Enhydriodon
| Enhydriodon Временный диапазон: поздний миоцен до раннего плейстоцена ,
| |
|---|---|
| |
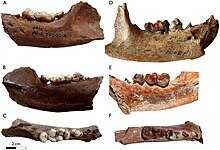
| Нижние зубные зубные зубные зубные зубцы Enhydriodon Hendeyi (AC) и Enhydriodon Africanus (DF) | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Class: | Mammalia |
| Order: | Carnivora |
| Family: | Mustelidae |
| Subfamily: | Lutrinae |
| Tribe: | †Enhydriodontini |
| Genus: | †Enhydriodon Falconer, 1868 |
| Type species | |
| †Enhydriodon sivalensis Falconer, 1868
| |
| Other species | |
| |
| Synonyms | |
|
Genus synonymy Synonyms of E. sivalensis Synonyms of E. ekecaman | |
Enhydriodon - вымерший род мустелидов , известных из Африки , Пакистана и Индии , который жил от позднего миоцена до раннего плейстоцена . Он содержит девять подтвержденных видов , два обсужденных вида и, по крайней мере, несколько других неписанных видов из Африки. Род принадлежит к племени Enhydridontini (который также содержит Sivaonyx и Vishnuonyx ) в подсемействе выдр . Enhydriodon означает «зуб выдры» на древнегреческом языке и является ссылкой на его зубной зуб, а не на род Enhydra , который включает в себя современную море и два доисторических родственника.
Точные размеры и длина видов Enhydriodon неизвестны, учитывая отсутствие полных окаменелостей и большинство связанных ископаемых лютринов. По оценкам, индийские субконтинентальные виды имеют вес, аналогичные существующей морской выдре , но африканские виды, по оценкам, являются более тяжелыми, чем существующие лютрины. В частности, несколько видов, таких как E. Kamuhangirei , E. Dikikae и E. Omoensis , были оценены весом более 100 кг (220 фунтов). Учитывая эти оценки веса, три вида, вероятно, достигли размеров, сравнимых с существующими медведями или львами , что делает их самыми крупными известными мусталидами, хотя отсутствие полных образцов делает точные оценки невозможными.
Its advanced dentition is well-known, its broad, bunodont carnassials allowing the lutrine to consume prey by crushing them rather than shearing them like the modern sea otter and unlike most other extant otters. As such, it is grouped among the bunodont otters, a categorical term referring to fossil lutrines with non-bladelike carnassials in the premolars or molars of the Miocene to Pleistocene and the sea otter of the sole extant Enhydra genus. Its I3 teeth (or third incisors) are canine-like and much larger than its other incisors (although shorter than its canines), a trait not seen in extant and extinct lutrine genera. It is hypothesized that Indian species of Enhydriodon were semiaquatic and consumed bivalves because their bunodont dentitions would have allowed them to consume hard-shelled invertebrates. It is unknown whether African species were generally aquatic, semiaquatic, or terrestrial, but their potential diets suitable for bunodont dentitions include bivalves, catfish, reptiles, eggs, and carrion. E. omoensis of Ethiopia in particular could have been a terrestrial locomotor that at least semiregularly hunted or scavenged terrestrial prey with C4 plant diets which if true makes its behaviour unlike any extant otters. It is unknown whether the species is an outlier amongst African bunodont otter species, but it has been suggested that Enhydriodon dikikae and Sivaonyx beyi were both large terrestrial bunodont otters of Africa as well.
The taxonomic status of Enhydriodon species have been complicated by its affinities and similarities with other bunodont lutrine genera like Sivaonyx and Paludolutra up to the modern day, although Paludolutra is presently considered a distinct genus not closely related to Enhydriodon. Currently, the Enhydriodontini tribe is considered evolutionarily closer to the modern Enhydra genus than any other known bunodont otter genus that may have gained bunodont dentition as a result of parallel evolution, but the extent to which they are closely related remains unresolved.
Taxonomy
[edit]Early history
[edit]
Enhydriodon was first erected in 1868 by Hugh Falconer based on several crania from Siwalik Hills, India that he attributed to E. sivalensis.[1][2] He explained that the scientific name, meaning "otter tooth", is derived from the Ancient Greek terms ἐνυδρίς (otter) and ὀδούς (tooth) and is not a reference to the genus Enhydra, which includes the modern sea otter (Enhydra lutris). According to Falconer, the Siwalik Hill fossils belonging to E. sivalensis were previously classified by Falconer and Proby Cautley under the genus and species name Amyxodon sivalensis in an 1835 synopsis of the fossil genera in the Siwalik Hills that the two palaeontologists found, in which the fossil taxon was considered to be a carnivoran of an unknown family, although no holotype or diagnostic descriptions were attributed to it. As a result of the rename, Amyxodon had been considered a "dead name" or synonym of Enhydriodon despite being the older genus name. Using the available specimens of E. sivalensis, Falconer calculated that there were four premolars and molars in the species's upper jaw (the dental formula was calculated as 3.1.42.1.5), one less than in the extant genus Lutra but matching the total count of Enhydra. He described the upper carnassial of E. sivalensis as the most unique feature of its upper jaw, being nearly square and its coronal lobes being developed from conical mamelons unlike the two extant lutrine genera.[1][3]
В течение 19-го и 20-го веков больше видов энфидриодона, таких как E. campanii было введено , и было описано больше родов Luttrine с зубным протезом Bunodont , такими как Sivaonyx и Vishnuonyx , создавая особенно сложную историю для самого раннего принятия доисторического рода. В 1931 году Гай Пилигрим описал больше окаменелостей, обнаруженных на холмах Сивалик, в том числе более новые виды по имени Э. Фалконери . Он также подразумевал, что Enhydriodon и Sivaonyx , несмотря на их сходство, были дифференцированы структурой верхнечелюстного 4 -го премоляра (P 4 ) и очевидное отсутствие переднего верхнего премоляра (P 1 ) это предполагается, что это также отражено в нижней челюсти (оба из которых обсуждаются до сегодняшнего дня). [ 4 ] В том же году, когда E. Falconeri был описан, Эрнст Стромер описал E. africanus позднего плиоцена , его ископаемые зубы расположены в Южной Африке и первые описанные виды с континента Африки. [ 5 ]
Воспринимаемые отношения с Палудолатрой и Энфидрой
[ редактировать ]В 1976 году Чарльз Repenning вызвал идею о том, что Enhydriodon был связан с существующим родом Enhydra из-за предполагаемого вида первого, являющегося эволюционным «ветвью» «крабовых выдр» в Италии, Испании и Калифорнии, в конечном итоге приводя к Современная морская выдра. [ 6 ] Он правильно представил идею о том, что Enhydra была связана с Enhydriodon , учитывая их зубные зубные узоры, но предполагаемое европейское «ветвь» рода Enhydriodon была позже реклассифицирована Йоханнесом Хюрзелером и Буркартом Энгессером в более новый род Paludolura в 1976 Палеонтологические записи до тех пор, пока более позднее исследование пересмотрело его таксономическое состояние. [ 7 ] [ 8 ]
