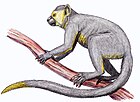
Долгопят
| Долгопяты [ 1 ] [ 2 ] Временной диапазон: От среднего эоцена до современности
| |
|---|---|

| |
| Филиппинский долгопят ( Carlito syrichta ) | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Приматы |
| Подотряд: | Хаплорини |
| Инфрапорядок: | Долгопятообразные |
| Семья: | Долгопяты Грей , 1825 г. |
| Типовой род | |
| Долгопяты Сторр , 1780 г.
| |
| Роды | |
Долгопяты ( / ˈ t ɑːr s i ər z / TAR -see-ərz ) — -гаплорины приматы семейства Tarsiidae , которое само по себе является единственным существующим семейством внутри инфраотряда Tarsiiformes . Хотя в доисторические времена эта группа была более распространена по всему миру, все виды, живущие сегодня, обитают только в морской Юго-Восточной Азии , преимущественно обитая в Брунее , Индонезии , Малайзии и на Филиппинах . [ 3 ]

Они встречаются в основном в лесных средах обитания, особенно в лесах с лианами , поскольку лоза обеспечивает долгопятам вертикальную поддержку при лазании по деревьям. [ 4 ]
Эволюционная история
[ редактировать ]Ископаемая запись
[ редактировать ]Окаменелости долгопятовидных приматов были обнаружены в Азии, Европе и Северной Америке (со спорными окаменелостями из Северной Африки), но дошедшие до нас долгопяты обитают только на нескольких островах Юго-Восточной Азии. Летопись окаменелостей показывает, что их зубной ряд не сильно изменился, за исключением размера, за последние 45 миллионов лет.
В семействе Tarsiidae имеются два вымерших рода — Xanthorhysis и Afrotarsius ; однако местонахождение Афротарсиуса не установлено, [ 5 ] и иногда его относят к отдельному семейству Afrotarsiidae внутри инфраотряда Tarsiiformes. [ 6 ] или считается человекообразным приматом. [ 7 ]
На данный момент из летописи окаменелостей известны четыре ископаемых вида долгопятов:
- Tarsius eocaenus известен из среднего эоцена в Китае . [ 5 ] [ 8 ]
- Hesperotarsius thailandicus жил в раннем миоцене на северо-западе Таиланда . [ 5 ] [ 9 ]
- Hesperotarsius sindhensis жил в миоцене на территории Пакистана . [ 10 ]
- Tarsius sirindhornae жил в среднем миоцене на севере Таиланда. [ 7 ]
Род Tarsius имеет более обширную летопись окаменелостей, чем любой другой род приматов, но отнесение окаменелостей эоцена и миоцена к этому роду сомнительно. [ 11 ]
Классификация
[ редактировать ]Филогенетическое стреппсириновыми положение современных долгопятов внутри отряда Приматы обсуждалось на протяжении большей части 20-го века, и долгопятов поочередно классифицировали со приматами в подотряде Prosimii или как сестринскую группу обезьян ( Anthropoidea) в инфраотряде Haplorhini . Утверждается, что анализ вставок SINE , типа макромутации ДНК, предоставляет очень убедительные доказательства монофилии Haplorhini , тогда как другие доказательства, такие как данные о последовательностях ДНК , остаются неоднозначными. Таким образом, некоторые систематики утверждают, что дебаты окончательно решены в пользу монофилетического Haplorrini. Как и у обезьян, у долгопятов есть мутация в гене L-гулонолактоноксидазы (GULO), которая не позволяет их организму синтезировать витамин С, поэтому им приходится находить его в рационе. Поскольку стрепсиррины не имеют этой мутации и сохранили способность вырабатывать витамин С, генетическая особенность, обуславливающая необходимость в нем в рационе, может привести к тому, что долгопяты будут помещены в гаплорины. [ 12 ]

На более низком филогенетическом уровне все долгопяты до недавнего времени были отнесены к роду Tarsius . [ 1 ] в то время как обсуждался вопрос о том, следует ли отнести этот вид к двум ( сулавеси и филиппинско -западной группе) или к трем отдельным родам (сулавеси, филиппинская и западная группы). [ 13 ] на уровне вида Таксономия сложна, при этом морфология часто имеет ограниченное применение по сравнению с вокализацией. [ нужна ссылка ] Дальнейшая путаница существовала по поводу действительности некоторых имен. широко используемый T. dianae Среди прочего, было показано, что является младшим синонимом T. dentatus , и, для сравнения, T. Spectrum теперь считается младшим синонимом T. tarsier . [ 1 ]
