Карторинх
| Карторинх Временной диапазон: Спатиан ,
| |
|---|---|

| |
| Образец голотипа (AGB 6257), Геологический музей Китая. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Рептилии |
| Клэйд : | † Ихтиозавроморфы |
| Клэйд : | † Ихтиозавриформные |
| Клэйд : | † Насоростра |
| Род: | † Карторинх Мотани и др ., 2014 г. |
| Разновидность: | † C. lenticarpus
|
| Биномиальное имя | |
| † Cartorhynchus lenticarpus Мотани и др ., 2014 г.
| |
Cartorhynchus (что означает «укороченная морда») — вымерший род ихтиозаврообразных ранних , морских рептилий живших в раннего триаса эпоху , около 248 миллионов лет назад. Род включает единственный вид, Cartorhynchus lenticarpus , названный в 2014 году Рёсуке Мотани и его коллегами по единственному почти полному скелету, найденному недалеко от Чаоху , провинция Аньхой , Китай . Наряду со своим близким родственником Sclerocormus , Cartorhynchus был частью разнообразия морских рептилий, которое произошло внезапно (в течение примерно одного миллиона лет) во время спатского подэтапа , вскоре после разрушительного пермско-триасового вымирания , но впоследствии они были доведены до исчезновения из-за вулканизма. и изменения уровня моря в среднем триасе .
имел длину около 40 сантиметров (16 дюймов) и Карторинх был небольшим животным с телом, похожим на ящерицу, и коротким туловищем; вероятно, он плавал, как угорь, на медленной скорости. Его конечности имели обширные хрящи и могли сгибаться, как ласты, что, возможно, позволяло ему ходить по суше. Наиболее отличительными чертами Cartorhynchus были короткая, суженная морда и несколько рядов зубов, похожих на коренные зубы, которые росли на внутренней поверхности его челюстных костей. Эти зубы не были обнаружены до тех пор, пока образец не был подвергнут компьютерной томографии . Cartorhynchus , вероятно, охотился на беспозвоночных с твердым панцирем, используя всасывание , хотя, как именно он использовал свои направленные внутрь зубы, пока неизвестно. Это было одно из пяти независимых приобретений коренных зубов среди ихтиозаврообразных.
Открытие и присвоение имени
[ редактировать ]
В 2011 году единственный известный экземпляр Cartorhynchus был обнаружен в слое 633 второго уровня карьера Мацзяшань недалеко от центра города Чаоху , провинция Аньхой , Китай ; пласты горных пород в этом карьере принадлежат верхней части формации Наньлинху . [ 1 ] Экземпляр представляет собой почти полный скелет, у которого отсутствует только часть хвоста. [ 2 ] и некоторые кости левой части задней части черепа. Сохранность образца, вероятно, стала результатом того, что он был отложен в осадке правой стороной вниз, оставив левую сторону открытой для воздействия стихии. Он получил полевой номер MT-II, а позже номер экземпляра AGB 6257 в Аньхойском геологическом музее . [ 3 ]
описали этот образец В 2014 году Рёсуке Мотани и его коллеги в журнале Nature как представитель нового рода и вида Cartorhynchus lenticarpus . они получили Родовое название Cartorhynchus от греческих слов kartos (καρτός, «укороченный») и rhynchos (ῥύγχος, «морда»), а видовое название lenticarpus — от латинских слов lentus («гибкий») и carpus («запястье»). . Оба названия относятся к анатомическим характеристикам, которые он имел бы при жизни. [ 1 ] Считалось, что экземпляр был беззубым, пока во время дальнейших попыток удалить камень из сомкнутых челюстей не был обнаружен изолированный зуб. Поскольку образец был слишком хрупким, чтобы обнажить внутреннюю часть челюстей, Цзянь-Донг Хуан, Мотани и другие коллеги впоследствии отсканировали и визуализировали образец в 3D с помощью микрокомпьютерной томографии (микро-КТ), выполненной в испытательной компании Yinghua в Шанхай , Китай. В 2020 году результаты их последующей работы были опубликованы в журнале Scientific Reports . [ 3 ]