Таксономии отдельных видов и родов Lutrine продолжали пересматриваться в 21 -м веке, поскольку были описаны более доисторические выдры, в то время как палеонтологи постоянно пересматривали ископаемые виды Luttrine Bunodont в разные роды. Палудолатра был первоначально реклассифицирован как Джерардом Ф. подрод Enhydrodon Виллемен в 1992 году. [ 5 ] Однако в январе 2005 года Мартин Пикфорд и его коллеги поставили диагноз Палудолатра как синоним Sivaonyx на основе диагноза Паломника последнего, отвергая синонимию Виллемсена о Палудолатре в Энхидрионе . Кроме того, они установили вид Enhydriodon по имени Э. Хенддей из типа местности Лангебаанвега , Южная Африка, который датируется нижним плиоценом и был назван в честь палеонтолога Куинтона Б. Хенди, который, как они сказали, описал первые известные образцы, которые были с тех пор, с тех пор были приписывается видам. [ 9 ] В декабре того же года Хорхе Моралес и Пикфорд вместо этого описали Палудолатру как отдельный род, который может быть связан с Sivaonyx , основанным на конвергенциях зубного зуба. [ 8 ] В 2007 году два палеонтолога подтвердили, что зубная морфология палудолатры была достаточно различна , чтобы быть реклассифицированным как род, основанный на полной общей дифференциации, что позволяет предположить, что виды P. campanii , P. lluecai и P. Maremmana больше не будут классифицированы под Enhydriodon в основе Палудолутра является подродом. [ 10 ] [ 11 ]
Современные пересмотра африканских видов
[ редактировать ]
В 2003 году Ларс Верделин вид Э. Экекамана из Канапоя палеонтологического сайта бассейна Туркана ранний плиоцена, ок. установил в Кении ( Этот вид был назван в честь термина «Экекаман» Туркана «Экекаман», что означает «рыбак», потому что он предположил, что рыба могла быть диетой для этого вида. Он также объявил виды « Э. Паттерсони », описанный RJG Savage в 1978 году, как номенсту E. ekecaman , поскольку в декабре 2005 года не было назначено образец или действительный диагноз. [ 12 ] [ 8 ]
E. Africanus , E. Ekecaman и E. Hendeyi были переклассифицированы в Sivaonyx Пикфордом и Моралесом в декабре 2005 года, где они дополнительно описали новый вид под названием Sivaonyx Kamuhangirei . [ 8 ] Реклассификация африканских ископаемых выдр в Sivaonyx привела к непрерывной дебатах о практичности различий между Enhydriodon и Sivaonyx , причем некоторые исследователи претендуют на нейтралитет из -за предпочтительного фокуса на изучении отдельных видов, а не на их роде. В 2022 году четыре вида были в конечном итоге реклассифицированы в Enhydriodon в исследовательской статье Camille Grohé et al. E. soriae также была изначально отсортирована на Sivaonyx, но в конечном итоге был назначен Enhydriodon , хотя его родовое размещение остается оспоренным. [ 13 ] [ 14 ] В 2005 году Моралес и Пикфорд отсортировали Enhydriodon в недавно созданном племени Enhydroidontini , которое они назвали, как принимая роды вымерших выдр из бундодонтов из холмов Сивалик и Африки, включая Vishnuonyx , Sivaonyx и Paludolura . В 2007 году Пикфорд синонимизировал вид « E. aethiopicus », ранее описанный Denis Geraads et al. В 2004 году для Pseudocivetta Ingens , вымершего члена семейства Viverridae . [ 10 ] В 2017 году Enhydra была явно исключена из племени Enhydroidontini, несмотря на его сходство, а Палудолатра был реклассифицирован как сестринский таксон для племени. [ 8 ] [ 11 ]
В 2011 году Денис Гераадс и его коллеги описали E. dikikae на основе его остатков частичного черепа и бедра в нижней части Дикики , Эфиопии, местности, датируемого средним плиоценом. Он был описан как имеющий особенно более тяжелый череп (хотя и сломан), чем другие виды Enhydriodon или современная морская выдра. Названный вид был основан непосредственно на месте Дикики. [ 15 ] Он считался самым большим видом усиления энхидриодов до тех пор, пока в нижней долине Омо также был описан другой вид из Эфиопии, E. Omoensis , датируемого поздним плиоценом до границы плиоплеистоцена . Подобно E. dikikae , название вида было получено непосредственно с места, в котором он был извлечен. [ 14 ] На конференции в сентябре 2022 года Альберто Валенсиано, Моралеса и Пикфорда (в том же месяце, что и исследовательский документ о E. Omoensis ), однако они ссылались на определенные виды Luttrine, ранее реклассифицированные в Enhydriodon как Sivaonyx , а также S. hendeyi и S. africana Полем [ 16 ]
Классификация
[ редактировать ]
Enhydriodon принадлежит к племени Enhydridontini в подсемействе Lutrinae , которая впервые появилась в Евразии и Африке во время поздней эпохи миоцена . [ 8 ] Это, пожалуй, самая известная доисторическая выдр, учитывая ее старую таксономическую историю, и она является основным источником сравнений с другими родами выдры Бундонта. Обычно считается, что Enhydriodon был результатом миоцен-плейстоценовой тенденции, которая давала доисторические лучи-зубы Bunodont и большие размеры по сравнению с их существующими родственниками. Он классифицируется как член группы Bunodont Otters, категориальный термин, обычно используемый исследователями, который также включает в себя Sivaonyx , Paludolutra , Vishnuonyx , Torolutra , Enhydrithritherium , Djourabus , Paralutra , Tyrrhenolutra , Siamogale и Enhydra . [ 11 ] [ 17 ] [ 18 ] Выдры Бундонта определяются как большие до очень больших муфелидов из Северной Америки, Евразии и Африки, у которых был надежный зубной зуб по сравнению с большинством существующих выдр, что обычно позволяет им охотиться за жестким вооружением. [ А ] [ 13 ] [ 17 ] Несмотря на то, что он разделяет особенность зубного зубного зуба Bunodont, существует как минимум несколько клад лютринов, принадлежащих к этой категории, а не к одному, поэтому термин «выдры бундодонта», следовательно, является категориальным, а не таксономическим и покрывает латрины в те же периоды с одинаковыми зубными зубными Это напрямую определяет их таксономическое состояние. [ 11 ]
Следующая кладограмма Xiaming Wang et al. В 2018 году определяет некоторые из следующих существующих и вымерших видов выдр и родов в рамках подсемейства Lutrinae на основе консенсуса большинства в 50% (роды выдр Bunodont, начиная с « Paralutra Jaegeri »): [ 11 ]
| Lutrinae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Как показано в вышеупомянутой филогении, Enhydriodon разделял более близкую морфологию с другими ее вымершими родственниками и Enhydra, чем другие существующие лютрины, в которых отсутствует карнассиальные зубы Bunodont ( Lutra aonychoides были описаны как не связанные с Lutra ). Несмотря на то, что согласованное дерево большинства демонстраций демонстрирует тесную морфологическую связь между Enhydriodon и Enhydra , авторы консенсусного дерева также создали байесовское дерево вывода, предлагающее, чтобы Enhydra в качестве изолированной клады отделяется от типичных членов Enhydridontini (« Паралутра » членов была предложена в качестве изолированных Клада из Siamogale также). Несмотря на это, они утверждали, что Enhydra ближе к кладке, сочиняющей Enhydriodon , Sivaonyx и Vishnuonyx , чем любой другой род Bunodont Выдр. Исследователи объяснили, что приобретение зубного проведения Бундонта происходило не менее трех раз в эволюции лютринов, отраженных кладами типа филогения: в Sivaonyx - Enhydriodon - Enhydra , в Палудолатре - Enhydritherium и в Siamogale . [ 11 ] Выдры, не являющиеся бунодонтом, вероятно, разветвлялись отдельно от выдр-бундонтов в эпоху плиоцена, но их плохие ископаемые записи и ограничение от отложений плио-плейстоцена по сравнению с эволюционными филогениями мало понимают. [ 20 ]
Описание
[ редактировать ]Череп
[ редактировать ]
В настоящее время существуют только два известных частичных черепа, которые приписываются Enhydriodon : один из E. sivalensis из холмов Сивалика, а другой из E. dikikae из долины Awash. В настоящее время неизвестно, являются ли особенности черепа любого вида хорошо представляют другие виды Enhydriodon , но известные черепа E. dikikae и E. sivalensis имеют несколько отличающиеся друг от друга. [ 15 ]
Череп E. sivalensis , идентифицированный как принадлежащий полностью выращенному человеку, относительно хорошо сохранился с идентифицируемыми височными гребнями , лобными частями , максиллами , предчексиллами , носовой , морной и палатиновой костью . Тем не менее, он также страдал от износа и слегка скрученной по часовой стрелке. В частности, зубная арка завершена, хотя левая М 1 и оставил я 1 оба отсутствуют, и большинство зубов разбиты из их коронок. Он имеет большой корпус мозга, широкий и короткий дуло и большое носовое отверстие. Контуры орбит вокруг лобных черепов также могут быть идентифицированы. [ 2 ]
