуляция. [ 15 ] [ 16 ]
Патологическое значение
[ редактировать ]Потеря теломерной ДНК в результате повторяющихся циклов деления клеток связана со старением или старением соматических клеток. Напротив, зародышевая линия и раковые клетки обладают ферментом теломеразой , который предотвращает деградацию теломер и поддерживает целостность теломер, что делает эти типы клеток очень долгоживущими.
У человека роль субтеломерных нарушений показана при лице-лопаточно-плечевой мышечной дистрофии (ЛЛПД), болезни Альцгеймера , эпилепсии. [ 17 ] и своеобразные синдромальные заболевания ( пороки развития и умственная отсталость). Например, ЛЛПД связан с делецией субтеломерной области хромосомы 4q. Ряд повторов от 10 до >100 т.п.н. расположен в нормальном субтеломере 4q, но у пациентов с ЛЛПД имеется только 1–10 повторов. Считается, что эта делеция вызывает заболевание из-за эффекта положения, который влияет на транскрипцию близлежащих генов, а не из-за потери самого массива повторов. [ 1 ]
Преимущества и эффекты
[ редактировать ]Субтеломеры гомологичны другим субтеломерам, расположенным в разных хромосомах, и представляют собой тип мобильных элементов — сегментов ДНК, способных перемещаться по геному. Хотя субтеломеры являются псевдогенами и не кодируют белки, они обеспечивают эволюционное преимущество за счет диверсификации генов. Дупликация, рекомбинация и делеция субтеломеров позволяют создавать новые гены и новые хромосомные свойства. [ 1 ] Преимущества субтеломеров были изучены на различных видах, таких как Plasmodium falciparum , [ 1 ] Дрозофила меланогастер , [ 1 ] и Saccharomyces cerevisiae , [ 1 ] поскольку они имеют генетические элементы, аналогичные человеческим, без учета длины и последовательности. [ 1 ] Субтеломеры могут играть ту же роль в растениях, поскольку такое же преимущество было обнаружено у обыкновенной фасоли, известной как Phaseolus vulgaris . [ 18 ]
Различные разновидности субтеломеров часто перестраиваются во время мейотической и митотической рекомбинации, что указывает на то, что субтеломеры часто перетасовываются, что вызывает новые и быстрые генетические изменения в хромосомах. [ 1 ] У Saccharomyces cerevisiae область 15 КБ хромосомы 7L в субтеломерах сохраняла жизнеспособность клеток при удалении теломеразы, тогда как удаление последних 15 КБ увеличивало старение хромосомы . [ 19 ] Нокаут субтеломеров в клетках делящихся дрожжей Schizosaccharomyces pombe не препятствует возникновению митоза и мейоза, что указывает на то, что субтеломеры не необходимы для клеточного деления. [ 20 ] Они пока не нужны для процесса митоза и мейоза, субтеломеры используют рекомбинацию клеточной ДНК. Нокаут субтеломер в клетках Schizosaccharomyces pombe не влияет на регуляцию множественных стрессовых реакций при лечении высокими дозами гидроксимочевины , камптотецина , ультрафиолетового излучения и тиабендазола . [ 20 ] Нокаут субтеломеров в клетках Schizosaccharomyces pombe не влиял на длину теломер, что указывает на то, что они не играют никакой роли в регуляции длины. [ 20 ] Однако субтеломеры сильно влияют на время репликации теломер. [ 21 ] Нокаут субтеломеров в клетках Schizosaccharomyces pombe после потери теломеразы не влияет на выживаемость клеток, что указывает на то, что субтеломеры не необходимы для выживания клеток. [ 20 ] Объяснение того, почему субтеломеры не нужны после потери теломеразы, заключается в том, что хромосомы могут использовать внутри- или межхромосомную циркуляризацию. [ 22 ] или МАТЬ [ 23 ] для поддержания хромосомной стабилизации. Однако использование межхромосомной циркуляризации вызывает нестабильность хромосом за счет создания двух центромер в одной