Описание
[ редактировать ]
На момент открытия Cartorhynchus был самым маленьким известным представителем Ichthyosauriformes . Сохранившийся экземпляр имел длину 21,4 сантиметра (8 дюймов); предполагая, что пропорции хвоста у него были сравнимы с близкими родственниками, Мотани и его коллеги оценили полную длину тела в 40 сантиметров (1 фут 4 дюйма) и вес в 2 килограмма (4,4 фунта). [ 1 ] [ 2 ] В 2021 году Сандер и его коллеги дали гораздо меньшую оценку веса — 237 г (8,4 унции). [ 4 ]
Череп
[ редактировать ]
У Карторинха была необычно короткая и суженная морда, занимавшая лишь половину длины черепа, и глубокая челюсть. Кончик морды имел ширину всего 6 миллиметров (0,2 дюйма). [ 3 ] В отличие от большинства рептилий, его носовая кость доходила до передней части морды. Из-за столь же удлиненной предчелюстной кости его костные ноздри располагались относительно далеко назад на черепе, а лобная кость также не имела расширения в заднем внешнем углу. Все эти характеристики были общими с его близким родственником Sclerocormus . Однако, в отличие от последнего, лобная кость не входила в глазницу у Cartorhynchus ; префронтальная и постфронтальная кости не сходятся над глазницей; [ 5 ] а расположение большого отверстия для шишковидной железы оно находилось на месте соприкосновения лобной и теменной костей на крыше черепа различалось: у Sclerocormus исключительно на теменных костях , а у Cartorhynchus . У Cartorhynchus также была характерно большая подъязычная кость . [ 1 ] [ 6 ]

Первоначально Мотани и его коллеги пришли к выводу, что Карторинх был беззубым; однако микро-КТ-сканирование впоследствии выявило наличие округлых зубов, похожих на коренные зубы, которые выступали внутрь почти перпендикулярно длинной оси челюсти, что делало их невидимыми снаружи. Все зубы были либо полностью уплощенными, либо слабо заостренными, многие зубы имели перетяжку между корнем и коронкой. В отличие от других ихтиозаврообразных с коренными зубами, все коронки зубов были «раздуты» в одинаковой степени. На верхней челюсти (верхняя челюсть) и зубной кости (нижняя челюсть) зубы располагались в три ряда, причем в крайнем ряду было больше всего и самых крупных зубов; на верхних челюстях было по семь, пять и, вероятно, по одному зубу, а на зубных – по десять, семь и четыре зуба. Среди ихтиозаврообразных только Cartorhynchus и Xinminosaurus имеют несколько рядов зубов. [ 7 ] Расположение зубов означало, что самые передние нижние зубы не имели соответствующего верхнего зуба, а также то, что две зубные кости, образующие нижнюю челюсть, не могли быть плотно слиты. Эта характеристика была свойственна его близким родственникам, беззубым хупесухиям . [ 3 ] [ 8 ]
Посткраниальный скелет
[ редактировать ]
У Cartorhynchus , по-видимому, было 5 шейных позвонков и 26 задних позвонков , всего 31 предкрестцовый позвонок ( позвонки перед крестцом или бедром). Наряду со Sclerocormus (с 34 пресакралями) и Chaohusaurus (с 36 пресакралями) Cartorhynchus попадает в типичный для наземных животных диапазон, в отличие от 40–80 пресакралей, распространенных среди более производных (специализированных) ихтиоптеригий . [ 1 ] В отличие от Sclerocormus , нервные отростки , выступающие из вершины позвонков у Cartorhynchus, были относительно узкими и наклонными, а не широкими и ребристыми. Cartorhynchus также можно отличить по парапофизам, позвоночным отросткам, сочленяющимся с ребрами; их передние края сливались с краями позвонков. [ 6 ]