Сломанный череп, принадлежащий E. dikikae, содержит короткую и непрогнирующую морду, части орбит, почти полная верхняя зубная арка, которой отсутствует оба я 1 S и право я 2 и часть нижней челюсти. Морда на черепе E. dikikae короткая, небольшая передняя орбитальная граница, расположенная чуть выше задней стороны собаки . Передняя часть морды идентифицируется как короткая, тем самым сравнимая с мордой Enhydra . Несмотря на то, что эволюция таких выдр, таких как Enhydriodon, неясна, предполагается, что короткая морда E. dikikae и очень большой размер собаки, которые явно делают вид различным и более эволюционно полученным (или эволюционно недавно), чем E. sivalensis . [ 15 ]
Зубной зуб
[ редактировать ]Уточечная зуба Enhydriodon хорошо определена его чрезвычайно широкими карнассиальными бирнами в молярах и премолярах, аналогичных современной море. Различия видов Enhydriodon . и Sivaonyx обычно приписываются зубным зубам, поэтому предмолярные зубы или окаменелости молярных зубов рассматриваются для различения двух родов Bunodont Общие различия (более крупные p 4 Гипоконы, конические постпротоконные бугорки и очевидное отсутствие передних верхних премоляров для энмидриодона ) измерениями зубов было трудно доказать из-за фрагментарной природы окаменелостей и относительных несоответствий измерений/измерений зубов по видам. [ 10 ] [ 15 ] Реклассификация всех видов «африканского sivaonyx », кроме S. beyi , в Enhydriodon в 2022 году, была отнесена к «[a] метакониду выше протоконида на М 1 , наличие карнассиальной выемки и один или несколько островов между протоконом и Гипокон на р 4 , и/или дистолингальное расширение на М 1 . " [ 14 ]
Предполагается, что Enhydriodon как последний род, который имеет наибольшее количество зубов Bunodont племени Enhydroidontini, которое включает в себя самые ранние вишнуоникс , а затем Sivaonyx . Уточечная зуба Enhydroidon предполагает почти подавление карнассиальных функций в пользу сокрушительного в качестве преобладающей функции. I. 3 (или третий верхний резчик ) Enhydriodon намного больше, чем его I 1 (наименьший резчик) и я 2 , выглядя более крупнее и более качественным собачьим по сравнению с Палудолатрой и Энфидрой . По сравнению с другими родами Lutrine Bunodont, где известен верхний резчик, его третьи резцы лишь незначительно больше, чем их первые и вторые резцы. [ 10 ] Право я 1 Черепа E. sivalensis, например, измеряет 3 мм (0,12 дюйма) в переднепостериальном диаметре (APD) и 4,5 мм (0,18 дюйма) поперечного диаметра (TD). Череп прав, я 2 Измеряют 5,2 мм (0,20 дюйма) в APD и 5,5 мм (0,22 дюйма) в TD. Для сравнения, право я 3 является крупнейшим резцом голотипа с измерениями 10,5 мм (0,41 дюйма) в APD и 8 мм (0,31 дюйма) в TD (клыки больше резцов, измеряя 17,1 мм (0,67 дюйма) в APD и 13,8 мм ( 0,54 дюйма) в TD). [ 2 ] Большой я 3 Черта также применима к E. dikikae , который был описан после общего описания Пикфорда энфидриодона как имеющего гораздо больший I 3 чем я 1 - я 2 и быть более коническим по форме. Dik-56's i 3 Зуб измеряет 12,4 мм (0,49 дюйма) по мезиодистальной ширине (MD) и 11,6 мм (0,46 дюйма) в ширине щечной линии (BL) по сравнению с его I 2 Измерения 5,5 мм (0,22 дюйма) в MD и 9,7 мм (0,38 дюйма) в BL. Как Э. Сиваленсис , я 3 короче, чем собаки, с c 1 измерение 16,9 мм (0,67 дюйма) в MD плюс 15 мм (0,59 дюйма) в BL и C 1 измеряет 19,5 мм (0,77 дюйма) в MD и 15,3 мм (0,60 дюйма) в BL. [ 15 ]
Конечности
[ редактировать ]
Посткраниальные останки выдр Бундонта, в том числе Enhydriodon , мало, оставляя слишком мало информации об общей анатомиях многих родов. Единственными известными видами усиленного энхидриона с посткраниальными останками являются Э. Хендий , Э. Дикика и Э. Омоэнсис . [ 13 ]
E. Hendeyi Fossil Onkil включает фрагментарную плечевую кость , локтевую кость , два бедра и астрагалу (также известный как кость талус). Бедра Э. Хендей меньше, чем у С. Бейи и Э. Дикика, но также больше, чем у Энидритера , Сатериума и существующей африканской выдры. Astragalus похож на E. omoensis, но отличается меньшей головой и более тонкой шеей, а также более крупной дистальной проекцией бугорки кости . ) является мелкой и широко широко суставы костей Трохлея кости талусной кости (катящая поверхность, образующая широко Расширение ноги на лодыжке по сравнению с африканской выдрой. [ 13 ]
Посткраниальные останки E. dikikae известны проксимальной (верхней частью) левой бедренной кости, дистальной (нижней части) правой бедренной кости и плечевой кости. Проксимальная левая бедренная кость известна большим бугорком вдоль задней области шеи, среднего возрастающего трохантера и аналогичного положения медиального мыщелка бедра . Образцы бедра указывают на большие общие размеры бедра E. dikikae по сравнению даже с самыми крупными существующими выдрами. Компания (полная, но выветритная и потрескавшаяся) гораздо длиннее и немного надежна по сравнению с плечами Sivaonyx Beyi , и его дельтовидная бугристость хорошо сформирована. Латеральный супракондиларный гребень длиннее по сравнению с S. beyi , в то время как медиальный эпикондиль не такой заметный по размеру. Фосса по олекранонам маленькая и более круглая по сравнению с S. beyi . [ 15 ]
E. Omoensis представлен только одним полным левым бедром, у которого есть короткая шея и круглая головка, которая ориентирована в проксимальном направлении (недалеко от центра), а не в медиальном направлении (в центре), причем первое смещается при 40 ° относительно долготы срезы диафиза кости. Ширина латерамедиального эпифиза узкая . В бедренной кости также есть большая бедренная головка расположенная на вентромедиальной головке (выровняясь к ее средней нижней стороне) вертранс больший , , Трохантер, который центрируется больше посередине, чем на брюшной (или нижней стороне) и тем самым видимо в обратном виде. Медиальный мыщелок бедра больше, чем боковой мыщелок той же кости. Межконциларная ямка бедра прямоугольная и широкая. [ 14 ]
Масса тела
[ редактировать ]
Некоторые виды Enhydriodon , особенно некоторые, которые проживали в Африке, являются крупнейшими известными мусталидами , когда -либо существовавшими на основе оценки веса, но их точные размеры и веса остаются неизвестными, учитывая отсутствие полных образцов в их ископаемых записях. Некоторые виды, такие как E. latipes (?) . [ 21 ] Обычно подсчитано, что некоторые виды энфидриодона имеют одинаковые по весу для современных выдр большого размера, в то время как другие оцениваются гораздо больше, чем они (также следует отметить, что оценки веса чаще всего производятся для выдров Бундонта, таких как Enhydriodon , чем оценки размера, оценки размера, оценки размера, оценки размера, оценки размера, оценки размера, оценки размера, оценки размера, оценки размера, оценки размера, оценки размера, оценки размера, оценки размера, оценки размера, оценки размера, оценки размера, оценки размера, оценки размера, оценки размера, оценки размер, Хотя сравнение размеров с современными животными можно ссылаться). [ 14 ]
У двух видов усиления энфидриона, уроженцев субконтинента Индии, были скромные оценки веса, сравнимых с большинством других родов выдр Бундонта, а также существующими родами выдры. Мемуары Falconer 1868 года описали E. sivalensis как лютрин размером с пантера . [ 1 ] В 1932 году Пилигрим диагностировал Э. Фалконери как меньше, чем E. sivalensis , хотя для него не было предложено никаких оценок размера или веса. [ 4 ] В 2007 году Пикфорд оценил E. sivalensis как крупнейшую доисторическую выдру в Индии, в диапазоне от 22 кг (49 фунтов) до 25 кг (55 фунтов) максимума по массе тела, его череп, возможно, имеет значение для волки. Он также оценил тело E. falconeri на основе его более низких размеров зубов M 1 , чтобы быть похожим на африканскую выдру без когтя ( A. capensis ), в среднем до 16 кг (35 фунтов). [ 10 ]