В 2010 году Колин Гровс и Майрон Шекеле предложили разделить род Tarsius на три рода: филиппинских долгопятов (род Carlito ), западных долгопятов (род Cephalopachus ) и восточных долгопятов (род Tarsius ). Это было основано на различиях в зубном ряду , размере глаз, длине конечностей и рук, пучках хвоста, подушечках хвоста, количестве молочных желез , количестве хромосом, социоэкологии , вокализации и распространении. Старший таксон вида, T. tarsier, был ограничен популяцией острова Селаяр , что затем потребовало воскрешения несуществующего таксона T. fuscus . [ 2 ]
В 2014 году ученые опубликовали результаты генетического исследования всего ареала филиппинского долгопята , выявив ранее непризнанное генетическое разнообразие. В установленной систематике признаны три подвида: Carlito syrichta syrichta из Лейте и Самара , C. syrichta fraterculus из Бохола и C. syrichta Carbonarius из Минданао . Их анализ последовательностей митохондриальной и ядерной ДНК показал, что ssp. syrichta и fraterculus могут представлять одну линию, тогда как ssp. Carbonarius может представлять две линии: одна занимает большую часть Минданао, а другая находится на северо-востоке Минданао и близлежащем острове Динагат , который авторы назвали «долгопятом Динагат-Карага». Для обзора систематики долгопятов на Филиппинах потребуются более детальные исследования, объединяющие морфологические данные. [ 14 ]
- Инфраотряд Tarsiiformes
- Семейство Tarsiidae : долгопяты.
- Род Карлито
- Филиппинский долгопят , Carlito syrichta
- К. с. Сирихта
- К. с. fraterculus (объединить в C. s. syrichta ?)
- К. с. карбонарий
- Филиппинский долгопят , Carlito syrichta
- Род Цефалопах.
- Долгопят Хорсфилда , Cephalopachus Bancanus
- К. б. банканус
- К. б. натуненсис
- К. б. Борей
- К. б. джемпер
- Долгопят Хорсфилда , Cephalopachus Bancanus
- Род Тарсиус
- Долгопят Дианы , T. dentatus
- Макассарский долгопят T. fuscus
- Ларианский долгопят , Т. ларианг
- Долгопят Нимица , Т. niemitzi
- Пеленг долгопят , T. pelengensis
- Сангихе долгопят , T. sangirensis
- Долгопят спектральный Гурского , T. Spectrumgurskyae [ 15 ]
- Долгопят Джатны , T. supriatnai [ 15 ]
- Спектральный долгопят , Т. долгопят
- Долгопят с острова Сиау , T. tumpara
- Карликовый долгопят , T. pumilus
- Долгопят Уоллеса , T. wallacei
- Род Карлито
- Семейство Tarsiidae : долгопяты.
Анатомия и физиология
[ редактировать ]
Долгопяты — маленькие животные с огромными глазами; Каждое глазное яблоко имеет диаметр примерно 16 миллиметров (0,63 дюйма) и по размеру равно всему мозгу, а в некоторых случаях больше его. [ 16 ] [ 17 ] Уникальная анатомия черепа долгопятов обусловлена необходимостью сбалансировать их большие глаза и тяжелую голову, чтобы они могли молча ждать питательную добычу. [ 18 ] У долгопятов сильное слуховое чутье, и их слуховая кора уникальна. [ 18 ] У долгопятов также длинные задние конечности, главным образом из-за удлиненных костей предплюсны , от которых животные и получили свое название. Сочетание удлиненных лапок и сросшихся голеней делает их морфологически приспособленными к вертикальному цеплянию и прыжкам. [ 19 ] Длина головы и тела составляет от 10 до 15 см, но задние конечности примерно в два раза длиннее (включая ступни), а также имеют тонкий хвост длиной от 20 до 25 см. Их пальцы также удлинены, причем третий палец примерно такой же длины, как плечо. На большинстве пальцев есть ногти, но на втором и третьем пальцах задних лап вместо этого есть когти, которые используются для ухода за шерстью. У долгопятов мягкий бархатистый мех, обычно желтовато-коричневого, бежевого или охристого цвета. [ 20 ]
Морфология долгопятов позволяет им поворачивать голову на 180 градусов в любом направлении, что позволяет им видеть вокруг себя на 360 градусов. [ 21 ] Их зубная формула также уникальна: 2.1.3.3 1.1.3.3 [ 22 ] В отличие от многих ночных позвоночных, долгопяты лишены светоотражающего слоя ( tapetum lucidum ) сетчатки и имеют ямку .