И Cartorhynchus , и Sclerocormus имели массивные грудные клетки, наиболее глубокие у плеч, с широкими, уплощенными и толстостенными ребрами, что обычно наблюдается у ранних представителей вторично-водных рептилий. [ 1 ] [ 9 ] Ихтиоптеригии лишились этих уплощенных ребер, за исключением моллезавра . [ 10 ] На нижней стороне груди гастралии Cartorhynchus Sclerocormus были тонкими и палочковидными, в отличие от уплощенной «корзины» , но у обоих отсутствовала еще одна пара симметричных элементов на средней линии тела. [ 6 ]
Конечности Cartorhynchus были слабо окостеневшими (окостенели только три пальца руки) с широко расставленными костями, особенно между костями запястья (запястья) и пальцами, что позволяет предположить наличие обширного хряща в конечностях. Это сделало бы конечности похожими на плавники. Ласты передних конечностей Cartorhynchus были изогнуты назад, при этом пальцы были наклонены на 50° относительно оси длинных костей (zeugopodium), а задние конечности были изогнуты вперед. Бедренная кость Cartorhynchus . была прямой и не расширенной на нижнем конце У Sclerocormus были аналогичные конечности, за исключением того, что они были лучше окостеневшими, а их сохранившаяся кривизна могла быть неестественной. [ 1 ] [ 6 ]
Классификация
[ редактировать ]
Отсутствие полных ископаемых останков привело к отсутствию ясности в происхождении ихтиоптеригий, включая ихтиозавров . В течение многих лет считалось, что их окаменелости внезапно появились в среднем триасе и подверглись сильной водной адаптации. Открытие Cartorhynchus и Sclerocormus частично восполнило этот пробел. Филогенетический анализ, проведенный Мотани и его коллегами, показал, что эти два вида были тесно связаны друг с другом, образуя кладу (группу) под названием Nasorostra , а также с ихтиоптеригией, для которой назоространы образовали сестринскую группу . Cartorhynchus и Sclerocormus объединяли короткие морды, удлиненные носовые кости, глубокие челюсти, отсутствие расширений лобных костей, самая глубокая грудная клетка возле плеча и лопатки (лопатки), более широкие на нижнем конце, чем на верхнем. [ 1 ] [ 6 ]
Включение назоростран в филогенетический анализ также предоставило доказательства в пользу того, что хупезухии являются близкими родственниками ихтиоптеригий. В 2014 году Мотани и его коллеги назвали кладу, образованную Nasorostra и Ichthyopterygia, Ichthyosauriformes, а кладу, образованную Ichthyosauriformes и Hupehsuchia, — Ichthyosauromorpha . Примечательно, что тесная связь между этими различными группами была обнаружена в ходе их анализа независимо от того, были ли исключены из анализа характеристики, связанные с водной адаптацией. [ 1 ] [ 6 ] Такие характеристики могли развиться гомопластически (в результате конвергентной эволюции ) среди нескольких линий из-за схожего образа жизни, что может привести к искажению филогенетического анализа, чтобы реконструировать их как гомологии (полученные на основе общего происхождения). Сохранение этих реконструированных связей даже после удаления водных характеристик указывает на их устойчивость. [ 11 ] [ 12 ]
Ниже частично воспроизведено филогенетическое дерево из филогенетического анализа, опубликованного Хуангом, Мотани и коллегами в 2020 году в описании Chaohusaurus brevifemoralis : [ 3 ]
| Ихтиозавроморфа |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Эволюционная история
