Длиннопалая саламандра
| Длиннопалая саламандра | |
|---|---|

| |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Амфибия |
| Заказ: | Уродела |
| Семья: | Амбистоматиды |
| Род: | Амбистома |
| Разновидность: | А. макродактилум
|
| Биномиальное имя | |
| Амбистома макродактилум Бэрд , 1950 год.
| |
| Подвиды | |
|
Являюсь. Колумбианум | |
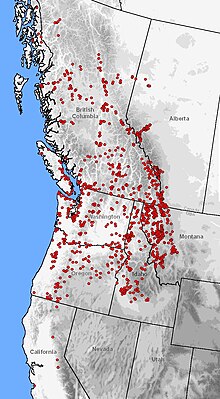
| |
| Распространение A. macrodactylum (красные точки) на западе Северной Америки. | |
| Синонимы [ 2 ] | |
| |

Длиннопалая саламандра ( Ambystoma macrodactylum ) — кротовая саламандра из семейства Ambystomatidae . [ 2 ] Этот вид, обычно достигающий в зрелом возрасте 4,1–8,9 см (1,6–3,5 дюйма) в длину, характеризуется пестрой черной, коричневой и желтой пигментацией, а также длинным внешним четвертым пальцем на задних конечностях. Анализ записей окаменелостей , генетики и биогеографии позволяет предположить, что A. macrodactylum и A. Laterale произошли от общего предка, который получил доступ к западным Кордильерам с утратой срединного континентального морского пути к палеоцену .
Распространение длиннопалой саламандры в основном происходит на северо-западе Тихого океана , с высотным диапазоном до 2800 м (9200 футов). Он обитает в различных средах обитания, включая тропические леса умеренного пояса , хвойные леса , горные прибрежные зоны, полынные равнины, красные еловые леса, полузасушливые полынные равнины, равнины читграсса и альпийские луга вдоль скалистых берегов горных озер. он обитает в медленных ручьях, прудах и озерах Во время водной фазы размножения . Длиннопалая саламандра впадает в спячку в холодные зимние месяцы, выживая за счет запасов энергии, хранящихся в коже и хвосте.
Пять подвидов имеют разную генетическую и экологическую историю, фенотипически выраженную в различных цветах и рисунках кожи. Хотя длиннопалая саламандра классифицируется МСОП как вид, вызывающий беспокойство наименьшее , многие формы освоения земель угрожают и отрицательно влияют на среду обитания саламандры.
Таксономия
[ редактировать ]Ambystoma macrodactylum — представитель семейства Ambystomatidae , также известного как кротовые саламандры. Ambystomatidae произошли примерно 81 миллион лет назад (поздний мел ) от родственного таксона Dicamptodontidae . [ 3 ] [ 4 ] [ 5 ] Ambystomatidae также являются членами подотряда Salamandroidea , в который входят все саламандры, способные к внутреннему оплодотворению. [ 6 ] Родственным видом A. macrodactylum является A. Laterale , распространенный в восточной части Северной Америки. Ambystomatidae на уровне вида Однако филогения является предварительной и требует дальнейшего тестирования. [ 7 ]
Систематика и биогеография
[ редактировать ]Эволюционное происхождение
[ редактировать ]Предковое происхождение этого вида происходит из восточной части Северной Америки , где видовое богатство амбистоматид самое высокое. [ 8 ] [ 9 ] Следующая биогеографическая интерпретация происхождения A. macrodactylum в западной части Северной Америки основана на описательном описании окаменелостей, генетики и биогеографии. [ 10 ] [ 11 ] длиннопалой саламандры Ближайшим родственным видом является A. Laterale , обитающий на северо-востоке Северной Америки. [ 4 ] [ 9 ] Ambystomatidae были изолированы к юго-востоку от среднеконтинентального или Западного внутреннего морского пути в меловой период (~ 145,5–66 млн лет назад). [ 9 ] [ 12 ] В то время как три других вида Ambystomatidae ( A. tigrinum , A.californiense и A.gracile длиннопалой саламандры ) имеют перекрывающиеся ареалы в западной части Северной Америки, ближайшим родственным видом является A. Laterale , обитающий на северо-востоке Северной Америки. [ 4 ] [ 9 ] Было высказано предположение, что A. macrodactylum произошел от A. Laterale после палеоцена (~ 66–55,8 млн лет назад), когда потеря Западного внутреннего морского пути открыла путь доступа для общего предка в Западные Кордильеры . [ 11 ] Когда-то обитавшие в горных регионах западной части Северной Америки, видам приходилось иметь дело с динамической пространственной и композиционной экологией, реагирующей на изменения высоты по мере роста гор и изменения климата. Например, на северо-западе Тихого океана в палеоцене стало прохладнее , проложив путь к лесам умеренного пояса , которые заменили более теплые тропические леса периода мелового . [ 13 ] Сценарий отделения A. macrodacylum и других западных видов умеренного пояса от их восточных аналогов предполагает поднятие Скалистых гор в конце олигоцена - миоцене . Орогения создала климатический барьер, удалив влагу из западного воздушного потока, и высушила центральную часть континента, от южной Альберты до Мексиканского залива. [ 10 ] [ 14 ]
Предки современных саламандр, вероятно, смогли расселиться и мигрировать в среду обитания Скалистых гор и прилегающих территорий к эоцену . Мезические леса возникли на западе Северной Америки в середине эоцена и достигли своего современного распространения к раннему плиоцену . Лесные долины умеренного пояса и горная среда этих периодов времени ( от палеогена до неогена ) могли обеспечить физико-географические и экологические особенности, подтверждающие аналоги современных местообитаний Ambystoma macrodactylum . [ 10 ] [ 11 ] [ 15 ] [ 16 ] Каскадный хребет поднялся в середине плиоцена и создал эффект дождевой тени , вызвав ксерификацию бассейна Колумбии , а также изменив диапазоны мезических экосистем умеренного пояса на возвышенностях. Подъем Каскадов, вызвавший ксерификацию бассейна Колумбии, является основной биогеографической особенностью западной части Северной Америки, которая разделила многие виды, включая A. macrodactylum , на прибрежные и внутренние линии. [ 11 ] [ 14 ] [ 16 ] [ 17 ]
Подвиды
[ редактировать ]Существует пять подвидов длиннопалой саламандры. [ 18 ] Подвиды различаются по географическому положению и узору на спинной полосе; [ 19 ] Дензел Фергюсон дает биогеографическое описание структуры и морфологии кожи; на основе этого анализа он ввел два новых подвида: A. macrodactylum columbianum и A. m. сигиллатум . [ 18 ] Ареалы подвидов проиллюстрированы в полевых справочниках Роберта Стеббина по амфибиям. [ 19 ]


Внешний вид (фенотипы)
[ редактировать ]Краткое изложение отличительных рисунков кожи и морфологических особенностей подвида включает: [ 18 ] [ 21 ]
- Являюсь. крест
- Оранжевая окраска спины хвоста, распадающаяся на пятна вдоль черного тела и на мелкие точки на голове, часто отсутствует перед глазами. По бокам имеются беловатые крапинки. Число реберных бороздок равно 13.
- Являюсь. Колумбианум
- Полоса на спине от желтого до коричневого цвета на черном теле, сплошные пятна или пятна вдоль тела, заканчивающиеся суженными пятнами с пятнистым рисунком, распределенным по голове. Белые крапинки по бокам и снизу остаются отдельными мелкими крапинками. Число сошковых зубов более 35.