в Африке Предполагается, что виды Enhydriodon являются одними из крупнейших видов выдр, которые когда-либо существуют, отражая миоцено-плейстоценовую тенденцию, растущие выдры Бундонта, превышающие их кузены без бунодонта. Пикфорд описал Э. Камухангирея из Западной Рифт-Вэлли, Уганда (в то время в весах Sivaonyx kamuhangirei ), возможно, превышает 100 кг (220 фунтов), что делает его самой большой доисторической выдрой в то время, хотя он упомянул, что неписанный Выдры в Эфиопии (вероятно, отсортированы позже под E. dikikae и/или E. Omoensis ), возможно, были бы больше, чем это. [ 10 ] E. Hendeyi (тогда Sivaonyx Hendeyi ), по оценкам, размером с волков и около 40 кг (88 фунтов), в то время как E. Africanus и E. Ekecaman считаются аналогичными размерами. [ 22 ] [ 14 ] E. dikikae of Ethiopia, по оценкам, имела минимальный минимальный и 200 кг (440 фунтов) составляет 100 кг (220 фунтов) и 200 кг (440 фунтов) (последний, упомянутый более вероятно), его голотип указывает на медвежье. По сравнению с большинством других видов Enhydriodon или Enhydra , он имел предполагаемую длину черепа около 25 см (9,8 дюйма). [ 15 ] E. Omoensis позже, по оценкам, весит более 200 кг (440 фунтов), что делает его тяжелее E. dikikae и современных львов . Согласно Grohé et al., E. Omoensis потенциально «размером с льва», что делает его самым большим видом Mustelid, когда-либо существовавшим. [ 14 ]
Палеобиология
[ редактировать ]
Поскольку роды выдры ископаемого бундодонта, включая Enhydriodon, обычно не имеют полных образцов и посткраниальных элементов, их локомоция и экологические ниши остаются неопределенными. Общая теория индийских субконтинентальных видов Enhydriodon заключается в том, что, основываясь на их надежном зубном протезе Bunodont, аналогичной Enhydra , E. Falconeri и E. sivalensis, были специализированы для обычно употребления моллюсков . [ 5 ] Это утверждение было сделано первым Виллеменом из аналогий диеты Enhydra (абалоны и морских бивали) и Aonyx (пресноводные крабы), но существует мало палеонтологических доказательств, чтобы непосредственно подтвердить это утверждение. Независимо от того, предполагается, что толстая эмаль в заднем зубном зубном виде индийских энфидриодонов делает их более цветными, чем Cancrivivory (напротив, предполагается, что индийские виды Sivaonyx , в общих коронах Bunodont, чтобы добывать на терковых лодках, с крон bunodont, чтобы добывать на раковых лодках, на терковых лодках, на терковых лодках, ракообразные, на ракообразных, на ракообразных, на терпках на терковых классах имеют комбинированные функции сдвига карнассиальных с короками Bunodont, чтобы добывать на раковых заводах , Хотя двустворчатые моллюски могли быть вторичной добычей для этого). Возможность поживания в ура усилении племени энхидриодонтини на индийском субконтиненте (Индия и Пакистан). [ 10 ]
Предполагается, что более крупные виды Enhydriodon на африканском континенте охотились на более широкое разнообразие пищевых продуктов в дополнение к их основной добыче, включая более мягкую добычу, несмотря на их зубные зубные зубные уборы, что отличает их потенциальные диеты от их индийских субконтинентальных коллег. Одним из предполагаемых типа добычи была большая рыба с твердыми внешними покрытиями, такими как сома . [ 23 ] Несколько родов сома присутствовали в Африке, начиная с их первых появлений во время позднего миоцена, совпадающего с наличием Enhydriodon , включая существующие роды Clarotes , Bagrus , Auchenoglanis и Chrysichthys и вымерший род Nkondobagrus . [ 24 ] В отличие от медленного, обильного сома, крабов в Африке были исключены в качестве потенциальной добычи для африканских видов энхидриодона, учитывая отсутствие окаменелых крабов в Дикике, маловероятность биомасс зубной зуб. Быстро плавающая рыба, возможно, вряд ли была бы обычными источниками пищи из-за специализированного зубного проведения для сокрушительного пищи, в дополнение к крупным животным, вероятно, не имеющим возможности улавливать добычу. Другая бронированная добыча, такая как юношеские крокодилы, черепахи и страусные яйца, также была предложена добычей E. dikikae . [ 15 ]

Бедренные и стоматологические остатки африканского энхидриона, возможно, намекают как на полуакватический, так и на земнный образ жизни, что означает, что он мог съесть как добычу водную добычу, так и земля. Однако предположения о образе жизни Энчераодона были противоречивы друг другу, поэтому на нем нет консенсуса большинства. В 2008 году предполагалось, что меньшие африканские виды Enhydriodon на основе их меньших размеров бедренной кости были более локомоторными универсалами, похожими на большинство мустлидов, в то время как более крупные виды были полностью водными, поскольку их структуры бедренной кости разделяют сходство с Enhydra . Тем не менее, проксимальные концы Omo и Hadar Femoras указывали на более водный характер, чем большинство лютринов, в то время как их относительные длины напоминали общие обобщенные мастера, включая полуакватические выдры. [ 23 ] [ 14 ] Sivaonyx Beyi из Chad , предположительно весить 56,4 кг (124 фунта) до 60,1 кг (132 фунта), имели неспециализированные пропорции конечности, которые подразумевали общепринятые локомоции и плохие водные адаптации. [ 25 ] Из -за гипотезы о том, что S. beyi был наземным хищником, E. dikikae предположительно, в основном был наземным на основе ее общего расположения окаменелостей как с водной, так и с наземной фауной в Дикике. [ 15 ] Палеобиологии и нишевые разделы Э. Экекамана и Э. С. С. Dikikae в Канапои , Кения, остается неясной, поскольку их ископаемые материалы, обнаруженные в 1960 -х годах, не были специально произнесены за пределы «Канапоя», которые будущие исследования должны будут покрывать. [ 26 ] Также отмечается, что африканские виды выдр -эрэт -энфиодон и Sivaonyx всегда были обнаружены на участках в связи с постоянными водоемами, в отличие от верхних слоев латолила в Лаэтоли , Танзании, в которой не хватало такой особенности, задавая вопрос на все. возможно, наземного образа жизни африканских видов Enhydriodon и Sivaonyx . [ 27 ]
E. Hendeyi был проанализирован на основе индекса устойчивости к бедренной кости (FRI) и индекса бедренного эпикондилара (FEI), в котором его значение Fri сравнимо с вымершим S. beyi , Enhydrhitherium и Sathericium (последние два аналогичны крупному морская выдца и река-выдца ( Pteronura brasiliensis ) соответственно и имеют большие значения в бедренных индексах, чем большинство других существующих выдр), в то время как его значение FEI аналогична существующей африканской выдре и выдр азиатской гигантская . Поскольку и африканская выдра, и азиатская выдр из малой когти, как правило, менее связаны с водоемами по сравнению с другими существующими выдрами, предполагается, что Э. Хендий и С. Бейи были полуакватическими локомоторами, у которых были более низкие ассоциации с водой, чем водные локомоторы энхидритра и Самиум , хотя С. Бейи, как говорили, был более наземным, чем Э. Хендей . Между тем, самые низкие значения соответствуют E. dikikae , что имеет аналогичные значения с наземным полуфоссористом (Адаптируется к копанию и жизни несколько под землей), такие как американские барсука и полосатый скунс , тем самым укрепляя гипотезу о том, что E. dikikae была более обобщенным наземным мускулом, похожим на S. beyi . [ 13 ]