Мозг долгопята отличается от мозга других приматов расположением связей между двумя глазами и латеральным коленчатым ядром , которое является основным отделом таламуса , получающим зрительную информацию. Последовательность клеточных слоев, получающих информацию от ипсилатеральных (одна и та же сторона головы) и контралатеральных (противоположная сторона головы) глаз в латеральном коленчатом ядре, отличает долгопятов от лемуров , лори и обезьян , которые все похожи в этом отношении. [ 23 ] Некоторые нейробиологи предположили, что «эта кажущаяся разница отличает долгопятов от всех других приматов, подтверждая мнение о том, что они возникли на ранней независимой линии эволюции приматов». [ 24 ]
Филиппинские долгопяты способны слышать частоты до 91 кГц. Они также способны издавать звуки с преобладающей частотой 70 кГц. [ 25 ]
нет В отличие от большинства приматов, у самцов долгопятов бакулы . [ 26 ]
Поведение
[ редактировать ]Карликовые долгопяты отличаются от других видов морфологией, общением и поведением. [ 27 ] Различия в морфологии, которые отличают карликовых долгопятов от других видов, вероятно, основаны на их высокогорной среде обитания. [ 28 ]
Все виды долгопятов ведут ночной образ жизни, но, как и многие ночные организмы, некоторые особи могут проявлять большую или меньшую активность в дневное время. Судя по анатомии всех долгопятов, все они приспособлены к прыжкам, хотя все они различаются в зависимости от своего вида. [ 29 ] [ 30 ] [ 31 ] [ 32 ]
Экологическая изменчивость ответственна за различия в морфологии и поведении долгопятов, поскольку разные виды адаптируются к местным условиям в зависимости от уровня высоты. [ 33 ] Например, более холодный климат на возвышенностях может повлиять на морфологию черепа. [ 34 ]
Долгопяты, как правило, чрезвычайно пугливые животные и чувствительны к яркому свету, громким звукам и физическому контакту. Сообщается, что они ведут себя суицидально, когда подвергаются стрессу или содержатся в неволе. [ 35 ] [ 36 ]
Хищники
[ редактировать ]Из-за своего небольшого размера долгопяты становятся добычей других животных. Долгопяты в основном обитают в нижних слоях растительности, поскольку им угрожают как наземные хищники, такие как кошки, ящерицы и змеи, так и воздушные хищники, такие как совы и птицы. Проживая в этих нижних слоях, они могут свести к минимуму свои шансы стать жертвами, оставаясь над землей и при этом оставаясь достаточно низкими, чтобы избегать хищных птиц.
Долгопяты, хотя и известны как застенчивые и замкнутые люди, известны как хищники. В природе моббинг – это преследование хищников с целью снизить вероятность нападения. Когда хищники приближаются, долгопяты издают предупреждающий сигнал. На зов откликнутся другие долгопяты, и через короткий промежуток времени от 2 до 10 долгопятов появятся и набросятся на хищника. Большую часть группы составляют взрослые самцы, но иногда встречаются одна-две самки. В то время как группы долгопятов содержат только одного взрослого самца, самцы с других территорий присоединятся к толпе, а это означает, что на хищника нападают несколько альфа-самцов долгопятов. [ 37 ] [ 21 ] [ 38 ]
Диета
[ редактировать ]Долгопяты — единственные ныне полностью приматы плотоядные , хотя в основном насекомоядные , ловящие беспозвоночных, прыгая на них. Долгопяты также охотятся на различных древесных и мелких лесных животных, включая прямокрылых , жуков-скарабеев , маленьких летающих лягушек , ящериц и, изредка, крабов -амфибий , которые забираются на нижние части деревьев. [ 39 ] [ 40 ] [ 41 ] Однако установлено, что их любимой добычей являются членистоногие , жуки , паукообразные , тараканы , кузнечики , кузнечики , цикады и трости . [ 21 ] Также известно, что долгопяты редко охотятся на птенцов, маленьких древесных змей и даже детенышей летучих мышей. [ 20 ]
Воспроизведение
[ редактировать ]Беременность длится около шести месяцев, [ 42 ] а долгопяты рожают одного потомства. Молодые долгопяты рождаются покрытыми шерстью, с открытыми глазами и способны лазить уже в день рождения. Половой зрелости они достигают к концу второго года жизни. Социальный образ жизни и система спаривания различаются: долгопяты из Сулавеси живут небольшими семейными группами, в то время как филиппинские и западные долгопяты, как сообщается, спят и кормятся в одиночку.