- Являюсь. Краузей
- Полоса на спине от желтого до коричневого цвета, непрерывные пятна или пятна вдоль тела, заканчивающиеся расширенными пятнами с пятнистым рисунком, распределенным по голове. Белые крапинки по бокам и снизу остаются отдельными мелкими крапинками. Число сошковых зубов 32. Число реберных бороздок 12.
- Являюсь. запечатанный
- Полоса на спине от восково-желтой до коричневой, образующая пятнистые или неправильной формы пятна вдоль тела, оканчивающиеся точками или крапинками дорсального цвета на голове. Число сошковых зубов 44. Число реберных бороздок 13.
- Являюсь. макродактилия
- Цитрин, тусклый цитрин, с коричневой полосой на спине, которая размыта и непрерывна вдоль сероватого тела. Рисунок заканчивается размытыми крапинками полосатого цвета или отсутствует на голове и морде. Белые пятна по бокам иногда сливаются, образуя более крупные пятна. Число сошковых зубов 33, образующих выраженную поперечную дугу. Число реберных бороздок равно 13.
Биогеография и генетика
[ редактировать ]митохондриальной ДНК Анализ [ 11 ] определяет несколько разные диапазоны линий подвидов. [ 11 ] Генетический анализ, например, выявляет дополнительную картину глубокой дивергенции в восточной части ареала. Пространственное распределение популяций и генетика этого вида пространственно и исторически связаны между собой соединяющими горными и долинными системами умеренного пояса западной части Северной Америки. [ 11 ] [ 22 ] Преданность размножению длиннопалых саламандр ( филопатрия ) и другие миграционные модели поведения снижают скорость расселения между регионами, например, в горных бассейнах. Этот аспект их поведения ограничивает поток генов и увеличивает степень и скорость генетической дифференциации . Генетическая дифференциация между регионами у длиннопалой саламандры выше, чем у большинства других групп позвоночных . [ 23 ] Естественные разрывы в диапазоне расселения и миграции происходят там, где экосистемы переходят в более сухие ксерические низменности (например, климат прерий ) и в замерзшую или более суровую местность на экстремальных высотах (2200 метров (7200 футов)). [ 24 ]
- Являюсь. Колумбианум
- Генетические данные о «центральном» подвиде ( A.m. columbianum ) позволяют предположить, что он не распространяется на север, в Британскую Колумбию, а ограничивается горами Блу и Уоллова в центральных и северо-восточных частях Орегона. Популяции ограничены этими территориями каньоном реки Снейк (Айдахо) на востоке и низкими засушливыми или ксерическими землями в бассейне Мадраса на западе. [ 11 ]
- Являюсь. макродактилия
- Линия «прибрежного» или «западного» подвида ( A.m. macrodactylum ) простирается на север от северо-восточной Калифорнии, через хребет Кламат-Сискию , через долину Уилламетт , вдоль прибрежных горных хребтов, включая Каскадные горы, и продолжается на север через Британскую Колумбию. Колумбия и вплоть до Аляски. [ 11 ]
- Являюсь. крест
- ( Длиннопалая саламандра Санта-Крус A.m. croceum ) наиболее тесно связана с «прибрежным» или «западным» подвидом. Этот вывод представляет собой наиболее экономное биогеографическое объяснение с учетом ближайших популяций A. m. macrodactylum разделены примерно 300 км через дельту реки Сакраменто-Сан-Хоакин , Калифорния. [ 10 ] Изолированные эндемичные популяции занесены в список исчезающих подвидов. [ 20 ] Судя по биогеографии и калибровке молекулярных часов, этот подвид мог быть отделен от остальной части ареала начиная с миоцена, калибровка молекулярных часов оценивает разделение в 13,9 миллиона лет. [ 10 ]
- Являюсь. Краузей
- Ареал «восточного» подвида ( A.m. krausei ) распространен во внутренних горах, причем западная часть его ареала вторгается в низменные районы центрального внутреннего плато Вашингтона и Британской Колумбии, а восточная часть его ареала - Диапазон простирается через долины Скалистых гор в низменные предгорья и прерии Монтаны и Альберты. [ 25 ] [ 11 ]
- Являюсь. запечатанный
- Традиционный «южный» подвид ( A.m. sigillatum ) не имеет митохондриальной генетической идентичности. [ 11 ] Этот подвид был идентифицирован Фергюсоном как интерградный с A. m. columbianum на юге центральной части Орегона. [ 18 ]
Томпсон и Рассел обнаружили еще одну эволюционную линию, которая берет свое начало в ограниченной ледниковой зоне гор Салмон-Ривер , штат Айдахо. [ 11 ] С наступлением голоценового межледниковья , примерно 10 000 лет назад, плейстоценовые ледники отступили и открыли миграционный путь, связавший эти южные популяции с северными районами, где они в настоящее время пересекаются с A. m. krausei и мигрировали на север, в долину Пис-Ривер (Канада) . [ 11 ] Фергюсон также отметил интерградацию в том же географическом районе, но между морфологическими подвидами A. m. columbianum и A.m. krausei , которые проходят параллельно хребтам Биттеррут и Селкирк . [ 18 ] Томпсон и Рассел предполагают, что эта зона контакта находится между двумя разными линиями подвидов, потому что A. m. Columbianum географически изолирована и ограничена горами центрального Орегона. [ 11 ]
Описание
[ редактировать ]Тело длиннопалой саламандры темно-черное с полосой на спине коричневого, желтого или оливково-зеленого цвета. Эту полосу также можно разбить на ряд пятен. Бока тела могут иметь мелкие белые или бледно-голубые крапинки. Брюшко темно-коричневого или сажистого цвета с белыми крапинками. Корневые бугорки присутствуют, но они не так развиты, как у других видов, например у тигровой саламандры . [ 19 ]
Яйца этого вида похожи на яйца родственной северо-западной саламандры ( A. gracile ) и тигровой саламандры ( A. tigrinum ). [ 26 ] Как и у многих земноводных, яйца длиннопалой саламандры окружены желатиновой капсулой. Эта капсула прозрачна, что делает эмбрион видимым во время развития. [ 19 ] В отличие от яиц A. gracile , здесь нет видимых признаков зеленых водорослей , что придает яичному студню зеленый цвет. длиннопалой саламандры В яйце эмбрион темнее сверху и белее снизу по сравнению с эмбрионом тигровой саламандры, который имеет цвет от светло-коричневого до серого сверху и кремовый снизу. Яйца имеют диаметр около 2 мм (0,08 дюйма) или больше с широким внешним слоем желе. [ 26 ] [ 27 ] До вылупления — как в яйце, так и в виде новорожденных личинок — у них есть балансиры — тонкие кожные выступы, торчащие по бокам и поддерживающие голову. Балансиры со временем отпадают, а их внешние жабры увеличиваются. [ 28 ] После потери балансиров личинки отличаются резко заостренным расширением жабр. личинок По мере взросления и метаморфоза их конечности с пальцами становятся видимыми, а жабры рассасываются. [ 26 ] [ 28 ]
Кожа личинки испещрена черной, коричневой и желтой пигментацией. Цвет кожи меняется по мере развития личинок, а пигментные клетки мигрируют и концентрируются в разных участках тела. Пигментные клетки, называемые хроматофорами , происходят из нервного гребня . Три типа пигментных хроматофоров саламандр включают желтые ксантофоры, черные меланофоры и серебристые иридиофоры (или гуанофоры). [ 29 ] [ 30 ] По мере взросления личинок меланофоры концентрируются вдоль тела и создают более темный фон. Желтые ксантофоры располагаются вдоль позвоночника и на верхней части конечностей. Остальная часть тела испещрена светоотражающими иридиофорами по бокам и снизу. [ 29 ] [ 18 ]