хромосоме, что приводит к разрыву хромосом во время митоза. В ответ на это хромосома может вызвать инактивацию центромер, чтобы препятствовать образованию двух центромер, но это будет индуцировать образование гетерохроматина в центромерах. Гетерохроматин может быть вредным, если он попадает в место, где он не должен находиться. Субтеломеры несут ответственность за блокирование попадания гетерохроматина в область эухроматина . Субтеломеры могут смягчать эффекты инвазии гетерохроматина, распределяя гетерохроматин вокруг концов субтеломеров. Без субтеломеров гетерохроматин распространился бы по области субтеломеров, подбираясь слишком близко к важным генам. На этом расстоянии гетерохроматин может заставить замолчать гены, находящиеся поблизости, что приводит к более высокой чувствительности к осмотическому стрессу . [ 20 ]
Субтеломеры выполняют важные функции с белком Шугошина . Шугошин — центромерный белок, обеспечивающий сегрегацию хромосом во время мейоза и митоза. Существует два типа белка Шугошина: SGOL1 и SGOL2 . Sgo1 экспрессируется только в мейозе 1 для центромерного сцепления сестринских хромосом. [ 24 ] в то время как Sgo2, экспрессирующийся при мейозе и митозе, отвечает за сегрегацию хромосом на центромерах в М-фазе. У делящихся дрожжей Sgo2 локализован не только в центромерах, но и в субтеломерах. Sgo2 взаимодействует с субтеломерами во время интерфазы; середине фазы G2 и играет главную роль в формировании «шишка», представляющего собой высококонденсированное тело хроматина. Sgo2 остается в субтеломерах, в клетках которых отсутствует теломерная ДНК. Sgo2 репрессирует экспрессию субтеломерных генов, которые находятся в пути, отличном от H3K9me3 -Swi6-опосредованного гетерохроматина. Sgo2 также оказывает репрессивное воздействие на время репликации субтеломеров путем подавления Sld3, [ 25 ] фактор репликации в начале репликации. [ 26 ] Т.о., Sgo2 регулирует экспрессию и репликацию генов, обеспечивая правильную экспрессию субтеломерных генов и время репликации.
Анализ
[ редактировать ]Анализ субтеломеров, особенно секвенирование и профилирование субтеломеров пациентов, затруднен из-за повторяющихся последовательностей, длины участков и отсутствия баз данных по этой теме.
 [ оригинальное исследование? ]
[ оригинальное исследование? ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я Меффорд, Хизер С.; Траск, Барбара Дж. (февраль 2002 г.). «Сложная структура и динамическая эволюция субтеломеров человека». Обзоры природы Генетика . 3 (2): 91–102. дои : 10.1038/nrg727 . ПМИД 11836503 . S2CID 18918401 .
- ^ Луи, Э.Дж.; Наумова, Е.С.; Ли, А.; Наумов Г.; Хабер, Дж. Э. (март 1994 г.). «Конец хромосомы у дрожжей: его мозаичная природа и влияние на рекомбинационную динамику» . Генетика . 136 (3): 789–802. дои : 10.1093/генетика/136.3.789 . ПМК 1205885 . ПМИД 8005434 .
- ^ Уолмсли, Ричард В.; Чан, Кларенс С.М.; Тай, Бик-Квун; Питс, Томас Д. (июль 1984 г.). «Необычные последовательности ДНК, связанные с концами хромосом дрожжей». Природа . 310 (5973): 157–160. Бибкод : 1984Natur.310..157W . дои : 10.1038/310157a0 . ПМИД 6377091 . S2CID 4330408 .
- ^ Куассак, Эрик; Майлье, Эвелин; Робино, Сильвиана; Неттер, Пьер (декабрь 1996 г.). «Последовательность фрагмента ДНК длиной 39 411 п.о., покрывающего левый конец хромосомы VII Saccharomyces cerevisiae». Дрожжи . 12 (15): 1555–1562. doi : 10.1002/(SICI)1097-0061(199612)12:15<1555::AID-YEA43>3.0.CO;2-Q . ПМИД 8972578 . S2CID 44553592 .
- ^ Луи, Э.Дж.; Хабер, JE (июль 1992 г.). «Структура и эволюция субтеломерных Y'-повторов у Saccharomyces cerevisiae» . Генетика . 131 (3): 559–574. дои : 10.1093/генетика/131.3.559 . ПМК 1205030 . ПМИД 1628806 .