[ редактировать ]Появление ихтиозавроморф было частью восстановления морских экосистем после разрушительного пермско-триасового вымирания . Считалось, что морские экосистемы восстановили свое полное разнообразие только через 5–10 миллионов лет после массового вымирания и что морские рептилии восстанавливались после вымирания медленнее, чем другие линии. [ 13 ] [ 14 ] Однако открытие множества разнообразных фаун морских рептилий, встречавшихся в раннем триасе, включая Cartorhynchus , впоследствии показало, что это не так. [ 8 ] [ 15 ] В частности, по-видимому, ихтиозавроформы впервые появились в спатском подэтапе оленекского яруса и быстро достигли высокого функционального разнообразия в первый миллион лет своей эволюции. [ 6 ] они заняли самые разнообразные ниши . Несмотря на относительно низкое видовое разнообразие, [ 16 ] включая как демерсальные (донные) виды, такие как хупезухии и назоространы, так и пелагические (открытые водные) виды, такие как ихтиоптеригии. [ 6 ]

Многие ихтиозавроформы из раннего и среднего триаса имели молариформные зубы (показаны на филогенетическом дереве выше), включая Cartorhynchus ; такие зубы указывают на диету, по крайней мере частично основанную на животных с твердым панцирем. [ 17 ] В 2020 году Хуанг и его коллеги провели реконструкцию наследственного состояния зубов ихтиозавроморфов. Вероятностные методы показали, что округлые или плоские зубы, скорее всего, эволюционировали независимо пять раз, тогда как методы, основанные на экономии, предположили, что они эволюционировали независимо три-пять раз. Хуанг и его коллеги заметили, что развитие молариформных зубов много раз происходило независимо у водных животных (включая многочисленные линии варанов , мурен , спаридов и цихлид ), и поэтому частота среди ихтиозаврообразных не является необычно высокой. Они также заметили, что ихтиозавроформы раннего триаса обычно имели маленькие закругленные зубы; зубы ихтиозаврообразных среднего триаса были более разнообразными по размеру и форме, что коррелирует с увеличением разнообразия беспозвоночных. [ 18 ] Таким образом, они предположили, что диверсификация ихтиозавроформ частично была обусловлена эволюцией добычи с твердым панцирем. [ 3 ]
Однако хупезухии и назоространы в конечном итоге вымерли на границе раннего и среднего триаса, создав «таксономическое узкое место». На границе уровень моря меняется. [ 19 ] и вулканизм [ 20 ] привело к тому, что океаны стали плохо насыщены кислородом, что привело к появлению характерных изотопов углерода из разлагающегося органического материала в пластах горных пород на границе. [ 21 ] Разнообразие ихтиозаврообразных после этого оборота не восстановилось. [ 22 ] при этом Sauropterygia и Saurosphagidae вызвали вторую волну диверсификации, продолжавшуюся от трех до пяти миллионов лет. [ 6 ] [ 16 ]
Палеобиология
[ редактировать ]Диета
[ редактировать ]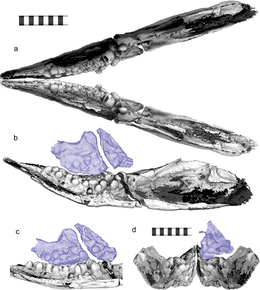
В 2014 году Мотани и его коллеги выдвинули гипотезу, что Cartorhynchus питался всасыванием , питаясь за счет концентрации давления в своей узкой морде. Это также подтверждалось прочностью его подъязычной кости и гиобранхиального элемента (который должен был закреплять язык), а также неправильным наблюдением отсутствия зубов. [ 1 ] Подобные выводы впоследствии были сделаны и для Sclerocormus . [ 6 ] Шастазавр и Шонизавр ранее интерпретировались как ихтиозавры, питающиеся всасыванием. [ 23 ] но количественный анализ триасовых и юрских ихтиозавров, проведенный Мотани и его коллегами в 2013 году, показал, что ни у одного из них не было достаточно крепких жаберных костей и достаточно узкой морды, чтобы можно было питаться всасыванием. [ 24 ]