По мере метаморфоза личинки у них развиваются пальцы из выступов зачатков конечностей. Полностью метаморфизованная длиннопалая саламандра имеет четыре пальца на передних конечностях и пять пальцев на задних конечностях. [ 31 ] Его голова длиннее, чем ширина, а длинный внешний четвертый палец на задней конечности зрелых личинок и взрослых особей отличает этот вид от других, а также является этимологическим происхождением его видового эпитета : макродактилум (греч. makros = длинный и daktylos = палец ноги) . ). [ 32 ] Кожа взрослой особи имеет темно-коричневый, темно-серый или черный фон с желтой, зеленой или тускло-красной пятнистой полосой с точками и пятнами по бокам. Под конечностями, головой и телом саламандра имеет цвет от белого, розоватого до коричневого с более крупными белыми и более мелкими желтыми пятнами. Взрослые особи обычно имеют длину 3,8–7,6 см (1,5–3,0 дюйма). [ 26 ] [ 20 ]
Среда обитания и распространение
[ редактировать ]Длиннопалая саламандра — экологически универсальный вид, обитающий в самых разных средах обитания: от тропических лесов умеренного пояса , хвойных лесов , горных прибрежных зон , полынных равнин, красных еловых лесов, полузасушливых полынных равнин , равнин с читграсом до альпийских лугов вдоль скалистых берегов гор. озера. [ 19 ] [ 18 ] [ 25 ] Взрослые особи могут располагаться в лесном подлеске , прячась под грубыми древесными остатками , камнями, а также в норах мелких млекопитающих. В весенний период размножения взрослых особей можно найти под мусором или на прибрежных отмелях рек, ручьев, озер и прудов. Часто посещаются эфемерные воды. [ 19 ]
Этот вид — один из наиболее широко распространенных саламандр в Северной Америке, уступающий только тигровой саламандре . Его высотный диапазон простирается от уровня моря до 2800 метров (9200 футов), охватывая самые разнообразные растительные зоны. [ 19 ] [ 18 ] [ 33 ] [ 34 ] [ 35 ] Ареал включает изолированные эндемичные популяции в заливе Монтерей и Санта-Крус, Калифорния . [ 36 ] Распространение возобновляется на северо-востоке Сьерра-Невады, непрерывно проходя вдоль Тихоокеанского побережья до Джуно, Аляска , с популяциями, разбросанными по долинам рек Таку и Стикин . От побережья Тихого океана ареал простирается в продольном направлении до восточных предгорий Скалистых гор в Монтане и Альберте . [ 37 ] [ 21 ] [ 38 ]
Экология и жизненный цикл
[ редактировать ]
Яйца
[ редактировать ]Как и у всех земноводных, жизнь длиннопалой саламандры начинается с яйца. В северной части ареала яйца откладываются комковатыми массами на траве, палках, камнях или илистом субстрате спокойного пруда. [ 39 ] Количество яиц в одной массе колеблется по размеру, возможно, до 110 яиц в грозди. [ 40 ] Самки вкладывают значительные ресурсы в производство яиц, при этом яичники составляют более 50% массы тела в период перед размножением. У одной самки было обнаружено максимум 264 яйца - большое количество, учитывая, что каждое яйцо имеет диаметр примерно 0,5 миллиметра (0,02 дюйма). [ 41 ] Яичная масса удерживается желеобразным внешним слоем, защищающим внешнюю капсулу отдельных яиц. [ 42 ] Яйца иногда откладываются поодиночке, особенно в более теплом климате к югу от границы Канады и США. Яичное желе обеспечивает ежегодный запас биологического материала, который поддерживает химический состав и динамику питательных веществ мелководных водных экосистем и прилегающих лесных экосистем . [ 43 ] Яйца также служат средой обитания для водяных плесневых грибов, также известных как оомицеты . [ 44 ]

Личинки
[ редактировать ]Личинки вылупляются из яичной оболочки через две-шесть недель. [ 39 ] Они рождаются хищниками , инстинктивно питаясь мелкими беспозвоночными , которые перемещаются в поле их зрения . В число пищевых объектов входят мелкие водные ракообразные ( кладоцеры , копеподы и остракоды ), водные двукрылые и головастики . [ 45 ] По мере развития они естественным образом питаются более крупной добычей . Чтобы увеличить свои шансы на выживание, некоторые особи отрастают большие головы и становятся каннибалами и питаются своими товарищами по выводку. [ 46 ]
Метаморфозы и молодые особи
[ редактировать ]После того, как личинки подрастут и созреют, в течение как минимум одного сезона (личиночный период на Тихоокеанском побережье длится около четырех месяцев ), [ 37 ] они поглощают свои жабры и превращаются леса в наземных молодых особей, которые бродят по зарослям . Метаморфозы наблюдались уже в июле на уровне моря. [ 47 ] для А. м. croceum в октябре-ноябре и даже в январе. [ 20 ] На возвышенностях личинки могут перезимовать , развиваться и расти в течение дополнительного сезона, прежде чем метаморфизоваться. [ 48 ] В озерах на возвышенностях личинки могут достигать размеров 47 миллиметров (1,9 дюйма) от длины морды до длины устья (SVL) при метаморфозе, но на более низких высотах они развиваются быстрее и метаморфизируются, когда достигают 35–40 миллиметров (1,4–1,6 дюйма). СВЛ. [ 23 ]
Взрослые
[ редактировать ]Взрослые длиннопалые саламандры часто остаются незамеченными, потому что они ведут подземный образ жизни: копают, мигрируют и питаются беспозвоночными в лесных почвах, гниющих бревнах, небольших грызунов норах или трещинах в скалах . Рацион взрослой особи состоит из насекомых, головастиков, червей, жуков и мелкой рыбы. саламандр охотятся На подвязочные змеи , мелкие млекопитающие, птицы и рыбы. [ 49 ] Взрослая особь может жить 6–10 лет, при этом самые крупные особи весят примерно 7,5 граммов (0,26 унции), длина от морды до вентиляционных отверстий достигает 8 см (3,1 дюйма), а общая длина достигает 14 см (5,5 дюйма). [ 50 ] [ 51 ]
Поведение
[ редактировать ]Сезонный
[ редактировать ]История жизни длиннопалой саламандры сильно варьируется в зависимости от высоты и климата. Сезонные даты миграции в нерестовые пруды и обратно могут быть связаны с приступами продолжительных дождей, таянием льда или таянием снега, достаточными для пополнения (часто) сезонных прудов. Икра может нереститься на низких высотах уже в середине февраля в южном Орегоне . [ 52 ] с начала января по июль на северо-западе Вашингтона , [ 53 ] с января по март на юго-востоке Вашингтона, [ 54 ] и с середины апреля до начала мая в национальном парке Уотертон-Лейкс , Альберта. [ 55 ] Время размножения может сильно различаться; Следует отметить, что несколько яичных масс на ранних стадиях развития были обнаружены 8 июля 1999 года вдоль Британская Колумбия границы провинции недалеко от Джаспера, Альберта . [ 10 ] Взрослые особи сезонно мигрируют, чтобы вернуться в свои родные пруды для размножения, при этом самцы прибывают раньше и остаются там дольше, чем самки, а некоторые особи были замечены мигрирующими по сугробам в теплые весенние дни. [ 56 ] Гендерные различия (или половой диморфизм ) у этого вида проявляются только во время сезона размножения, когда у зрелых самцов появляется увеличенная или выпуклая область вентиляционного отверстия.