Сохранение
[ редактировать ]Долгопяты никогда не образовывали в неволе успешные гнездовые колонии. Частично это может быть связано с их особыми потребностями в питании. [ 43 ] [ 44 ] [ 45 ] [ 46 ] [ 47 ]
В заповеднике недалеко от города Корелла на филиппинском острове Бохол добились определенных успехов в восстановлении популяций долгопятов. [ 48 ] Филиппинский фонд долгопятов (PTFI) разработал большой полудикий вольер, известный как Центр исследований и разработок долгопятов. Карлито Писаррас, также известный как «Человек-долгопят», основал этот заповедник, где посетители могут наблюдать за долгопятами в дикой природе. По состоянию на 2011 год святилище содержалось им и его братом. [ нужна ссылка ] Деревья в заповеднике населены ночными насекомыми, составляющими рацион долгопята. [ 49 ]
Природоохранный статус всех долгопятов уязвим к исчезновению. Долгопяты являются видом, зависящим от сохранения, а это означает, что им необходимо больше и лучше управлять охраняемыми местами обитания, иначе они обязательно вымрут в будущем. [ 18 ]
Первое количественное исследование моделей активности филиппинских долгопятов (Tarsius syrichta) в неволе было проведено в Центре охраны филиппинских долгопятов Субайон в Биларе, Бохол, Филиппины. С декабря 2014 г. по январь 2016 г. наблюдения за самками и самцами T. syrichta проводились в зависимости от их времени, отведенного на нормальную деятельность в периоды без спаривания и в период спаривания. В период отсутствия спаривания значительная часть времени их бодрствования тратилась на сканирование, а затем на отдых, поиск пищи и путешествия. Кормление, мечение запахами, уход за собой, социальная активность и другие виды деятельности были минимальными. Сканирование по-прежнему было обычным занятием парных полов во время брачного сезона. Тем не менее, время отдыха заметно сократилось, в то время как увеличение количества путешествий и поиска пищи было очевидным. Эти результаты рассматриваются как возможность продолжения успешного содержания T.syrichta в неволе из-за антропогенных угроз. [ 50 ]
Описанный в 2008 году долгопят с острова Сиау в Индонезии считается находящимся под угрозой исчезновения и в 2008 году был включен в число 25 приматов мира, находящихся под наибольшей угрозой исчезновения, по версии Conservation International и Группы специалистов по приматам МСОП/SCC. [ 51 ] Правительство Малайзии защищает долгопятов, внося их в список полностью охраняемых животных Саравака , малайзийского штата на Борнео, где они обычно встречаются. [ 52 ]
Гражданское правительство Тупи и благотворительная организация Endangered Species International (ESI) организуют новую программу по сохранению долгопятов горы Матутум возле Тупи в Южном Котабато на острове Минданао. Tarsier UK также участвует в работе, помогая правительству Тупи рассказать детям Тупи о важности этого животного. ESI надеется построить центр для посетителей на склонах горы Матутум и помочь местным коренным народам вести более экологичное сельское хозяйство и заботиться о долгопятах. Первым этапом в этом процессе является информирование местного населения о важности обеспечения безопасности животных. Несколько местных деревьев, благоприятных для долгопятов, были пересажены на землю, которая ранее была расчищена для посадки фруктовых деревьев и кокосовых пальм . [ нужна ссылка ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с Гроувс, CP (2005). Уилсон, Делавэр ; Ридер, Д.М. (ред.). Виды млекопитающих мира: таксономический и географический справочник (3-е изд.). Балтимор: Издательство Университета Джонса Хопкинса. стр. 127–128. ISBN 0-801-88221-4 . OCLC 62265494 .
- ^ Jump up to: а б Гроувс, К.; Шекеле, М. (2010). «Роды и виды долгопятов». Международный журнал приматологии . 31 (6): 1071–1082. дои : 10.1007/s10764-010-9443-1 . S2CID 21220811 .