С общим отсутствием консенсуса в отношении образа жизни рассматриваемых африканских энфидриононов , исследование 2022 года о E. Omoensis измеряло стабильные соотношения изотопов углерода и кислорода видов Enhydriodon по сравнению с существующими наземными млекопитающими, такими как Felids , Hyainids и Bovids вместе с полуакватическими млекопитающими, гиенидами и бовидами вместе с Млекопитающие, такие как бегемоты . Авторы объяснили, что использование изотопных соотношений кислорода, или Δ18O , может использоваться для понимания зависимости таксона от воды, в которой существующие водные и полуакированные таксоны, которые включают речные и морские выдры, имеют значительно более низкие изотопные отклонения кислорода по сравнению с земляными карниворанами. Исследователи, которые изучали E. Omoensis, обнаружили, что его значения зубной эмали Δ18O имели стандартное отклонение 2,7%, выходящее за пределы стандартных отклонений Δ18O морской выдры, и североамериканская река ( Lontra Canadensis ), которые были зарегистрированы 0,6 % и 0,3% -0,9% соответственно. Стандартное отклонение Enhydriodon Omo в большей степени выравнивается в диапазоне существующих наземных хищников, таких как Hyaenids, что предполагает, что это E. Omoensis был не таким полуакватическим, как изначально. Результаты исследования противоречат предположению 2008 года о том, что виды Enhydriodon Omo были водными. [ 14 ]
Grohe et al. Первоначально считалось, что диета Enhydriodon могла быть эллиптикой Oysteria Etheria , которая присутствовала на континенте в то же время. Основываясь на исследованиях с использованием стабильных изотопов углерода, диета чистых устриц приведет к значению эмали Δ13C -11,3%. Диета E. omoensis , однако, не была основана исключительно на Etheria , поскольку его значения углерода с минимально максимумом (от -9,7% до -4,7%) на ~ 2-7% более положительно, чем ожидаемое значение чистой устричной диеты. Его значения эмали Δ13C попадают в диапазон смешанных кормушек C 3 - C 4 , лишь частично попадающих в диапазон рационов водных кормушек растений C 3 , таких как рыба, черепахи или двустворчатые моллюски. Однако стандартное отклонение Δ13c Enhydriodon Omo Enhydriodon выходит за рамки изученных существующих популяций выдры пресной воды. Вместо этого считается, что E. Omoensis потреблял наземную добычу с диетой C 4 , по крайней мере, полурегулярно посредством охоты и/или удаления. Большой зубной зуб Bunodont из видов предполагает дурофажные способности, которые позволяют ему питаться на костях, включая кости, потенциально аналогично HEANAS или Mustelids. [ 14 ]
Палеоэкология
[ редактировать ]Пакистан и Индия
[ редактировать ]
E. Falconeri и E. Sivalensis , в то время как оба вида Enhydriodon , которые присутствовали на холмах Сивалик в Индии и Пакистане в течение периода неогена , не сосуществовали для тех же эпох, основанных на их появлении депозитов. Останки E. Falconeri присутствовали на формировании Нагри ( Дхок Милан и Сэти Нагри , Пакистан ) и формирование Дхока Патана (Dhok Pathan и Hasnot , Pakistan), оба образования датируются средним сиваликам, представляющим поздний миоцен. Этот вид также присутствовал в формировании Татрота ( Татрот , Индия) верхних сиваликов из раннего или среднего плиоцена . В формациях Нагри и Дхока Патана было показано, что Э. Фалконери существовал с несколькими архаичными плотоядными семействами млекопитающих, которые вымерли перед плиоценом, такими как HyainaiLourine Hyaenodonts и амфичиниды . Ранние виды выдры также существовали с различными вымершими плотоядными членами существующих семейств во время позднего миоцена, представляющих другие мустики , урсиды, Фелидс ( Felines и Махародонтины ), Гиниды ( Percrocutinae Hyeenids ictitheres и Hyaenines), Viverrids и Herpestids . Предполагается, что вымирание амфичинидов и перккрокутидов оставили пустые хищные ниши, которые быстро были заполнены другими родами Hyaenid, которые стали высоко диверсифицированными и сосуществовали с Felids в субконтиненте. [ 28 ]
Другие вымершие члены существующих и вымерших семей млекопитающих были обнаружены в формации Нагри и, таким образом, существовали с E. falconeri, включая бычьи, жираффиды , антракотеры , трагулиды , суиды , гиппарионинов эквиды , риноцеротиды , шаликотеры , гоминиды , гоминиды и спалакиды . [ 29 ] Вымерший рептилийский вид Гариал , Гавиалис Льюиси (?), Сообщается из формирования Дхока Патана в Пакистане и плиоцена в возрасте. [ 30 ] Роды млекопитающих, которые были обнаружены в формировании Dhok Pathan, как правило, согласуются с родами млекопитающих, обнаруженных в формации Нагри, но также включают в себя другие бычьи, жирафиды, шэвки швейцы , антракотеры, сауиды, гиппарионины, риноцеротиды, «тетратратоподоры», Cercopithecids и Hystrids , Полем [ 31 ] [ 32 ] [ 33 ] [ 34 ]

когда -то покрывающих большую часть индийского субконтинента и были заменены луга вечнозеленые до лиственных тропических лесов , Переход от среднего миоцена к позднему миоцене отражал период, когда Полем [ 35 ] Переход от поймы Нагри на поймы Дхока Патана предполагает меньше истощения в речной системе последнего по сравнению с первыми, и меньшие реки Дхока Патана имеют больше сезонного потока, чем раньше. Это отражает общую тенденцию позднего миоценового климата, что приводит к увеличению сезонности, что приводит к большим оборотам фауны. Более сухой и более сезонный климат наряду с речными изменениями, постепенно приводимыми к более крупным, открытым лесам, преимущественно состоящим из растений C 4 возле рек миоценовых рек , в то время как сообщества исключительно или преимущественно состоящие из C 3 растений значительно уменьшились и в конечном итоге исчезли на 7,0 мА вместе с C 3 кормушки, которые больше зависели от закрытой растительности. Эти изменения произошли вскоре после прибытия гиппарионинов и заметного снижения в группах млекопитающих в индийском субконтиненте, таких как вымирание гоминидного сивапитека и Deinothere deinotherium в результате фрагментации закрытых средств обитания в пользу открытых средств обитания, которые устраняют еду для c 3 браузера и бетоядные. [ 36 ] [ 37 ] [ 38 ]

Стоимость карниворана ископаемых отчетов о формировании Татрота в Индии мало, но среди вымерших членов, которые существовали с E. Falconeri в плиоцене, были другие лютрины, махародонтины и гиэениды. [ 28 ] Травоядные млекопитающие, обнаруженные на формировании Татрота на плато Потвара, содержат очень разнообразные собрания бычьи, но также включают в себя шэки, суиды, слоны , стегодонтиды , гиппарионины, антракотеры, гиппопотамиды, жираффиды и траглиды. [ 40 ] [ 41 ] [ 42 ] Crocodilians Crocodylus и Rhamphosuchus , пеликан Пелеканус , черепахи ( батагур , геоклеми , Hardella и pangshura ) и пресноводный краб -акантопотамон , по крайней мере, из формаций Tatrot или Pinjor, также указывают на активную среду пресноводного вода, что E. Falconeri , а затем и E. Sivalensis присутствовали. [ 30 ] [ 43 ] [ 44 ] [ 45 ]
Среди карнаворских таксонов Enhydriodon является самым продолжительным родообразным родом, когда-либо существовавшим в рамках сигаликов индийского субконтинента, идентифицированного из формаций Нагри-пинжор. Тем не менее, виды, идентифицированные в рамках формирования PINJOR эпох Plio-Pleistocene, являются E. sivalensis , который предполагает, что E. falconeri после долгого времени относительного успеха в конечном итоге мог пройти через анагенез плиоценом. Другие роды карнаворана, которые были обнаружены в формировании Pinjor, включали в себя вновь прибывшие каниды , а также мустлиды, урсиды, фелиды (макародонтины, пантерины и кошачьи), гиэениды и виверриды. [ 28 ] Другие роды млекопитающих, обнаруженные в формировании PINJOR, включают гоминиды, Cercopithecids, грызуны из различных семейств, хобоскейцы, лошади племени Equini , носороги, суиды, швейки, жираффиды и бычьи. [ 46 ]
Эфиопия
[ редактировать ]
E. Dikikae и E. Omoensis были большими, проведенные Luettrine в разных местах Эфиопии современной E. Dikikae окаменелости были обнаружены в двух нижних последовательностях формирования хадара низкой долины Awash , Эфиопия, что указывает на то, что ее фокус варьируется от 4 мА до 3,2 млн. Лет. Окаменелости E. omoensis были расположены в формировании USNO и формировании Shungura нижней долины Омо в Эфиопии, фокус в диапазоне от 3,44 млн. Лет до 2,53 млн. Лет. E. Dikikae был назван в честь базального члена дикика, в то время как E. Omoensis имел свой название дизайна из нижней долины Омо. [ 15 ] [ 14 ]