Разведение
[ редактировать ]
Время размножения зависит от высоты и широты места обитания саламандры. Обычно саламандры, расположенные на более низких высотах, размножаются осенью, зимой и ранней весной. Высотные саламандры размножаются весной и в начале лета. Особенно в более высоком климате саламандры заходят в пруды и озера, где все еще плавает лед. [ 19 ]
Взрослые особи собираются в больших количествах (>20 особей) под камнями и бревнами вдоль непосредственной границы мест размножения и бурно размножаются в течение нескольких дней. [ 20 ] Подходящие места для размножения включают небольшие пруды, свободные от рыбы, болота, мелкие озера и другие водно-болотные угодья со стоячей водой. [ 57 ] Как и другие амбистоматидные саламандры, они развили характерный брачный танец , во время которого они трутся телами и выделяют феромоны из подбородочной железы, прежде чем принять позицию для совокупления . Оказавшись в нужном положении, самец откладывает сперматофор , который представляет собой липкий стебель с пакетиком спермы на конце, и ведет самку вперед для оплодотворения . Самцы могут спариваться более одного раза и откладывать до 15 сперматофоров в течение пятичасового периода. [ 20 ] [ 39 ] Брачный танец длиннопалой саламандры похож на танец других видов Ambystoma и очень похож на A. jeffersonianum . [ 58 ] [ 59 ] У длиннопалой саламандры нет трения и бодания головой; самцы напрямую приближаются к самкам и хватаются за нее, а самки стараются быстро уплыть. [ 59 ] Самцы хватают самку за передние конечности и трясут – такое поведение называется амплексусом . Во время размножения самцы иногда обхватывают другие виды амфибий и трясут их. [ 53 ] Самец хватается только передними конечностями и никогда не использует задние конечности во время брачного танца, поскольку он трет подбородок из стороны в сторону, надавливая на голову самки. Женщина борется, но позже становится подавленной. Самцы увеличивают темп и движения, потирая ноздри самки, бока, а иногда и вентиляционные отверстия. Когда самка становится достаточно послушной, самец движется вперед, поднимая над ее головой хвост и машет кончиком. Если самка принимает ухаживание самца, самец направляет ее морду к своей области жерлового отверстия, в то время как оба с трудом двигаются вперед, покачивая тазом. Когда самка следует за ней, самец останавливается и откладывает сперматофор, а самка движется вперед вместе с самцом, чтобы поднять хвост и получить пакет со спермой. Полный брачный танец редко удается осуществить с первой попытки. [ 59 ] Самки откладывают яйца через несколько дней после спаривания. [ 20 ]
Хранение энергии и защитные механизмы
[ редактировать ]

В некоторых равнинных районах взрослые саламандры остаются активными всю зиму, за исключением похолоданий. Однако в холодные зимние месяцы в северных частях своего ареала длиннопалая саламандра зарывается ниже линии замерзания в грубом субстрате и зимует группами по 8–14 особей. [ 40 ] [ 60 ] В состоянии спячки он выживает за счет запасов белковой энергии, которые хранятся в его коже и вдоль хвоста. [ 61 ] Эти белки выполняют второстепенную функцию как часть смеси кожных выделений, которая используется для защиты. [ 62 ] При угрозе длиннопалая саламандра взмахивает хвостом и выделяет липкое белое молочное вещество, которое ядовито и, вероятно, ядовито. [ 55 ] [ 63 ] Цвет его кожи может служить предупреждением хищникам ( апосематизм ), что он будет невкусным. [ 62 ] Цвета и узоры его кожи разнообразны: от темно-черного до красновато-коричневого фона с пятнами или пятнами бледно-красновато-коричневого, бледно-зеленого цвета до ярко-желтой полосы. [ 37 ] [ 39 ] Взрослая особь также может уронить часть своего хвоста и ускользнуть, в то время как хвостовой кусочек действует как извилистая приманка; это называется автотомией . [ 64 ] Регенерация и повторный рост хвоста — один из примеров развития физиологии земноводных, представляющий большой интерес для медицины. [ 65 ]
Статус сохранения
[ редактировать ]Хотя длиннопалая саламандра классифицируется МСОП как вызывающая наименьшее беспокойство , [ 1 ] Многие формы освоения земель отрицательно влияют на среду обитания саламандр и открывают новые перспективы и приоритеты в биологии их сохранения . Приоритеты сохранения сосредоточены на уровне разнообразия популяций, которое сокращается темпами в десять раз быстрее, чем вымирание видов. [ 66 ] [ 67 ] [ 68 ] [ 69 ] Разнообразие на уровне населения – это то, что обеспечивает экосистемные услуги . [ 70 ] например, ключевую роль, которую саламандры играют в почвенных экосистемах, включая круговорот питательных веществ , который поддерживает водно-болотные и лесные экосистемы. [ 71 ]
Две особенности жизненного цикла земноводных часто называют причиной того, почему земноводные являются хорошими индикаторами состояния окружающей среды или «канарейками в угольной шахте». Как и все земноводные, длиннопалая саламандра имеет как водный, так и наземный переход к жизни и полупроницаемую кожу. Поскольку в воде они выполняют разные экологические функции, чем на суше, потеря одного вида земноводных эквивалентна утрате двух экологических видов. [ 72 ] Второе мнение состоит в том, что земноводные, такие как длиннопалые саламандры, [ 73 ] более восприимчивы к поглощению загрязняющих веществ, поскольку они естественным образом поглощают воду и кислород через кожу. Однако обоснованность этой особой чувствительности к загрязнителям окружающей среды была поставлена под сомнение. [ 74 ] Проблема более сложна, потому что не все земноводные одинаково восприимчивы к экологическому ущербу, поскольку у видов очень разнообразный жизненный цикл. [ 75 ]
Популяциям длиннопалых саламандр угрожает фрагментация , интродуцированные виды и УФ-излучение . Лесное хозяйство, дороги и другие освоения земель изменили среду, в которую мигрируют земноводные, и увеличили смертность . [ 76 ] В таких местах, как национальный парк Уотертон-Лейкс, установлен подземный автомобильный туннель, чтобы обеспечить безопасный проход и поддержать экологию миграции этого вида. [2] Распространение длиннопалой саламандры во многом пересекается с лесной промышленностью, доминирующим ресурсом, поддерживающим экономику Британской Колумбии и западной части Соединенных Штатов. Длиннопалые саламандры изменят миграционное поведение, и на них негативно повлияет практика лесного хозяйства, не обеспечивающая значительных буферов управления и защиты для небольших водно-болотных угодий, где размножаются саламандры. [ 77 ] [ 78 ] Население вблизи долины Пис-Ривер в Альберте погибло в результате расчистки и осушения водно-болотных угодий для нужд сельского хозяйства. [ 79 ] Форель , завезенная для спортивного рыболовства в озера, когда-то безрыбные, также уничтожает популяции длиннопалых саламандр. [ 80 ] Интродуцированные золотые рыбки питаются икрой и личинками длиннопалых саламандр. [ 81 ] Повышенное воздействие UVB- излучения является еще одним фактором, причастным к глобальному сокращению численности земноводных, и длиннопалая саламандра также восприимчива к этой угрозе, что увеличивает частоту уродств и снижает их выживаемость и скорость роста. [ 82 ] [ 83 ] [ 84 ]
Подвид Ambystoma macrodactylum croceum ( Длиннопалая саламандра Санта-Крус ) вызывает особую озабоченность, и в 1967 году ему была предоставлена защита в соответствии с Законом США об исчезающих видах . [ 85 ] Этот подвид обитает в узком диапазоне ареалов в округах Санта-Крус и Монтерей , Калифорния. До получения защиты некоторые оставшиеся популяции находились под угрозой развития. Этот подвид экологически уникален, имеет уникальный неровный рисунок кожи на спине, уникальную устойчивость к влаге, а также является эндемиком , географически изолированным от остального ареала вида. [ 18 ] [ 86 ] [ 87 ] [ 88 ] Другие подвиды включают A. m. columbianum , A.m. краузей , А.м. macrodactylum и A.m. сигиллатум . [ 21 ]
См. также
[ редактировать ]- Длиннопалая саламандра Санта-Крус , исчезающий подвид.