- ^ Редакторы Британской энциклопедии. "Долгопят." Британская энциклопедия , Британская энциклопедия, Inc., 14 апреля 2019 г., http://www.britannica.com/animal/tarsier .
- ^ Симеон, SGF; Дуя, МРМ; Дуя, МВ; Галиндон, JMM; Пасион, БО; Онг, PS (2020). «Жизнь на небольших пространствах: характеристика лесных фрагментов и их использование филиппинскими долгопятами (Tarsius syrichta Linnaeus, 1758) на острове Минданао, Филиппины». Приматы . 61 (3): 529–542. дои : 10.1007/s10329-020-00798-2 . ПМИД 32043166 . S2CID 211075031 .
- ^ Jump up to: а б с Ганнелл, Г.; Роуз, К. (2002). «tarsiiformes: эволюционная история и адаптация». В Хартвиге, WC (ред.). Летопись окаменелостей приматов . Издательство Кембриджского университета. ISBN 978-0-521-66315-1 .
- ^ Маккенна, MC, и Белл, SK 1997. Классификация млекопитающих выше уровня вида. Издательство Колумбийского университета, Нью-Йорк, 337–340 стр. ISBN 0-231-11013-8
- ^ Jump up to: а б Чиамани Ю., Лебрен Р., Ями К. и Джагер Ж.-Дж. (2010). «Новый среднемиоценовый долгопят из Таиланда и реконструкция его орбитальной морфологии с использованием геометрическо-морфометрического метода» . Труды Королевского общества B: Биологические науки . 278 (1714): 1956–1963. дои : 10.1098/рспб.2010.2062 . ПМК 3107645 . ПМИД 21123264 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Росси, Дж.Б.; Ни, Х.; Борода, КК (2006). «Черепные остатки эоценового долгопята» (PDF) . ПНАС . 103 (12): 4381–4385. дои : 10.1073/pnas.0509424103 . ПМК 1450180 . ПМИД 16537385 .
- ^ Новак, Р.М. (1999). Млекопитающие мира Уокера (6-е изд.). Издательство Университета Джонса Хопкинса. стр. 94–97. ISBN 978-0-8018-5789-8 .
- ^ Зийлстра, Йелле С.; Флинн, Лоуренс Дж.; Весселс, Вильма (2013). «Самый западный долгопят: новый род и вид из миоцена Пакистана». Журнал эволюции человека . 65 (5): 544–550. дои : 10.1016/j.jhevol.2013.06.015 . ПМИД 23928350 .
- ^ Саймонс, Э.Л. (2003). «Ископаемые записи эволюции долгопятов». В Райте, ПК; Саймонс, Эл.; Гурский, С. (ред.). Долгопяты: прошлое, настоящее и будущее . Издательство Университета Рутгерса. ISBN 978-0-8135-3236-3 .
- ^ Поллок, Дж.И. и Маллин, Р.Дж. (1986). «Биосинтез витамина С у полуобезьян: доказательства родства Tarsius с антропоидами » . Американский журнал физической антропологии . 73 (1): 65–70. дои : 10.1002/ajpa.1330730106 . ПМИД 3113259 . Архивировано из оригинала 28 июня 2012 г. Проверено 16 марта 2010 г.
- ^ Брэндон-Джонс, Д.; и др. (2004). «Азиатская классификация приматов». Международный журнал приматологии . 25 (1): 97–164. дои : 10.1023/B:IJOP.0000014647.18720.32 . S2CID 29045930 .
- ^ Браун, Рэйф М.; Вегхорст, Дженнифер А.; Олсон, Карен В.; Дуйя, Мариано Р.М.; Барли, Энтони Дж.; Дуя, Мелизар В.; Шекеле, Мирон; Нери-Арболеда, Ирен; Эссельстин, Джейкоб А.; Домини, Натаниэль Дж.; Онг, Перри С.; Мориц, Джиллиан Л.; Люсон, Адриан; Дисмос, Мэй Лоу Л.; Дисмос, Арвин К. (19 августа 2014 г.). «Генетика сохранения филиппинского долгопята: загадочные генетические вариации меняют приоритеты сохранения приматов островного архипелага» . ПЛОС ОДИН . 9 (8): е104340. Бибкод : 2014PLoSO...9j4340B . дои : 10.1371/journal.pone.0104340 . ISSN 1932-6203 . ПМК 4138104 . ПМИД 25136854 .