Существует четыре члена композитной последовательности Дикика в рамках формирования плиоценового хадара, от основания на вершину: базальные, Сиди Хакома, Денен Дора и члены Када Хадар. Все вместе, они датируются ок. 3,5-2,9 млн. Лет и наиболее известны за многочисленные останки Австралопитека Afarensis . [ 47 ] E. Dikikae окаменелости известны из членов базальных и Sigi Hakoma Formation и Sigi Hakoma и неизвестны в других двух лучших членах. [ 15 ]
Основываясь на методах определения палеооплаты, таких как экоморфологический анализ, зубной микровол из бычьей и изотопы углерода и кислорода эмали, базальный член (BM) имеет наибольшее содержание боидов и суидов в образовании Хадра, что предполагает, что окружающая среда, которых они, которых они Были присутствовали, возможно, древесные луга, а также речные леса. Aepycerotini предпочтения были распространены внутри участника, подходящего для экотональной племени среды обитания между лугами и лесами. [ 48 ]
Судьба Sidi Hakoma 1 (SH-1), в диапазоне от ~ 3,45 до 3,35 млн. Лет, имела аналогичную фауну и, следовательно, аналогичные места обитания с другими членами в формировании хадара, но также, вероятно, включали водно-болотные угодья в определенных регионах. Таксоны, такие как вид в племени цефалофини и пять видов приматов, были обнаружены у члена, что еще больше указывает на большой речный лес, в основном лесные массивы в окрестностях. Aepyceros был самым распространенным бовидом, и у SH-1 была самая низкая доля выпавших боид в любом подчиненном формации хадара. Растительность SH-1 могла бы очень напоминать тех, кто в Гвинее или суданском саваннах в центральной африке , которые взаимодействуют с тропическим лесом , который создает мозаики среды обитания пастбищ, лесных массивов и некоторых лесных ремней. Сборка остракода членов базальной и сиди-хакомы указывает на источники ввода пресной воды, в которых их раковины также указывают только на трехмесячный сухой сезон, характерный для центральной африканской саванны. Единственный сухой сезон, указывающий на девятимесячный сезон дождей, является отличительным фактором члена Sidi Hakoma из современного климата в Восточной Африке, который имеет бимодальный формат сухого сезона (два сухого сезона), а не один. Sidi Hakoma Summember 2 (SH-2) похожа на SH-1 и, как полагают, была связана с лесами с некоторыми травянистыми равнинами, из которых из которых Aepycerotini был наиболее распространенным. [ 48 ]
Sidi Hakoma Samember 3 (SH-3) указывает на наличие лесных массивов и пастбищ с большим количеством водно-болотных угодий на берегу озера по сравнению с предыдущими субмаристами, с повышенным присутствием рюкшины и самым высоким содержанием бовидов трагелафина , которые указывают на либо больше закрытых мест обитания или водно-боевых угодий. Полем Он также содержит самые большие микромамные сборки существующих муридских родов, таких как существующие Acomys , Golunda и oenomys и вымершие указания , из которых Golunda в настоящее время вымерла в Африке. Sidi Hakoma Summember 4 указывает на среды обитания водно -болотных угодий, которые окружают озера в более сухой среде. Дальнейшее увеличение Reduncinae Bovids и снижение Alcelaphin Bovids указывают на указанную среду озера и окружающие водно -болотные угодья. Данные об изобилии Bovid предполагают аналогичные количества деревьев для SH-3 и SH-4, причем разница в том, что последний был немного более сухим, чем первое. [ 48 ]

Формирование хадара представляет собой много окаменелостей Australopithecus afarensis , особенно частичный скелет, известный как « Люси ». Совокупный промежуток времени вида составляет не менее 0,7 млн. Лет, от 3,7 до 3,0 мА. [ 49 ] Формирование хадара также известно своим представлением о большом разнообразии видов бовидов, которые представляли большинство основных племен в Африке. Бовидские племена, которые были обнаружены в формировании, включали аепикоротини, алькелафини, антилопини , Бовини , Каприни , Цефалофини, Гиппотрагини , Неотрагини , Редунсини и Трагелафини. Другие группы, представленные вымершими видами существующих или вымерших родов, включают жираффиды, бегемоты, суиды, каниды, фелиды (макародонтины были наиболее распространенными), гианиды, другие мускулиды, виверриды, носороги, равные, катаррины , деинотеры и элементы -элементы. также найдено в местности. Небольшие группы млекопитающих включают летучие мыши , лепориды, дикобразы в старом мире, грызуны, спалакиды , белки и аардварки . [ 48 ] [ 47 ]
Другие плиоценовые образования в Эфиопии показывают сходные тенденции большого разнообразия в семействе Бовиде из его множественных племен, а также суиды, гиппопатамиды, сермопитециды, гоминиды и равные, как правило, те же роды, что и формация Хад. Большинство травоядных животных, присутствующих в формировании Shungura, показывают либо согласованные диеты C 4 , либо обычно смещались с смешанных диет C 3 -C 4 на диеты C 4 , как показано из изменений в зубном виде членом формирования. Эти тенденции предполагают, что африканские травоядные животные в плиоцене все чаще переходили к травоядным в C 4 , в отличие от просмотра и смешанного кормления в результате растущего доминирования пастбищ C 4 в Африке. Однако было несколько исключений, поскольку Giraffidae и Deinotheriidae были последовательно C 3 браузерами в форме, в то время как племена Bovid Aepycerotini и Tragelaphini были преимущественно смешанными питателями с небольшими изменениями в рационе. [ 50 ] [ 51 ] [ 52 ] Остатки ископаемой рыбы также известны из формирования Шунгуры, а именно в родах Polypterus , Sindacharax , Synodontis , Auchenoglanis и Lates . [ 53 ]
Примечания
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а беременный в Хью Фалконер (1868). Палеонтологические мемуары и ноты покойного Хью Фалконера: Фауна Антиква Сиваленсис . Великобритания: Р. Хардвик. ISBN 112-0-015839 .
- ^ Jump up to: а беременный в Верма, Британская Колумбия; Гупта, С.С. (1992). «Enhydriodon Sivalensis, гигантская ископаемая выдр из формирования сакети (верхний плиоцень), Siwalik Group, Sirmur District, Himachal Pradesh». Журнал Палеонтологического общества Индии . 37 : 31–36.
- ^ Cautley, Proby Thomas; Falconer, Hugh (1835). «Синопсис родов ископаемых и видов из верхних месторождений третичных слоев сивальских холмов, в сборе авторов». Журнал Азиатского общества Бенгалии . 4 (12).
- ^ Jump up to: а беременный Гай Э. Пилигрим (1932). Ископаемое плотояд Индии . Индия: Палеонтология указывает. стр. 80–88.
- ^ Jump up to: а беременный в Виллемен, Джерард Ф. (1992). «Пересмотр плиоцена и четвертичного латрина из Европы». Scripta Geologica . 101 : 89–94.
- ^ Repenning, Charles A. (1976). «Enhydra и Enhydriodon с Тихоокеанского побережья Северной Америки». Журнал исследований геологической службы США . 4 (3): 305–315.
- ^ Хюрзелер, Йоханнес; Engesser, Burkart (1976). «Фауны неогенных млекопитающих бассейна Бакцинелло (Гроссет, Италия)» Образует учетные записи Академии наук Парижа (на французском языке). 283 : 333–336.
- ^ Jump up to: а беременный в дюймовый и фон Моралес, Хорхе; Пикфорд, Мартин (2005). «Гигантский Bunodont Latrinae из мио-плиоцена Кении и Уганды» . Геологические исследования (на испанском). 61 (3): 233–245. Doi : 10.3989/egeol.05613-666 .
- ^ Пикфорд, Мартин; Сория, боль; Моралес, Хорхе (2005). «Престивонные из покойного миоцена и базальный плиоцень холмов Тугена, Кения». Revista de la Sociedad Geológica de España . 18 (1–2): 39–61.
- ^ Jump up to: а беременный в дюймовый и фон глин час Пикфорд, Мартин (2007). «Пересмотр мио-плиоценовых выдких млекопитающих индийского субконтинента индийского субконтинента». Estudios Geológicos (по -испански). 63 (2): 108–124. doi : 10.3989/egeol.07631192 .