Ссылки
[ редактировать ]- ^ Jump up to: а б Группа специалистов МСОП по амфибиям SSC (2015 г.). « Амбистома макродактилум » . Красный список исчезающих видов МСОП . 2015 : e.T59063A56539990. doi : 10.2305/IUCN.UK.2015-4.RLTS.T59063A56539990.en . Проверено 19 ноября 2021 г.
- ^ Jump up to: а б Фрост, Даррел Р. (2018). « Ambystoma macrodactylum Baird, 1850» . Виды амфибий мира: онлайн-справочник. Версия 6.0 . Американский музей естественной истории . Проверено 23 марта 2018 г.
- ^ Тихен Дж (1958). «Комментарии к остеологии и филогении амбистоматидных саламандр» . Бюллетень Государственного музея Флориды . 3 (1): 1–50 . Проверено 11 января 2010 г.
- ^ Jump up to: а б с Джонс Т.Р., Клюге А.Г., Вольф А.Дж. (1993). «Когда теории и методологии сталкиваются: филогенетический повторный анализ североамериканских амбистоматидных саламандр (Caudata: Amybstomatidae)». Систематическая биология . 42 (1): 92–102. дои : 10.1093/sysbio/42.1.92 .
- ^ Винс Джей-Джей (2007). «Глобальные закономерности диверсификации и видового богатства амфибий» (PDF) . Американский натуралист . 170 (С2): С86–С106. дои : 10.1086/519396 . ПМИД 17874387 . S2CID 36017698 . Архивировано из оригинала (PDF) 7 декабря 2010 г. Проверено 9 февраля 2011 г.
- ^ Чжан П., Wake DB (2009). «Отношения саламандр более высокого уровня и даты расхождения, выведенные из полных митохондриальных геномов» (PDF) . Молекулярная филогенетика и эволюция . 53 (2): 492–508. дои : 10.1016/j.ympev.2009.07.010 . ПМИД 19595776 .
- ^ Ларсон А. (1996). «Амбистоматиды» . Веб-проект «Древо жизни» . Архивировано из оригинала 03 марта 2021 г. Проверено 14 января 2010 г.
- ^ Милнер А.Р. (1983). «Биогеография саламандр в мезозое и раннем кайнозое: кладистически-викариантная модель». В Sims RW, Price JH, Whalley PE (ред.). Эволюция, время и пространство: возникновение биосферы . Специальный выпуск Ассоциации Систематики. Том. 23. Лондон: Академик Пресс. стр. 431–468. ISBN 978-0-12-644550-3 .
- ^ Jump up to: а б с д Дуэллман Э.В. (1999). Закономерности распространения земноводных: глобальная перспектива . Джу Пресс. п. 633. ИСБН 978-0-8018-6115-4 . Проверено 12 января 2010 г.
- ^ Jump up to: а б с д и ж г Томпсон, Марк Д. (2003). Филогеография длиннопалой саламандры Ambystoma macrodactylum (магистерская диссертация). Университет Калгари . ISBN 978-0-612-87451-0 . OCLC 150649401 .
- ^ Jump up to: а б с д и ж г час я дж к л м н Томпсон, доктор медицинских наук, Рассел А.П. (2005). «Отступление ледника и его влияние на миграцию митохондриальных генов у длиннопалой саламандры ( Ambystoma macrodactylum ) в западной части Северной Америки». В Элева АМТ (ред.). Климатология, география, экология: причины миграции организмов . Гейдельберг, Германия: Издательство Springer-Verlag. стр. 205–246. ISBN 978-3-540-26603-7 .
- ^ Милнер А.Р. (1983). Биогеография саламандр в мезозое и раннем кайнозое: кладистически-викариантная модель. В Sims, RW, Price JH, Whalley PES. (Ред.), Эволюция, время и пространство: возникновение биосферы. (стр. 431–468) Том. 23 Ассоциации Систематики, Специальный Том. Академическое издательство, Лондон.
- ^ Нуссбаум РА. (1974). Географическая изменчивость и систематика саламандр рода Dicamptodon Strauch (Ambystomatidae). 94 стр. Разные публикации Зоологического музея Мичиганского университета, № 149.
- ^ Jump up to: а б Добенмайр Р. (март 1975 г.). «География флористических растений Восточного Вашингтона и Северного Айдахо». Журнал биогеографии . 2 (1): 1–18. дои : 10.2307/3038197 . JSTOR 3038197 .
- ^ Первоначальный источник, описывающий палеоэкологические аналоги, цитируемый Томпсоном (2003), см.: Heusser C, Minneapolis (1983). История растительности северо-запада США, включая Аляску: поздний плейстоцен. В: Райт Х., Портер С. (ред.). Позднечетвертичная среда США. (стр. 239–258) Университет Миннесоты Press.
- ^ Jump up to: а б Брунсфельд С., Салливан Дж., Солтис Д., Солтис П. (2001). Сравнительная филогеография северо-запада Северной Америки: синтез. В: Сильвертон Дж., Антонович Дж. (ред.), Интеграция экологии и эволюции в пространственном контексте. 14-й специальный симпозиум Британского экологического общества. Британское экологическое общество, Blackwell Science Ltd., Ch. 15, стр. 319–339.
- ^ Стил, Калифорния; Карстенс, Британская Колумбия; Сторфер, А.; Салливан, Дж. (2005). «Проверка гипотезы о времени видообразования у Dicamptodon copyi и Dicamptodon aterrimus (Caudata: Dicamptodontidae)» (PDF) . Молекулярная филогенетика и эволюция . 36 (1): 90–100. дои : 10.1016/j.ympev.2004.12.001 . ПМИД 15904859 . Архивировано из оригинала (PDF) 14 августа 2010 г.
- ^ Jump up to: а б с д и ж г час я дж Фергюсон Д.Э. (1961). «Географическая вариация Ambystoma macrodactylum Baird с описанием двух новых подвидов». Американский натуралист из Мидленда . 65 (2): 311–338. дои : 10.2307/2422958 . JSTOR 2422958 .