- ^ Jump up to: а б Шекель, Мирон; Гроувс, Колин П .; Марианто, Ибн; Миттермайер, Рассел А. (май 2017 г.). «Два новых вида долгопятов (Tarsiidae, приматы) и биогеография Сулавеси, Индонезия » Сохранение приматов . 31 (1): 1–9.
- ^ Солури, К. Элизабет; Сабрина К. Агарвал (2016). Лабораторное пособие и рабочая тетрадь по биологической антропологии . WW Нортон. ISBN 978-0-393-91291-3 .
- ^ Шумейкер, Роберт В.; Бенджамин Б. Бек (2003). Приматы под вопросом . Смитсоновские книги. ISBN 978-1-58834-151-8 .
- ^ Jump up to: а б с Шекеле, Мирон; Гурский (2010). «Почему долгопяты? Почему сейчас? Введение в специальный выпуск о долгопятах». Международный журнал приматологии . 31 (6): 937–940. дои : 10.1007/s10764-010-9459-6 . S2CID 326565 .
- ^ Расмуссен, Д.Т.; Конрой, GC; Саймонс, Э.Л. (1998). «Локомоторные специализации олигоценового примата Afrotarsius, подобные долгопятам» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (25): 14848–14850. Бибкод : 1998PNAS...9514848T . дои : 10.1073/pnas.95.25.14848 . ПМК 24538 . ПМИД 9843978 .
- ^ Jump up to: а б Нимиц, Карстен (1984). Макдональд, Д. (ред.). Энциклопедия млекопитающих . Нью-Йорк: факты в архиве. стр. 338–339 . ISBN 978-0-87196-871-5 .
- ^ Jump up to: а б с Грон К.Дж. 1 декабря 2010 г. Информационные бюллетени о приматах: таксономия, морфология и экология долгопятов (tarsius). http://pin.primate.wisc.edu/factsheets/entry/tarsier . По состоянию на 12 ноября 2019 г.
- ^ Саймонс, Элвин Л .; Райт, Патрисия К .; Гурски, Шэрон (2003). Долгопяты: прошлое, настоящее и будущее . Издательство Университета Рутгерса. п. 65. ИСБН 0-8135-3236-1 .
- ^ Роза, МГ; Петтигрю Дж. Д.; Купер ХМ (1996). «Необычный образец ретиногенных проекций у спорного примата Tarsius ». Мозг, поведение и эволюция . 48 (3): 121–129. дои : 10.1159/000113191 . ПМИД 8872317 .
- ^ Коллинз, CE; Хендриксон, А.; Каас, Дж. Х. (2005). «Обзор зрительной системы предплюсней ». Анатомические записи, часть A: открытия в молекулярной, клеточной и эволюционной биологии . 287 (1): 1013–1025. дои : 10.1002/ar.a.20263 . ПМИД 16200648 . S2CID 21448186 .
- ^ Рамсьер, Марисса А.; Каннингем Эй Джей; Мориц Г.Л.; Финнеран Джей Джей; Резюме Уильямса; Онг PS; Гурски-Дойен С.Л.; Домини, Нью-Джерси (2012). «Общение приматов в чистом ультразвуке» . Письма по биологии . 8 (4): 508–11. дои : 10.1098/rsbl.2011.1149 . ПМЦ 3391437 . ПМИД 22319094 .
- ^ Фридерун Анкель-Саймонс (27 июля 2010 г.). Анатомия приматов: Введение . Академическая пресса. стр. 442, 521. ISBN. 978-0-08-046911-9 .
- ^ Расти, Нанда; Гурски-Дуайен, Шарон (2010). «Предварительные данные о поведении, экологии и морфологии карликовых долгопятов (tarsius pumilus)». Международный журнал приматологии . 31 (6): 1174–1191. дои : 10.1007/s10764-010-9456-9 . S2CID 23939646 .
- ^ Мюссер, Г.Г.; Дагосто, М. (1987). «Особенность Tarsius pumilus, карликового вида, эндемичного для горных мшистых лесов Центрального Сулавеси». Новитаты Американского музея (2867 г.): 1–53.