- ^ Jump up to: а беременный в дюймовый и фон Ван, Синьон; Грохе, Камилла; Су, Дениз Ф.; Белый, Стюарт С.; Ji, xueping; Келли, Джей; Jablonski, Nina G.; Дэн, Тао; Ты, Юшан; Ян, Синь (2017). «Новая выдра гигантского размера, Siamogale Melilutra sp. Nov. (Lutrinae: Mustelidae: Carnivora), из последнего места миоценового Шуитунгба на северо-востоке Юньнани, юго-западного Китая и полной филогенией лютринов» . Журнал систематической палеонтологии . 16 (1): 1–24. Bibcode : 2018jspal..16 ... 39w . doi : 10.1080/14772019.2016.1267666 . S2CID 58892181 .
- ^ Верделин, Ларс (2003). «Преступники из гоминидного участка Канапоя, бассейн Туркана, северная Кения » Вклад в науку 498 : 115–1 Doi : 10.5962/ p.2 S2CID 82943977
- ^ Jump up to: а беременный в дюймовый и Валенсиано, Альберто; Говендер, Ромала (2020). «Новое понимание гигантских Mustelids (Mammalia, Carnivora, Mustelidae) от Langebaanweg Fossil Site (парк ископаемого западного побережья, Южная Африка, ранний плиоцен)» . ПЕРЕЙ . 8 : E9221. doi : 10.7717/peerj.9221 . PMC 7271888 . PMID 32547866 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k Грохе, Камилла; Уно, Кевин; Boisserie, Жан-Рен (2022). Plio-Pleiste (Cenivora) из города . Кошки Рендус бледный (по -французски). 30 (30): 684–6 doi : 10.5852/cr- palle S2CID 2521066648 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k Гераадс, Денис; Германия, Zereseseay; Бобе, Рене; Рид, Денне (2011). "Ornivoa: ноябрь. А 31 (2): 447–4 Bibcode : 201jvpal . doi : 10.1080/ 02724634.5011.503356 S2CID 84797296 .
- ^ Валенсиано, Альберто; Моралес, Хорхе; Пикфорд, Мартин (2022). «Лутра» Либика из мио/плиоцена Вади Натрун (Египет): новое предложение о систематической . 2-й международный симпозиум по палеонтологии позвоночных 20-го 22-го сентября 2022 года. С. 31–32.
- ^ Jump up to: а беременный Каргопулос, Николаос; Валенсиано, Альберто; Kampouridis, Panagiotis; Лехнер, Томас; Böhme, Madelaine (2021). «Новые ранние поздние миоценовые виды Vishnuonyx (Carnivora, Lutrinae) из гоминидного населенного пункта Хаммершмии, Бавария, Германия». Журнал палеонтологии позвоночных . 41 (3). Bibcode : 2021jvpal..41e8858k . doi : 10.1080/02724634.2021.1948858 . S2CID 240538139 .
- ^ "Enhydrithritherium terraenovae" . Флоридский музей естественной истории. 30 марта 2017 года . Получено 10 октября 2022 года .
- ^ Константино, Пол Дж.; Ли, Джеймс Дж. В.; Моррис, Дилан Дж.; Лукас, Питер В.; Хартстоун-Роуз, Адам; Ли, Ва-Киат; Домини, Натаниэль Дж.; Каннингем, Эндрю; Вагнер, Марк; Lawn, Брайан Р. (2011). «Адаптация к твердому кормлению в морских выдрах и гомининах». Журнал человеческой эволюции . 61 (1): 89–96. Bibcode : 2011jhume..61 ... 89c . doi : 10.1016/j.jhevol.2011.02.009 . PMID 21474163 .
- ^ Raghavan, P.; Пикфорд, М.; Patnaik, R.; Гаятри П. (2007). «Первая ископаемая выдра с мелкой когтикой, Amblonyx, с заметками о некоторых образцах Лутра, от верхнего сивалика, Индия» . Estudios Geológicos (по -испански). 63 (2): 144–145. doi : 10.3989/egeol.07632200 .
- ^ Koufos, George D. (2011). «Миоценовая плотояродная сборка Греции» . Геологические исследования (на испанском). 67 (2): 296–297. Doi : 10.3989/egeol.40560.190 .
- ^ Альберто Валенсиано Вакеро; Ромала Говендер (21 июня 2020 года). «Гигантские росомахи, выдры размером с волков: окаменелости дают свежую информацию о прошлом» . Разговор . Получено 27 ноября 2022 года .
- ^ Jump up to: а беременный Льюис, Маргарет Э. (2008). «Бедренная бедренная кость вымерших выдр из Бундонта в Африке (Carnivora, Mustelidae, Lutrinae)». Comptes rendus palevol (по -французски). 7 (8): 607–627. Bibcode : 2008crpal ... 7..607l . Doi : 10.1016/j.crpv.2008.09.010 .
- ^ Стюарт, Кэтлин М. (2001). «Пресноводная рыба неогена Африки (миоцен -платеистоцена): систематика и биогеография». Рыба и рыболовство . 2 (3): 177–230. Bibcode : 2001aqff .... 2..177s . doi : 10.1046/j.1467-2960.2001.00052.x .
- ^ Пейгне, Стефан; Де Бонис, Луи; ЛИМИЙС, Андосси; Маккей, Хассан Тайссо; Виньо, Патрик; Брюнет, Мишель (2008). «Поздний миоценовый хищник от Чада: Lutrinae (Mistelidae)» . Зоологический журнал Линневого общества . 152 (4): 793–846. Doi : 10.1111/j.1096-3642.2008 00377.x .
- ^ Верделин, Ларс; Мантити, Фредрик Кьяло (2012). «Плотно -килограмма с участка Канапоя гоминина, северная Кения» Журнал африканских наук о земле 64 : 1–8 Bibcode : 2012jaffes..64 .... 1W Doi : 10.1016/ j.jafrearsci.2011.11.0
- ^ Верделин, Ларс; Льюис, Маргарет Э. (2020). «Контекстуальный обзор плотива Канапоя» . Журнал человеческой эволюции . 140 : 102334. Bibcode : 2020jhume.14002334W . doi : 10.1016/j.jhevol.2017.05.001 . PMID 28625408 . S2CID 23285088 .
- ^ Jump up to: а беременный в Jasinski, Steven E.; Аббас, Сайед Гьюр; Махмуд, Халид; Бабар, Мухаммед Адиб; Хан, Мухаммед Акбар (2022). «Новые экземпляры Carnivoran (Mammalia: Carnivora) из сигаликов Пакистана и Индии, а также их фауны и эволюционные последствия». Историческая биология: международный журнал палеобиологии . 35 (11): 1–36. doi : 10.1080/08912963.2022.2138376 . S2CID 253346978 .
- ^ Хан, Мухаммед Акбар; Ахтар, Мухаммед; Икрам, Таснем. «Истинные копылы из местности типа Нагри (поздний миоцена), Северный Пакистан». Журнал наук о животных и растениях . 22 : 1–59.
- ^ Jump up to: а беременный Мартин, Жери (2018). «Таксономическое содержание рода gavialis из холмов Сивалика Индии и Пакистана» (PDF) . Документы по палеонтологии . 5 (3): 483–497. doi : 10.1002/spp2.1247 . S2CID 134966832 .
- ^ Хан, Мухаммед Акбар; Ахтар, Мухаммед; Илиопулос, Джордж (2012). «Tragguguds (Artiododactyla, Ruminantia, Tragulidae) из среднего сигаликов Hasnot (поздний миоцена), Пакистан» Rivist italiana di palontology e stragigia 118 (2): 325–3 Doi : 10.13130/ 2039-4942/ 6
- ^ Гаффар, Абдул; Ахтар, Мэнммд; Бабар, Мамадмад Адев; ROOHI, Газала (2018). Полем Revista brasilera de paleontolog 21 (2): 132–140. doi : 10.4072/rp . S2CID 59060740 .
- ^ Отец, Гюр; Бабар, Мухаммед Адеб; Хан, Мухаммед Акбар; un nisa, badar (2021). «Новый хоботокский материал из группы Пакистана Сивалика с замечаниями о некоторых видах » Палеобио 38 (1): 1–1 Doi : 10.5070/ p9381054113 S2CID 237772570
- ^ Бабар, Мухаммед Адеб; Aftab, аренда; Хан, Мухаммед Акбар; Аббас, Гюр (2021). «Терракондон и Сивачорус (Суйдае) из скрытого патана и не имеют площади Потвара, Северный Пакистан» Аравийский журнал о геологиях 14 (2473): 1–6 Bibcode : 2021Arjg ... 14.2473b Doi : 10.1007/s12517-021-08684- x 244120161S2CID
- ^ Ван, Синьон; Флинн, Лоуренс; Fortelius, Mikael (2013). «Глава 17: Индийская биостратиграфия млекопитающих индийского неогена: обзор». В Ван, Сяоминг; Флинн, Лоуренс Дж.; Fortelius, Mikael (Eds.). Ископаемые млекопитающие Азии: неогеновая биостратиграфия и хронология . Издательство Колумбийского университета. С. 423–444. doi : 10.7312/wang15012-017 . ISBN 9780231520829 .