- ^ Jump up to: а б с д и ж г час Стеббинс Р.А. (2003). Полевой справочник по западным рептилиям и амфибиям (Серия полевых справочников Петерсона) (3-е изд.). Бостон: Хоутон Миффлин. ISBN 978-0-395-98272-3 .
- ^ Jump up to: а б с д и ж г Петранка Ю.В. (1998). Саламандры США и Канады . Вашингтон, округ Колумбия: Смитсоновские книги. ISBN 978-1-56098-828-1 .
- ^ Jump up to: а б с Нуссбаум Р.А.; Броди ЭД-младший; Шторм РМ. (1983). Земноводные и рептилии северо-запада Тихого океана . Москва, Айдахо: Университетское издательство Айдахо. ISBN 978-0-89301-086-7 .
- ^ Таллмон Д.А., Фанк WC, Данлэп WW, Аллендорф FW (2000). МакИхран Дж.Д. (ред.). «Генетическая дифференциация популяций длиннопалых саламандр ( Ambystoma macrodactylum )». Копейя . 2000 (1): 27–35. doi : 10.1643/0045-8511(2000)2000[0027:GDALTS]2.0.CO;2 . JSTOR 1448236 . S2CID 37951483 .
- ^ Jump up to: а б Ховард Дж. Х., Уоллес Р. Л. (1981). «Микрогеографическая изменчивость электрофоретических локусов в популяциях Ambystoma macrodactylum columbianum (Caudata: Ambystomatidae)». Копейя . 1981 (2): 466–471. дои : 10.2307/1444241 . JSTOR 1444241 .
- ^ Крайняя высота варьируется в зависимости от климата, но> 2200 метров (7200 футов), вероятно, станет препятствием для распространения на большей части ареала этого вида к северу от Орегона. См. также: Джордано А.Р., Риденхор Б.Дж., Сторфер А. (2008). Влияние высоты и топографии на генетическую структуру длиннопалой саламандры ( Ambystoma macrodactulym ). Молекулярная экология 16 (8): 1625–1637. PDF
- ^ Jump up to: а б Грэм К.Л., Пауэлл Г.Л. (1999). Статус длиннопалой саламандры ( Ambystoma macrodactylum ) в Альберте. Отдел защиты окружающей среды, рыболовства и управления дикой природой Альберты и Ассоциация охраны природы Альберты, Отчет о состоянии дикой природы № 22 (PDF) . Эдмонтон, Альберта, Канада: Отдел охраны окружающей среды, рыболовства и управления дикой природой Альберты и Ассоциация охраны природы Альберты. п. 1. Архивировано из оригинала (PDF) 6 июля 2011 г. Проверено 15 января 2010 г.
- ^ Jump up to: а б с д Томс С., Коркран CC (2006). Амфибии Орегона, Вашингтона и Британской Колумбии: Руководство по полевой идентификации (Полевые справочники Lone Pine) . Эдмонтон, Альберта, Канада: Lone Pine Publishing. ISBN 978-1-55105-566-4 .
- ^ Салте С.Н. (1963). «Яйковые капсулы у амфибий». Журнал морфологии . 113 (2): 161–171. дои : 10.1002/jmor.1051130204 . ПМИД 14065317 . S2CID 22749214 .
- ^ Jump up to: а б Уотсон С., Рассел А.П. (2000). «Таблица стадий развития длиннопалой саламандры после вылупления Ambystoma macrodactylum krausei » (PDF) . Земноводные-рептилии . 21 (2): 143–154. дои : 10.1163/156853800507336 . Проверено 14 января 2010 г.
- ^ Jump up to: а б Паричий Д.М. (1996). «Пигментные узоры личинок саламандр (Ambystomatidae, Salamandridae): роль сенсорной системы боковой линии и эволюция механизмов формирования узоров» . Биология развития . 175 (2): 265–282. дои : 10.1006/dbio.1996.0114 . ПМИД 8626032 .
- ^ Педерзоли А., Гамбарелли А., Рестани С. (2003). «Миграция ксантофоров из дермы в эпидермис и ремоделирование кожи во время развития личинок Salamandra salamandra salamandra (L.)». Исследование пигментных клеток . 16 (1): 50–58. дои : 10.1034/j.1600-0749.2003.00013.x . ПМИД 12519125 .
- ^ Уотсон, Шери М (1997). Влияние уровня пищи на сроки метаморфизма у длиннопалой саламандры Ambystoma macrodactylum krausei (магистерская диссертация). Университет Калгари . ISBN 978-0-612-20859-9 . OCLC 150699685 .
- ^ Бэрд С.Ф. (1849 г.). «Пересмотр североамериканских хвостатых батрахий с описаниями новых родов и видов - описание четырех новых видов североамериканских саламандр и одного нового вида сцинков». Журнал Академии естественных наук Филадельфии . 1 (4): 281–292.
- ^ Ховард Дж. Х., Уоллес Р.Л. (1985). «Характеристики истории жизни популяций длиннопалой саламандры ( Ambystoma macrodactylum ) с разных высот». Американский натуралист из Мидленда . 133 (2): 361–373. дои : 10.2307/2425582 . JSTOR 2425582 .
- ^ Фанк WC, Данлэп WW (1999). «Заселение высокогорных озер длиннопалыми саламандрами ( Ambystoma macrodactylum ) после вымирания интродуцированных популяций форели» . Канадский журнал зоологии . 77 (11): 1759–1767. дои : 10.1139/cjz-77-11-1759 .
- ^ Джордано А.Р., Риденхор Б.Дж., Сторфер А. (апрель 2007 г.). «Влияние высоты и топографии на генетическую структуру длиннопалой саламандры ( Ambystoma macrodactulym )» (PDF) . Молекулярная экология . 16 (8): 1625–1637. дои : 10.1111/j.1365-294X.2006.03223.x . ПМИД 17402978 . S2CID 32775223 . Проверено 14 января 2010 г.
- ^ Рассел Р.В., Андерсон Дж.Д. (1956). «Дизъюнктивная популяция длиннопалой саламандры с побережья Калифорнии». Герпетологика . 12 : 137–140.
- ^ Jump up to: а б с Карл Г.К. (1950). Земноводные Британской Колумбии. 3-е изд. Справочник №2 . Виктория, Британская Колумбия: Провинциальный музей Британской Колумбии, Департамент образования.
- ^ Информацию о дистрибутивах Аляски см. в MacDonald SO. Земноводные и рептилии Аляски . См. также: Норман, БР (1999). «Географическое распространение: Ambystoma macrodactylum ». Герпетологическое обозрение . 30 : 171.
- ^ Jump up to: а б с д Грин ДМ, Кэмпбелл РВ. (1992). Земноводные Британской Колумбии. Справочник Королевского музея Британской Колумбии № 45. Провинция Британская Колумбия, Министерство туризма и Министерство, ответственное за культуру.
- ^ Jump up to: а б Томпсон, доктор медицины (2001). «Необычайно искусная амбистоматида, длиннопалая саламандра, справляющаяся с крайностями севера» (PDF) . Бореальная сеть погружений . 5 (2): 8–10.
- ^ Веррелл П. (2007). «Женский репродуктивный цикл североамериканской саламандры Ambystoma macrodactylum columbianum » . Земноводные-рептилии . 27 (2): 274–277. дои : 10.1163/156853806777239887 .