- ^ Дагосто, М.; Гебо, ДЛ; Долино, К. (2001). «Позиционное поведение и социальная организация филиппинского долгопята ( tarsius syrichta )». Приматы . 42 (3): 233–243. дои : 10.1007/bf02629639 . S2CID 41499839 .
- ^ Нимиц, К. (1977). «О функциональной морфологии и биометрии рода Tarsius, Storr, 1780». Курьерский научно-исследовательский институт Зенкенберга . 25 :1–161.
- ^ Нимиц, К. (1979). Взаимосвязь между анатомией, экологией и поведением: модель, разработанная для рода Tarsius, с мыслями о филогенетических механизмах и адаптивных взаимодействиях. В S. 1190 Н. Гроу, С. Гурски-ДойенМорбек, Х. Преушофт и Н. Гомберг (ред.), Окружающая среда, поведение и морфология: динамические взаимодействия (стр. 119–138). Нью-Йорк: Густав Фишер.
- ^ Нимиц, К. (1984). Исследование и обзор территориального поведения и социальной организации рода Tarsius. В К. Нимице (ред.), Биология долгопятов (стр. 117–128). Нью-Йорк: Густав Фишер
- ^ Кернер, К. (2007). «Использование высоты в экологических исследованиях». Тенденции экологии и эволюции . 22 (11): 569–574. дои : 10.1016/j.tree.2007.09.006 . ПМИД 17988759 .
- ^ Рэй, ТК; Хилл, Род-Айленд; Хамада, Ю.; Коппе, Т. (2003). «Клинические изменения объема верхнечелюстной пазухи у японских макак (Macaca fuscata)». Американский журнал приматологии . 59 (4): 153–158. дои : 10.1002/ajp.10072 . ПМИД 12682923 . S2CID 12290499 .
- ^ Ярош, Энди (20 мая 2019 г.). «Филиппины: Долгопят» . Нэшнл Географик . Архивировано из оригинала 21 мая 2019 г. Проверено 28 сентября 2019 г.
- ^ Синклер, Джо (12 декабря 2011 г.). «Туризм угрожает крошечному филиппинскому примату» . Мой СинЧью . АФП . Архивировано из оригинала 28 сентября 2019 г. Проверено 28 сентября 2019 г.
- ^ Ржегакова-Петру, М.; Пешке, Л. (2012). «Хищничество дикого филиппинского долгопята (Tarsius syrichta)». Акта Этологика . 15 (2): 217–220. дои : 10.1007/s10211-011-0096-7 . S2CID 254163428 .
- ^ Гурски, Шэрон (февраль 2005 г.). " "Хищник-банда в Тарсиус Спектр" " . Международный журнал приматологии . 26 (1): 207–221. дои : 10.1007/s10764-005-0731-0 . S2CID 21188050 — через EBSCOhost.
- ^ Кромптон, Робин Хью; Бланшар, Мэри Л.; Трус, Сэм; Александр, Р. Макнил; Торп, Сюзанна К. (1 декабря 2010 г.). «Возвращение к вертикальному цеплянию и прыжкам: передвижение и использование среды обитания западного долгопята, Tarsius Bancanus, исследованные с помощью логлинейного моделирования» . Международный журнал приматологии . 31 (6): 958–979. дои : 10.1007/s10764-010-9420-8 . ISSN 1573-8604 . S2CID 45884124 .
- ^ Кромптон, Робин Хью; Сэвидж, Рассел; Спирс, Иэн Р. (14 февраля 1998 г.). «Механика сокращения потребления пищи у Tarsius Bancanus» . Фолиа Приматологическая . 69 (7): 41–59. дои : 10.1159/000052698 . ISSN 1421-9980 . ПМИД 9595687 . S2CID 24464173 .
- ^ Шахрулла, Фахри Науфал; Маддус, Ун; Мустари, Абдул Харис; Гурски, Шэрон; Индраван, Мохамад (15 июля 2023 г.). «Распространение и численность пеленгского долгопята (Tarsius pelengensis) в группе островов Банггай, Индонезия» . Научные отчеты . 13 (1): 11445. Бибкод : 2023NatSR..1311445S . дои : 10.1038/s41598-023-30049-5 . ISSN 2045-2322 . ПМЦ 10349819 . ПМИД 37454197 .
- ^ Изард, Кей М.; Райт, Саймонс (1985). «Продолжительность беременности Tarsius Bancanus» Ам Дж Приматол . 9 (4): 327–331. дои : 10.1002/ajp.1350090408 . ПМИД 31979510 . S2CID 83711759 .