Открытие молариформных зубов у Cartorhynchus привело Хуана и его коллег в 2020 году к выводу, что Cartorhynchus был дурофагом и питался добычей с твердым панцирем. Они отметили, что это не противоречит сосательному образу жизни; некоторые спариды одновременно питаются дурофагами и питаются всасыванием. [ 25 ] Однако они предположили, что Cartorhynchus мог питаться только мелкой добычей. Что касается горизонтальной ориентации зубов, они наблюдали поверхности износа, что указывало на то, что стороны зубов смыкались друг с другом, а не с коронками. Однако они отметили, что зубы структурно прочнее на кончиках, а не по бокам, в условиях высоких нагрузок от дробящего укуса. [ 26 ] Они предположили, что челюсть могла быть искривлена во время консервации или что мягкие ткани, такие как коллаген , которые удерживали зубы при жизни, могли быть потеряны, хотя они признали, что ни одна из гипотез не может объяснить характер износа. Наконец, они отметили, что нижние зубы без соответствующих верхних зубов были необычными; на них нет следов износа, и нет никаких свидетельств наличия мышечных механизмов, которые позволили бы использовать две челюсти друг против друга. Таким образом, они пришли к выводу, что эти зубы, вероятно, не использовались против других зубов. [ 3 ]
Конечности и передвижение
[ редактировать ]
У Cartorhynchus были плохо окостеневшие конечности, несмотря на хорошо окостеневшие череп и позвонки. Однако в 2014 году Мотани и его коллеги предположили, что это все еще взрослая особь, поскольку многие представители линий морских рептилий, рано расходящихся, имеют плохо окостеневшие конечности из-за педоморфоза (сохранения незрелых признаков во взрослом возрасте), хотя они не полностью отвергли возможность того, что это была молодь из-за существования только одного экземпляра. [ 1 ]
В случае с Cartorhynchus Мотани и его коллеги предположили, что большие, похожие на ласты передние конечности позволили бы ему передвигаться по суше, что сделало его амфибией. Обширный хрящ в лучезапястном суставе позволил бы плавнику сгибаться без локтя; У молодых морских черепах есть такие же хрящевые ласты, которые они используют для передвижения по суше. [ 27 ] Хотя его плавник не был особенно сильным, Карторинх был относительно легким, с соотношением массы тела к площади поверхности плавника меньшим, чем у Chaohusaurus . Изогнутые ласты позволили бы им держаться близко к телу, увеличивая тем самым их механическое преимущество . Мотани и его коллеги предположили, что другие черты Cartorhynchus также способствовали амфибийному образу жизни, в том числе короткий туловище и морда, а также утолщенные ребра (которые служили бы балластом , стабилизирующим животное в прибрежных водах). [ 1 ]

В исследовании 2019 года, проведенном Сусаной Гутаррой и ее коллегами, использовалось компьютерное моделирование для оценки энергетических затрат на плавание ихтиозаврообразных. Раннерасходящиеся ихтиозавроформы с телом в форме ящерицы и удлиненным хвостом без плавников, такие как Cartorhynchus , использовали бы угловатое (угреобразное) плавание, в то время как более поздние ихтиозавроформы с более глубокими рыбоподобными телами и четко выраженными хвостовыми плавниками использовали бы карангиформное плавание. скумбрияобразная) плавание. Принято считать, что ангилиформное плавание менее эффективно, чем карангиформное. [ 28 ] [ 29 ] [ 30 ] были в 24–42 раза выше, Действительно, Гутарра и его коллеги обнаружили, что энергетические затраты на плавание со скоростью 1 метр в секунду (3,3 фута/с) у Cartorhynchus чем у ихтиозавра Ophthalmosaurus (в зависимости от того, учитывается ли режим плавания), а его коэффициент сопротивления была на 15% выше, чем у афалин . Однако Cartorhynchus , вероятно, плавал на более медленных скоростях, что требовало меньшей эффективности, а преимущества карангиформного плавания среди более поздних, более крупных ихтиозавров также нивелировались увеличенным размером тела. [ 31 ]
Палеоэкология
[ редактировать ]