- ^ Труеб Л., Дуэллман В.Е. (1994). Биология амфибий . Балтимор: Издательство Университета Джонса Хопкинса. п. 112. ИСБН 978-0-8018-4780-6 . Проверено 06 марта 2010 г.
- ^ Регестер К.Дж., В то время как М.Р. (2006). Тейлор CM (ред.). «Скорость разложения стадий жизни саламандры ( Ambystoma maculatum ) и связанные с этим потоки энергии и питательных веществ в прудах и прилегающих лесах на юге Иллинойса». Копейя . 2006 (4): 640–649. doi : 10.1643/0045-8511(2006)6[640:DROSAM]2.0.CO;2 . JSTOR 4126531 . S2CID 85753900 .
- ^ Петриско Дж.Э., Перл К.А., Пиллиод Д.С., Шеридан П.П., Уильямс К.Ф., Петерсон С.Р., Бери Б.Р. (2008). «Saprolegniaceae идентифицированы на яйцах амфибий на северо-западе Тихого океана, США, с помощью внутренних транскрибируемых спейсерных последовательностей и филогенетического анализа» (PDF) . Микология . 100 (2): 171–180. дои : 10.3852/микология.100.2.171 . ПМИД 18592894 . Проверено 7 марта 2010 г.
- ^ Андерсон Дж. Д. (1968). «Сравнение пищевых привычек Ambystoma macrodactylum sigillatum , Ambystoma macrodactylum croceum и Ambystoma tigrinum Californiense ». Герпетологика . 24 (4): 273–284. JSTOR 3891365 .
- ^ Уоллс С.К., Белангер С.С., Блауштайн А.Р. (1993). «Морфологические вариации личинки саламандры: диетическая индукция пластичности формы головы». Экология . 96 (2): 162–168. Бибкод : 1993Oecol..96..162W . дои : 10.1007/BF00317728 . ПМИД 28313411 . S2CID 24146096 .
- ^ Кезер Дж., Фарнер Д.С. (1955). «Особенности истории жизни саламандры Ambystoma macrodactylum в Высоких Каскадных горах Южного Орегона». Копейя . 1955 (2): 127–131. дои : 10.2307/1439318 . JSTOR 1439318 .
- ^ Марнелл Л.Ф. (1997). «Герпетофауна национального парка Глейшер». Северо-западный натуралист . 78 (1): 17–33. дои : 10.2307/3536855 . JSTOR 3536855 .
- ^ Грегори П.Т., Мацуда Б.М., Грин Д. (2006). Земноводные и рептилии Британской Колумбии . Виктория: Королевский музей Британской Колумбии. ISBN 978-0-7726-5448-9 .
- ^ Рассел А.П., Пауэлл Г.Л., Холл Д.Р. (1996). «Рост и возраст длиннопалых саламандр Альберты ( Ambystoma macrodactylum krausei ): сравнение двух методов оценки» (PDF) . Канадский журнал зоологии . 74 (3): 397–412. дои : 10.1139/z96-047 . Проверено 7 марта 2010 г.
- ^ Дополнение к диапазону веса и размеров взято из базы данных амфибий NAMOS BC [1] . Архивировано 27 апреля 2017 г. на Wayback Machine .
- ^ Кезер Дж., Фарнер Д.С. (1955). «Образцы истории жизни саламандры Ambystoma macrodactylum в высоких Каскадных горах южного Орегона». Копейя . 1955 (2): 127–131. дои : 10.2307/1439318 . JSTOR 1439318 .
- ^ Jump up to: а б Слейтер-младший (1936). «Заметки об Ambystoma gracile Baird и Ambystoma macrodactylum Baird». Копейя . 1936 (4): 234–236. дои : 10.2307/1436330 . JSTOR 1436330 .
- ^ Веррелл П., Пелтон Дж. (1996). «Сексуальная стратегия центральной длиннопалой саламандры Ambystoma macrodactylum columbianum на юго-востоке Вашингтона». Журнал зоологии . 240 : 37–50. дои : 10.1111/j.1469-7998.1996.tb05484.x .
- ^ Jump up to: а б Фукумото Дж.М. (1995). Экология и управление длиннопалой саламандрой ( Ambystoma macrodactylum ) в национальном парке Уотертон-Лейкс (диссертация ME). Университет Калгари . ISBN 978-0-612-04397-8 . OCLC 70487881 .
- ^ Бенески-младший; Залиско Е.Ю.; Ларсен-младший (1986). «Демография и характер миграции восточной длиннопалой саламандры Ambystoma macrodactylum columbianum ». Копейя . 2 (2): 398–408. дои : 10.2307/1444998 . JSTOR 1444998 .
- ^ Стеббинс RC, Коэн NW. (1995). Естественная история амфибий . Издательство Принстонского университета ISBN 0-691-10251-1 .
- ^ Кнудсен Дж.В. (1960). «Ухаживание и яичная масса Ambystoma gracile и Ambystoma macrodactylum ». Копейя . 1 (1): 44–46. дои : 10.2307/1439844 . JSTOR 1439844 .
- ^ Jump up to: а б с Андерсон Дж. Д. (1961). «Брачное поведение Ambystoma macrodactylum croceum ». Копейя . 1961 (2): 132–139. дои : 10.2307/1439987 . JSTOR 1439987 .
- ^ Шеппард, Роберт Франк (1997). Экология и ареал передвижения Ambystoma macrodactylum krausei (Amphibia:Urodela) (диплом магистра наук). Университет Калгари . OCLC 15847219 .
- ^ Уильямс, Томас А; Ларсен, Джон Х (1986). «Новая функция зернистых кожных желез восточной длиннопалой саламандры Ambystoma macrodactylum columbianum ». Журнал экспериментальной зоологии . 239 (3): 329–333. дои : 10.1002/jez.1402390304 .
- ^ Jump up to: а б Грант Дж. Б., Эванс Дж. А. (2007). «Методика сбора и анализа неклейких выделений кожи саламандр амбистоматид ». Герпетологическое обозрение . 38 (3): 301–5.
- ^ Толедо, Р. (1995). «Кожные зернистые железы и яды амфибий». Сравнительная биохимия и физиология А. 111 : 1–29. дои : 10.1016/0300-9629(95)98515-I .
- ^ «NAMOS BC (Северное общество мониторинга амфибий)» . Архивировано из оригинала 3 марта 2016 г. Проверено 24 июня 2009 г.
- ^ Одельберг С.Дж. (2005). «Клеточная пластичность в регенерации позвоночных». Анатомические записи, часть B: Новый анатом . 287 (1): 25–35. дои : 10.1002/ar.b.20080 . ПМИД 16308861 .
- ^ Блауштайн, Арканзас; Кизекер, Дж. М. (2002). «Сложность сохранения: уроки глобального сокращения популяций амфибий» (PDF) . Экологические письма . 5 (4): 597–608. дои : 10.1046/j.1461-0248.2002.00352.x .
- ^ Удачи, ГВ; Ежедневно, GC; Эрлих, PR (2003). «Разнообразие населения и экосистемные услуги» (PDF) . Тенденции экологии и эволюции . 18 (7): 331–336. дои : 10.1016/s0169-5347(03)00100-9 . Архивировано из оригинала (PDF) 5 февраля 2009 г. Проверено 15 января 2009 г.