- ^ Барри, Джон С.; Morgan, Michèle E.; Флинн, Лоуренс Дж.; Pilbeam, Дэвид; Беренсмейер, Анна К.; Раза, С. Махмуд; А. Хан, Имран; Баджли, Кэтрин; Хикс, Джейсон; Келли, Джей (2002). «Изменения фауны и окружающей среды в покойном миоценовом сигаликах северного Пакистана». Палеобиология . 28 (2): 1–71. doi : 10.1666/0094-8373 (2002) 28 [1: faecit] 2.0.co; 2 . S2CID 18408360 .
- ^ Нельсон, Шерри В. (2007). «Изотопные реконструкции среды обитания изменяют исчезновение Сивапитека, миоценового гомеиноида, в группе Сивалика в Пакистане» (PDF) . Палеогеография, палеоклиматология, палеоэкология . 243 (1–2): 204–222. Bibcode : 2007ppp ... 243..204n . doi : 10.1016/j.palaeo.2006.07.017 .
- ^ Патнаик, Раджив; Сингх, Нингтуджам Премедит; Пол, Дебаджёти; Сукумар, Раман (2019). «Диетические и среду обитания сдвигаются по отношению к климату неогеновых квалифицированных хобоскосков и связанных с ними млекопитающих индийского субконтинента». Кватернарные науки обзоры . 224 : 105968. BIBCODE : 2019QSRV..22405968P . doi : 10.1016/j.quascirev.2019.105968 . S2CID 210307849 .
- ^ Ма, Цзяо; Ван, Юань; Джин, Чанцжу; Ху, Яову; Bocherens, Hervé (2019). «Экологическая гибкость и дифференциальная выживаемость плейстоценового стегодона Orientalis и Elephas Maximus в материковой части Юго -Восточной Азии, выявленные с помощью стабильного анализа изотопа (C, O)». Кватернарные науки обзоры . 212 : 33–44. Bibcode : 2019qsrv..212 ... 33M . doi : 10.1016/j.quascirev.2019.03.021 . S2CID 135056116 .
- ^ Джукар, Совместный Мухеш; Солнце, мальчик; Нанда, Авинаш С.; Бернор, Рэймонд Л. (2019). «Первый происхождение Eurygnathipus van Hoepen, 1930 (Mammalia, Perissodactyla, Equidae) за пределами Африки и ее биогеографическое значение». Бюллетень итальянского палеонтологического общества . 58 (2): 171–179. Doi : 10.4435/bspi.2019.13 .
- ^ Хан, зрелый Акбар; Джамиль, Дейчя; Махмуд, Чалид; Бабар, Мухаммед Адиб (2014). "Sume News of Chervids. Биология . 60 (2): 225–2
- ^ Кумар, Суреш (2022). «Новые окаменелости Sivatherium Giganteum (Giraffidae, Mammalia) из верхних сиваликов индийского субконтинента» . Acta Palaeontologica Romaniae . 18 (2): 85–92. doi : 10.35463/j.apr.2022.02.04 . S2CID 248233024 .
- ^ Томас А., Стидхэм; Кришан, Кевал; Сингх, Бахадур; Гош, Абхик; Патнаик, Раджив (2014). «Пеликан Tarsometatarsus (Aves: Pelecanidae) из последних плиоценовых сиваликов Индии» . Plos один . 9 (11): 1–8. Bibcode : 2014ploso ... 9K1210S . doi : 10.1371/journal.pone.0111210 . PMC 4218845 . PMID 25365300 .
- ^ Гарбин, Рафаэлла С.; Баньядади, будьте осторожны; Джойс, Уолтер Г. (2020). «Пумомический пересмотр геоэмического пересмотра из Sik известного возраста Индии и Пакистана » : 1–6 doi : 10.5852/ejt . S2CID 229387265 .
- ^ Клаус, Себастьян; Сингх, Бахадур; Хартманн, Лукас; Кришан, Кевал; Гош, Абхик; Патнаик, Раджив (2017). «Ископая пресноводная краба из плаоцена татрота (Siwalik Group) в северной Индии (Crustacea, Brachyura, Potamidae)». Палеоворд . 26 (3): 566–571. doi : 10.1016/j.palwor.2016.08.003 .
- ^ Нанда, Авинаш С.; Сегал, Рамеш К.; Chauhan, Parth R. (2018). «Фауны Сивалик-эй из Гималайского бассейна из Южной Азии». Журнал азиатских наук о Земле . 162 : 54–68. Bibcode : 2018jaesc.162 ... 54n . doi : 10.1016/j.jseaes.2017.10.035 . S2CID 134929306 .
- ^ Jump up to: а беременный Герадс, Денис; Алемсегед, Zeresenay; Бобе, Рене; Ред, Денне (2015). «Плиоценовая плотная (млекопитающая) из формации хадара в Дикике, Нижняя долина Аваш, Эфиопия» Журнал африканских наук о земле 107 : 28–3 Bibcode : 2015Jefes.107 ... 28G Doi : 10.1016/ j.jafrearsci.2015.03.0
- ^ Jump up to: а беременный в дюймовый Камписано, Кристофер Дж.; Роуэн, Джон; Рид, Кэй Э. (2022). «Глава 18: Формирование хадара, региональное государство из Афар, Эфиопия: геология, фауна и палеоэкологические реконструкции». В Рейнольдсе, Салли С.; Бобе, Рене (ред.). Африканская палеоэкология и эволюция человека . Издательство Кембриджского университета. С. 214–228. doi : 10.1017/9781139696470.018 .
- ^ Германия, Zereseseay; Винн, Джонатан Дж.; Cembel, William H.; Рид, Денне; Гераадс, Денис; Бобе, Рене (2005). "Геологический контекст" человека Журнал эволюции 49 (4): 499–514. Bibcode : 2005jsho . doi : 10.1016/jhevol . PMID 16026815 .
- ^ Бобе, Рене (2011). «Ископаемые млекопитающие и палеоража в бассейне Омо-Туркана». Эволюционная антропология . 20 (6): 254–263. doi : 10.1002/evan.20330 . PMID 22170694 . S2CID 205826374 .
- ^ Негаш, Enquye W.; Алемсегед, Zeresenay; Бобе, Рене; Грин, Фредерик; Sponheimer, Matt; Г. Винн, Джонатан (2020). «Разрушенные тенденции в травоядных животных из формирования Шунгуры, юго -западной Эфиопии » Труды Национальной академии наук 117 (36): 21921–2 Bibcode : 2020pnas..11721921NN Doi : 10.1073/ pnas.2006982117 7486712PMC 32839326PMID
- ^ Биби, Файсал; Суйрон, Антуан; Bocherens, Hervé; Уно, Кевин; Boisserie, Ean-Renaud (2013). «Экологические изменения в нижней долине Омо около 2,8 млн. Лет » Биологические письма 9 (1): 1–4 Doi : 10.1098/ rsbl.2012.0 3565503PMC 23234862PMID 10791621S2CID
- ^ Стюарт, Кэтлин М.; Мюррей, Элисон (2008). «Рыба остается от плито-плейстоновой формирования Shungura, бассейна реки Омо, Эфиопия» Геобис 41 (2): 283–2 Bibcode : 2008 Gobob..41..283s Doi : 10.1016/ j.geobous.2007.06.0







- Выдры
- Доисторические мустиды
- Миоценовые мустиды
- Ископаемые таксоны описаны в 1868 году
- Миоценные млекопитающие Африки
- Плиоценовые млекопитающие Африки
- Плейстоценовые млекопитающие Африки
- Миоценовые млекопитающие Азии
- Плиоценовые млекопитающие Азии
- Таксоны, названные Хью Фальконером
- Доисторические карниворские роды