- ^ Робертс, М.; Кон, Ф. (1993). «Использование среды обитания, поведение при добывании пищи и модели деятельности при воспроизводстве долгопятов западных, Tarsius Bancanus , в неволе: синтез управления» (PDF) . Зоопарковая биология . 12 (2): 217–232. дои : 10.1002/zoo.1430120207 . Архивировано из оригинала (PDF) 12 марта 2012 г. Проверено 19 ноября 2010 г.
- ^ Шекеле, М.; Нич, А. (2008). Шекеле, М.; Мариано, Т.; Гроувс, К.; Шульце, Х.; Фитч-Снайдер, Х. (ред.). Долголетие долгопятов: данные повторной поимки в дикой природе и животных в неволе (PDF) . ЛИПИ Пресс. стр. 85–89. ISBN 978-979-799-263-7 . Архивировано из оригинала (PDF) 28 июля 2011 года.
{{cite book}}:|work=игнорируется ( помогите ) - ^ Северн, К.; Даханг, Д.; Шекеле, М. (2008). Шекеле, М.; Мариано, Т.; Гроувс, К.; Шульце, Х.; Фитч-Снайдер, Х. (ред.). Восточные долгопяты в неволе, Часть I: Вольер и обогащение (PDF) . ЛИПИ Пресс. стр. 91–96. ISBN 978-979-799-263-7 . Архивировано из оригинала (PDF) 24 июля 2011 года.
{{cite book}}:|work=игнорируется ( помогите ) - ^ Северн, К.; Даханг, Д.; Шекеле, М. (2008). Шекеле, М.; Мариано, Т.; Гроувс, К.; Шульце, Х.; Фитч-Снайдер, Х. (ред.). Восточные долгопяты в неволе, Часть II: Предварительная оценка рациона (PDF) . ЛИПИ Пресс. стр. 97–103. ISBN 978-979-799-263-7 . Архивировано из оригинала (PDF) 28 июля 2011 года.
{{cite book}}:|work=игнорируется ( помогите ) - ^ Фитч-Снайдер, Х. (2003). «История сохранения долгопятов в неволе». В Райте, ПК; Саймонс, Эл.; Гурский, С. (ред.). Долгопяты: прошлое, настоящее и будущее . Издательство Университета Рутгерса. стр. 227–295. ISBN 978-0-8135-3236-3 .
- ^ «Стивен МБланд» . Проверено 18 октября 2016 г.
- ^ Ячовски, Дэвид С.; Пиццарас, Карлито (2005). «Представляем инновационную среду полув неволе для филиппинского долгопята ( Tarsius syrichta )». Зоопарковая биология . 24 (1): 101–109. дои : 10.1002/zoo.20023 .
- ^ Войцеховский, Филип Дж.; Кашицка, К.А.; Вильбасс, AM; Ржегакова, М. (2019). «Модели деятельности содержащихся в неволе филиппинских долгопятов (Tarsius Syrichta): различия, связанные с полом и социальным контекстом». Фолиа Приматологическая . 90 (2): 109–23. дои : 10.1159/000495612 . ПМИД 30826810 . S2CID 73491766 .
- ^ Шекеле, Мирон; Салим, Агус. «Домингом с острова Сиау» . Группа специалистов МСОП/SSC по приматам. Архивировано из оригинала 6 сентября 2010 года . Проверено 1 января 2010 г.
- ^ «Полностью охраняемые животные Саравака» . Департамент лесного хозяйства Саравака . Проверено 1 января 2010 г. [ постоянная мертвая ссылка ]
Внешние ссылки
[ редактировать ]- Tarsier.org. Архивировано 16 января 2022 г. в Wayback Machine , международном исследовательском и природоохранном проекте.
- Долгопяты (Tarsiidae) в Wayback Machine (архивировано 15 октября 2004 г.), Доценты Сингапурского зоологического сада, 2000 г.
- Грон, Курт Дж. (июль 2008 г.). «Информационные бюллетени о приматах: таксономия, морфология и экология долгопятов (tarsius)» . Национальный центр исследования приматов, Университет Висконсин-Мэдисон.
- Долгопяты – Посещение двух заповедников долгопятов на Бохоле, Филиппины.
- Скелет долгопята – скелет из Техасского университета в Остине.