- ^ Луи, Эдвард Дж. (декабрь 1995 г.). «Концы хромосом Saccharomyces cerevisiae». Дрожжи . 11 (16): 1553–1573. дои : 10.1002/да.320111604 . ПМИД 8720065 . S2CID 36232717 .
- ^ Рубио, JP; Томпсон, Дж. К.; Коуман, AF (1 августа 1996 г.). «Гены var Plasmodium falciparum расположены в субтеломерной области большинства хромосом» . Журнал ЭМБО . 15 (15): 4069–4077. дои : 10.1002/j.1460-2075.1996.tb00780.x . ПМК 452127 . ПМИД 8670911 .
- ^ Су, Синь-чжуань; Хитвол, Вирджиния М.; Вертхаймер, Сэмюэл П.; Гине, Франсуаза; Херрфельдт, Жаклин А.; Петерсон, Дэвид С.; Рэвеч, Джеффри А.; Веллемс, Томас Э. (июль 1995 г.). «Большое разнообразное семейство генов var кодирует белки, участвующие в цитоадгезии и антигенных вариациях эритроцитов, инфицированных Plasmodium falciparum» . Клетка . 82 (1): 89–100. дои : 10.1016/0092-8674(95)90055-1 . ПМИД 7606788 .
- ^ Кано, Мария Изабель Н. (сентябрь 2001 г.). «Теломерная биология трипаносоматид: больше вопросов, чем ответов». Тенденции в паразитологии . 17 (9): 425–429. дои : 10.1016/S1471-4922(01)02014-1 . ПМИД 11530354 .
- ^ Барри, доктор юридических наук; Джинджер, ML; Бертон, П.; Маккалок, Р. (январь 2003 г.). «Почему гены непредвиденных обстоятельств паразитов часто связаны с теломерами?». Международный журнал паразитологии . 33 (1): 29–45. дои : 10.1016/S0020-7519(02)00247-3 . ПМИД 12547344 .
- ^ Шерф, Артур; Лопес-Рубио, Хосе Хуан; Ривьер, Лоик (октябрь 2008 г.). «Антигенная вариация Plasmodium falciparum». Ежегодный обзор микробиологии . 62 (1): 445–470. дои : 10.1146/annurev.micro.61.080706.093134 . ПМИД 18785843 .
- ^ Гизетти, Жюльен; Шерф, Артур (май 2013 г.). «Замолчать, активировать, уравновесить и переключиться! Механизмы антигенных вариаций» . Клеточная микробиология . 15 (5): 718–726. дои : 10.1111/cmi.12115 . ПМЦ 3654561 . ПМИД 23351305 .
- ^ Кросс, Джордж AM (апрель 1996 г.). «Антигенные вариации трипансосом: секреты раскрываются медленно». Биоэссе . 18 (4): 283–291. дои : 10.1002/bies.950180406 . ПМИД 8967896 . S2CID 37442327 .
- ^ Руденко, Г. (1 октября 2000 г.). «Полиморфные теломеры африканской трипаносомы Trypanosoma brucei» . Труды Биохимического общества . 28 (5): 536–540. дои : 10.1042/bst0280536 . ПМЦ 3375589 . ПМИД 11044370 .
- ^ Стрингер, Джеймс Р. (2014). «Субтеломеры Pneumocystis carinii». Субтеломеры . стр. 101–115. дои : 10.1007/978-3-642-41566-1_5 . ISBN 978-3-642-41565-4 .
- ^ Портной, Д.А.; Стрингер, Джеймс Р.; Кили, Скотт П. (1 февраля 2001 г.). «Генетика экспрессии поверхностного антигена у Pneumocystis carinii» . Инфекция и иммунитет . 69 (2): 627–639. дои : 10.1128/IAI.69.2.627-639.2001 . ПМК 97933 . ПМИД 11159949 .
- ^ Меффорд, Хизер С.; Кук, Джозеф; Госп, Сидни М. (19 ноября 2012 г.). «Эпилепсия, вызванная делецией субтеломер 20q13.33, маскирующаяся под пиридоксинзависимую эпилепсию» . Американский журнал медицинской генетики, часть A. 158А (12): 3190–3195. дои : 10.1002/ajmg.a.35633 . ПМИД 23166088 . S2CID 19998295 — через JSTOR.