- ^ Гаскон С., Коллинз Дж.П., Мур Р.Д., Черч Д.Р., Маккей Дж.Э., Мендельсон-младший III. (ред.). (2007). План действий по сохранению амфибий. Группа специалистов МСОП/SSC по земноводным. Гланд, Швейцария и Кембридж, Великобритания. 64 стр. PDF. Архивировано 4 июля 2007 г. в Wayback Machine.
- ^ Вуд, CW; Гросс, MR (2008). «Элементарные подразделения охраны природы: информирование о риске исчезновения без указания целей защиты» (PDF) . Биология сохранения . 22 (1): 36–47. дои : 10.1111/j.1523-1739.2007.00856.x . ПМИД 18254851 . S2CID 23211536 . Архивировано из оригинала (PDF) 1 октября 2018 г. Проверено 15 января 2009 г.
- ^ Карейва, П; Марвье, М. (2003). «Сохранение холодных точек биоразнообразия» (PDF) . Американский учёный . 91 (4): 344–351. дои : 10.1511/2003.26.869 . S2CID 120910582 . Архивировано из оригинала (PDF) 25 февраля 2009 г. Проверено 16 января 2009 г.
- ^ Давич, РД; Валлийский, Х.Х. младший (2004). «Об экологической роли саламандр» (PDF) . Ежегодный обзор экологии и систематики . 35 : 405–434. CiteSeerX 10.1.1.521.9086 . doi : 10.1146/annurev.ecolsys.35.112202.130116 .
- ^ В то время как, MR; Липс, КР; Прингл, CM; Килхэм, СС ; Биксби, Р.Дж.; Бренес, Р.; Коннелли, С.; и др. (2006). «Влияние сокращения популяции земноводных на структуру и функции экосистем неотропических ручьев» (PDF) . Границы в экологии и окружающей среде . 4 (1): 27–34. doi : 10.1890/1540-9295(2006)004[0027:teoapd]2.0.co;2 .
- ^ Джон, Фрейли (октябрь 2009 г.). Длиннопалая саламандра . Устойчивое развитие ресурсов Альберты. ISBN 978-0-7785-2002-3 .
{{cite book}}:|periodical=игнорируется ( помогите ) - ^ Коллинз, JP; Крамп, М. (2008). Вымирание в наше время: глобальное сокращение численности амфибий . Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-531694-0 .
- ^ Биби, TJC; Гриффитс, Р. (2005). «Кризис сокращения численности амфибий: переломный момент для природоохранной биологии?». Биологическая консервация . 125 (3): 271–285. doi : 10.1016/j.biocon.2005.04.009 .
- ^ Беккер, CG; Фонсека, ЧР; Хаддад, ЦФБ; Батиста, РФ; Прадо, ИП (2007). «Раскол среды обитания и глобальное сокращение численности амфибий». Наука . 318 (5857): 1775–1777. Бибкод : 2007Sci...318.1775B . дои : 10.1126/science.1149374 . ПМИД 18079402 . S2CID 22055213 .
- ^ Фергюсон К. (1999). Воздействие вырубки леса на длиннопалую саламандру ( Ambystoma macrodactylum ) на горе Опакс. Стр. 221–229 В книге К. Холлстедта, А. Вайса и Д. Хаггарда, ред. Новые сведения для управления сухими пихтовыми лесами Дугласа: Учеб. мастерская сухой дугласовой ели. Министр БК. для., Виктория, Британская Колумбия. PDF
- ^ Нотон, врач общей практики; Хендерсон, CB; Форесман, КР; МакГроу, RL II (2000). «Длиннопалые саламандры в вырубленных и нетронутых лесах дугласовой пихты западной Монтаны» . Экологические приложения . 10 (6): 1681–1689. doi : 10.1890/1051-0761(2000)010[1681:ltsiha]2.0.co;2 .
- ^ Уолш, Р. (1998). «Расширение известного ареала длиннопалой саламандры Ambystoma macrodactylum в Альберте» . Канадский полевой натуралист . 112 (2): 331–333. дои : 10.5962/стр.358412 .
- ^ Фанк, туалет; Данлэп, WW (1999). «Заселение высокогорных озер длиннопалыми саламандрами ( Ambystoma macrodactylum ) после вымирания интродуцированных популяций форели» . Канадский журнал зоологии . 77 (11): 1759–1767. дои : 10.1139/z99-160 . Архивировано из оригинала 7 июля 2012 г.
- ^ Монелло, Р.Дж.; Райт, Р.Г. (2001). «Хищничество золотой рыбки ( Carassius auratus ) на яйца и личинки восточной длиннопалой саламандры ( Ambystoma macrodactylum columbianum )». Журнал герпетологии . 35 (2): 350–353. дои : 10.2307/1566132 . JSTOR 1566132 .
- ^ Блауштайн, Арканзас; Кизекер, Дж. М.; Чиверс, ДП; Энтони, Р.Г. (1997). «Амбиентное УФ-излучение вызывает уродства у эмбрионов амфибий» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (25): 13735–13737. Бибкод : 1997PNAS...9413735B . дои : 10.1073/pnas.94.25.13735 . ПМК 28375 . ПМИД 9391095 .
- ^ Белден, ЛК; Уайлди, Эл.; Блаустейн, Арканзас (2000). «Рост, выживание и поведение личинок длиннопалых саламандр ( Ambystoma macrodactylum ), подвергшихся воздействию окружающего уровня УФ-B-излучения». Журнал зоологии (Лондон) . 251 (4): 473–479. дои : 10.1111/j.1469-7998.2000.tb00803.x .
- ^ Крото, MC; Дэвидсон, Массачусетс; Лин, ДР; Трюдо, В.Л. (2008). «Глобальное увеличение ультрафиолетового излучения B: потенциальное воздействие на развитие и метаморфоз земноводных» . Физиологическая и биохимическая зоология . 81 (6): 743–761. дои : 10.1086/591949 . ПМИД 18954263 . S2CID 31675246 . [ постоянная мертвая ссылка ]
- ^ «DFG - Программа дикой природы, не связанная с игрой - Амфибии, находящиеся под угрозой исчезновения» . Проверено 23 июня 2009 г.
- ^ Андерсон, доктор медицинских наук (1972). «Поведение трех подвидов Ambystoma macrodactylum в условиях градиента влажности почвы». Журнал герпетологии . 6 (3–4): 191–194. дои : 10.2307/1562770 . JSTOR 1562770 .
- ^ Рид Р.Дж. (1978). Исследование популяции длиннопалой саламандры Санта-Крус ( Ambystoma macrodactylum croceum ) в лагуне Валенсии в 1977–1978 годах с заметками о среде обитания и распространении в округах Санта-Крус и Монтерей. Калифорнийский отдел рыбы и дичи, контракт S-1180.
- ^ Фишер, Р.Н.; Шаффер, Х.Б. (2002). «Упадок численности амфибий в Большой Центральной долине Калифорнии». Биология сохранения . 10 (5): 1387–1397. дои : 10.1046/j.1523-1739.1996.10051387.x .
Внешние ссылки
[ редактировать ]![]() Данные, относящиеся к Ambystoma macrodactylum , в Wikispecies
Данные, относящиеся к Ambystoma macrodactylum , в Wikispecies
![]() СМИ, связанные с Ambystoma macrodactylum, на Викискладе?
СМИ, связанные с Ambystoma macrodactylum, на Викискладе?
| Базы данных органов управления : Национальные |
|---|