Омомииды
| Омомииды | |
|---|---|
| |
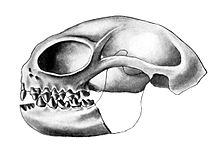
| Череп Анаптоморфа | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Приматы |
| Подотряд: | Хаплорини |
| Клэйд : | † Омомиообразные |
| Суперсемейство: | † Омомиоидея |
| Семья: | † Омомииды |
| Подгруппы | |
| |
| Синонимы | |
Omomyidae — это группа ранних приматов , которые распространились в эпоху эоцена примерно от 55 до 34 миллионов лет назад (млн лет назад). Ископаемые омомииды встречаются в Северной Америке, Европе и Азии, что делает их одной из двух групп приматов эоцена с географическим распространением, охватывающим голарктические континенты, вторая группа — адапиды (семейство Adapidae ). Ранние представители Omomyidae и Adapidae появляются внезапно в начале эоцена (56 млн лет назад) в Северной Америке, Европе и Азии и являются самыми ранними известными кроновыми приматами.
Характеристики
[ редактировать ]
Особенности, которые характеризуют многих омомид, включают большие орбиты (глазницы), укороченные ростры и зубные ряды, потерю передних премоляров, щечные зубы, приспособленные к насекомоядной или плодоядной диете, и относительно небольшую массу тела (т. е. менее 500 г). Однако к концу среднего эоцена (около 40 млн лет назад) некоторые североамериканские омомииды развили массу тела, превышающую 1 кг (2,2 фунта), и стали питаться плодоядными или листовидными растениями . Самыми крупными омомидами были Macrotarsius и Ourayia , оба весили 1,5–2 кг (3,3–4,4 фунта). [ 3 ] Большие орбиты у таких родов, как Tetonius , Shoshonius , Necrolemur и Microchoerus , указывают на то, что эти таксоны, вероятно, вели ночной образ жизни . По крайней мере, один род омомид из позднего эоцена Техаса ( Rooneyia ) имел маленькие орбиты и, вероятно, вел дневной образ жизни .
Как и ныне живущие приматы , омомиды имели хватательные руки и ноги с ногтями на концах вместо когтей, хотя у них были туалетные когти, как у современных лемуров. [ 4 ] Особенности их скелетов убедительно указывают на то, что омомииды жили на деревьях. [ 3 ] По крайней мере, у одного рода ( Necrolemur ) кости голени, большеберцовая и малоберцовая кости были слиты, как у современных долгопятов . Эта особенность может указывать на то, что Некролемур часто прыгал. Большинству других родов омомид (например, Omomys ) не хватает специализации для прыжков, а их скелеты больше похожи на скелеты ныне живущих карликовых и мышиных лемуров .
Систематика и эволюционные взаимоотношения омомид являются спорными. Авторы предположили, что омомииды бывают:
- стеблевые гаплорины [т.е. базальные члены группы, включая живых долгопятов и антропоидов]. [ 5 ]
- стебель tarsiiformes [т.е. базальные ответвления линии долгопятов]. [ 6 ]
- стволовые приматы более тесно связаны с адапидами, чем с современными таксонами приматов.
Недавние исследования показывают, что Omomyiformes являются стеблевыми гаплоринами, что делает их, вероятно, парафилетической группой. [ 7 ]
Попытки связать омомид с живыми группами осложнялись их примитивной ( плезиоморфной ) анатомией скелета. Например, у омомид отсутствуют многочисленные скелетные специализации, присущие современным гаплоринам. Эти адаптации гаплорина, отсутствующие у омомид, включают:
- значительная редукция канала стременной ветви внутренней сонной артерии .
- Маршрут канала для размещения мыса внутренней сонной артерии через слуховую буллу височной кости, т.е. «пербуллярный» (а не через мыс барабанной полости , «транспромонториальный»).
- контакт алисфеноидной и скуловой костей.
- наличие передней добавочной полости, сливающейся с барабанной полостью .