- ^ Чен, Николас В.Г.; Таро, Винсент; Рибейру, Тьяго; Магделенат, Гислен; Эшфилд, Том; Иннес, Роджер В.; Педроса-Харанд, Андреа; Жеффруа, Валери (14 августа 2018 г.). «Субтеломеры обыкновенных бобов являются горячими точками рекомбинации и эволюции генов устойчивости к благосклонности» . Границы в науке о растениях . 9 : 1185. doi : 10.3389/fpls.2018.01185 . ПМК 6102362 . ПМИД 30154814 .
- ^ Жоливе, Паскаль; Серхал, Камар; Граф, Фреймворк; Эберхард, Стивен; Сюй, Чжоу; Люк, Брайан; Тейшейра, Мария Тереза (12 февраля 2019 г.). «Субтеломерная область влияет на теломераза-отрицательное репликативное старение у Saccharomyces cerevisiae» . Научные отчеты 9 (1): 1845. Бибкод : 2019NatSR... 9.1845J. дои : 10.1038/ s41598-018-38000-9 ПМК 6372760 . ПМИД 30755624 .
- ^ Jump up to: а б с д и Таширо, Санки; Нишихара, Юки; Куго, Кадзуто; Охта, Кунихиро; Кано, Джунко (13 октября 2017 г.). «Субтеломеры обеспечивают защиту экспрессии генов и гомеостаза хромосом» . Исследования нуклеиновых кислот . 45 (18): 10333–10349. дои : 10.1093/nar/gkx780 . ПМЦ 5737222 . ПМИД 28981863 .
- ^ Пикере-Стефан, Лор; Рикул, Мишель; Хемпель, Уильям М.; Сабатье, Лор (2 сентября 2016 г.). «Время репликации теломер человека сохраняется во время иммортализации и зависит от соответствующих субтеломеров» . Научные отчеты . 6 (1): 32510. Бибкод : 2016НатСР...632510П . дои : 10.1038/srep32510 . ПМК 5009427 . ПМИД 27587191 .
- ^ Ван, Сяорун; Бауманн, Питер (22 августа 2008 г.). «Слияние хромосом после потери теломер опосредовано одноцепочечным отжигом» . Молекулярная клетка . 31 (4): 463–473. doi : 10.1016/j.molcel.2008.05.028 . ПМИД 18722173 .
- ^ Джайн, Деванши; Хебден, Анна К.; Накамура, Тору М.; Миллер, Кайл М.; Купер, Джулия Промизель (сентябрь 2010 г.). «Выжившие HAATI заменяют канонические теломеры блоками общего гетерохроматина». Природа . 467 (7312): 223–227. Бибкод : 2010Natur.467..223J . дои : 10.1038/nature09374 . ПМИД 20829796 . S2CID 205222290 .
- ^ Ватанабэ, Ёсинори (июль 2005 г.). «Слипание сестринских хроматид вдоль плеч и центромер». Тенденции в генетике . 21 (7): 405–412. дои : 10.1016/j.tig.2005.05.009 . ПМИД 15946764 .
- ^ Брук, Ирина; Каплан, Дэниел Л. (6 ноября 2015 г.). «Белок инициации репликации Sld3/Треслин управляет сборкой геликазы репликационной вилки во время S-фазы» . Журнал биологической химии . 290 (45): 27414–27424. дои : 10.1074/jbc.M115.688424 . ПМЦ 4646389 . ПМИД 26405041 . (Отозвано, см. дои : 10.1074/jbc.A115.688424 , PMID 28623197 , Часы втягивания )
- ^ Таширо, Тэцуя, Ацуси; Бан, Тору, Кадзуми; Куго, Кадзуто; Хираока, Ясуката, Хисао; 2016). «Шугошин формирует специализированный хроматиновый домен в субтеломерах, который регулирует время транскрипции и репликации» . 7 ( 1 ): 10393. Bibcode : ...710393T doi : 10.1038 /ncomms10393 . 4021 Nature . 2016NatCo . Communications
Внешние ссылки
[ редактировать ]- Поток генетической информации. Архивировано 10 июля 2010 г. в Wayback Machine — файл PDF. См. Таблицу 5.5.