У омомид также имеется щель между верхними центральными резцами, что предположительно указывает на наличие ринария и желобка для направления жидкости в сошниково-носовой орган. Омомииды как группа также лишены большинства производных специализаций современных долгопятов, таких как чрезвычайно увеличенные глазницы ( Shoshonius возможное исключение - ), большое надматочное отверстие для анастомоза между задним ушным и средним менингеальным кровообращением (опять же, Shoshonius - это возможное исключение, но содержимое отверстия у этого вымершего таксона неизвестно) и крайние посткраниальные приспособления к прыжкам.
Среди приматов омомииды обладают уникальной производной характеристикой. Это наличие афанерной (невидимой или легко различимой из-за своего положения) или «интрабуллярной» (внутри булл) эктотимпанальной кости, соединенной с латеральной стенкой слухового буллы неразрывной кольцевой перемычкой.
Классификация
[ редактировать ]Этот раздел нуждается в дополнительных цитатах для проверки . ( декабрь 2023 г. ) |
- Семья Mommyidae
- Альтаниус
- Местность
- Подсемейство Анаптоморфины.
- Подсемейство Microchoerinae.
- Подсемейство Мама
- Бронтомомис [ 9 ]
- Диабломомис
- Эквиемакиус [ 9 ]
- Ганнелл долгопятник [ 9 ]
- Хуэрфаниус
- Митоний
- Палеакодон
- Племя Рунейини
- Племя Штейнин
- Племя Уинтанини
- Племя гемиакодонтини
- Племя Провидец
- Племя Макротарсиини
- Племя Вашакиини
- Племя в Юте
Ссылки
[ редактировать ]- ^ Сэвидж, Р.Дж.Г., и Лонг, М.Р. (1986). Эволюция млекопитающих: иллюстрированное руководство . Нью-Йорк: факты в архиве. п. 365. ИСБН 978-0-8160-1194-0 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Морс, Пол Э.; Честер, Стивен ГБ; Бойер, Дуг М.; Смит, Тьерри; Смит, Ричард; Гигасе, Пол; Блох, Джонатан И. (01 марта 2019 г.). «Новые окаменелости, систематика и биогеография старейшего известного коронного примата Тейярдина из раннего эоцена Азии, Европы и Северной Америки» . Журнал эволюции человека . 128 : 103–131. дои : 10.1016/j.jhevol.2018.08.005 . ISSN 0047-2484 . ПМИД 30497682 . S2CID 54167483 .
- ^ Jump up to: а б Рэйчел Х. Данн. « Дополнительные посткраниальные останки омомидных приматов из формации Уинта, штат Юта, и последствия для двигательного поведения крупнотелых омомид ». Журнал эволюции человека Том 58, выпуск 5, май 2010 г., стр. 406–417.
- ^ «Ранние приматы с когтями» .
- ^ Кей, Ричард Ф.; Росс, Каллум; Уильямс, Блайт А. (1997). «Антропоидное происхождение». Наука . 275 (5301): 797–804. дои : 10.1126/science.275.5301.797 . ПМИД 9012340 . S2CID 220087294 .
- ^ Салай, Фредерик (1976). «Систематика Omomyidae (Tarsiiformes, приматов): таксономия, филогения и адаптации». Бюллетень Американского музея естественной истории . 156 (3): 157–450.
- ^ Росси, Джеймс Б.; Смит, Тимоти Д.; Борода, К. Кристофер; Годино, Марк; Роу, Тимоти Б. (2018). «Носослёзная анатомия и происхождение гаплорина» . Журнал эволюции человека . 114 : 176–183. дои : 10.1016/j.jhevol.2017.11.004 . ISSN 0047-2484 . ПМИД 29447758 .
- ^ Jump up to: а б Перри, JMG; Дутчак, Арканзас; Теодор, Дж. М. (2023). «Новые приматы из эоцена Саскачевана, Канада: пересмотр приматов из формации Сайпресс-Хиллз с описанием новых таксонов» . Электронная палеонтология . 26 (2). 26.2.20. дои : 10.26879/1246 .
- ^ Jump up to: а б с Эми Л. Этуотер; Э. Кристофер Кирк (2018). «Новые омомии среднего эоцена (приматы, Haplorhini) из округа Сан-Диего, Калифорния». Журнал эволюции человека. в прессе. doi:10.1016/j.jhevol.2018.04.010.
Внешние ссылки
[ редактировать ]


| Базы данных органов управления : Национальные |
|---|