Слой 633 карьера Маджиашань, место, где был обнаружен Cartorhynchus , представляет собой слой серого глинистого (глинистосодержащего) известняка, расположенный на высоте 13 метров (43 фута) над основанием верхней пачки формации Нанлинху. Сверху и снизу он ограничен слоями желтоватых мергелей . По аммонитов биостратиграфии этот пласт относится к зоне Subcolumbites . [ 1 ] [ 32 ] Оценки дат с высоким разрешением были произведены для слоев оленекского возраста, обнаженных в карьере Маджиашань, на основе изотопных записей углерода-13 цикла и спектральных гамма- каротажей (которые измеряют количество радиации в породах астрономического происхождения); В частности, возраст слоя 633 оценивается в 248,41 млн лет назад. [ 33 ] [ 34 ] [ 35 ] В течение среднего триаса толщи Чаоху отлагались в сравнительно удаленной от побережья океанической котловине , окаймленной на юге более мелководными и карбонатными платформами , а на севере — континентальным склоном и более глубокими впадинами. [ 36 ] [ 37 ]
Ихтиозавроформы Sclerocormus и Chaohusaurus обнаружены в карьере Маджиашань вместе с Cartorhynchus ; Sclerocormus известен из более молодого слоя 719 (248,16 млн лет назад), а Chaohusaurus встречается в обоих слоях. [ 33 ] Зауроптериг Majiashanosaurus известен из слоя 643. [ 38 ] Рыбное разнообразие в карьере Мацзяшань беднее, чем в других местах; наиболее распространенной рыбой является Chaohuperleidus , старейший известный представитель Perleidiformes , вид широко распространенного Saurichthys и несколько неописанных рыб. но известны также [ 39 ] [ 40 ] Потенциальной добычей беспозвоночных для Cartorhynchus являются мелкие аммониты и двустворчатые моллюски , а также тилакоцефальные членистоногие Ankitokazocaris . [ 3 ] [ 32 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л м Мотани, Р.; Цзян, Д.-Ю.; Чен, Г.-Б.; Тинтори, А.; Риппель, О.; Джи, К.; Хуанг, Ж.-Д. (2014). «Базальный ихтиозаврообразный вид с короткой мордой из нижнего триаса Китая» . Природа . 517 (7535): 485–488. Бибкод : 2015Natur.517..485M . дои : 10.1038/nature13866 . ПМИД 25383536 . S2CID 4392798 .
- ^ Jump up to: а б Перкинс, Сид (2014). «Как у ихтиозавра появились плавники» . Наука . Проверено 8 мая 2020 г.
- ^ Jump up to: а б с д и ж г час Хуанг, J.-D.; Мотани, Р.; Цзян, Д.-Ю.; Рен, Х.-Х.; Тинтори, А.; Риппель, О.; Чжоу, М.; Ху, Ю.-К.; Чжан, Р. (2020). «Повторяющаяся эволюция дурофагии во время радиации ихтиозавров после массового вымирания, на которую указывает скрытый прикус» . Научные отчеты . 10 (1): 7798. Бибкод : 2020NatSR..10.7798H . дои : 10.1038/s41598-020-64854-z . ПМК 7210957 . ПМИД 32385319 .
- ^ Сандер, премьер-министр; Грибелер, Э.М.; Кляйн, Н.; Хуарбе, СП; Винтрич, Т.; Ревелл, LJ; Шмитц, Л. (2021). «Ранний гигант обнаруживает более быструю эволюцию крупных размеров тела у ихтиозавров, чем у китообразных». Наука . 374 (6575): eabf5787. дои : 10.1126/science.abf5787 . ПМИД 34941418 . S2CID 245444783 .
- ^ Чжоу, М.; Цзян, Д.-Ю.; Мотани, Р.; Тинтори, А.; Джи, К.; Вс, З.-Я.; Ни, П.-Г.; Лу, Х. (2017). «Краниальная остеология, обнаруженная на трехмерно сохранившихся черепах ихтиозавриформ раннего триаса Chaohusaurus chaoxianensis (Reptilia: Ichthyosauromorpha) из Аньхой, Китай». Журнал палеонтологии позвоночных . 37 (4): e1343831. дои : 10.1080/02724634.2017.1343831 . S2CID 90332288 .
- ^ Jump up to: а б с д и ж г час я дж Цзян, Д.-Ю.; Мотани, Р.; Хуанг, J.-D.; Тинтори, А.; Ху, Ю.-К.; Риппель, О.; Фрейзер, Северная Каролина; Джи, К.; Келли, штат Нью-Йорк; Фу, В.-Л.; Чжан, Р. (2016). «Большой аберрантный стебель ихтиозавроморфов, указывающий на ранний расцвет и упадок ихтиозавроморфов после вымирания в конце пермского периода» . Научные отчеты . 6 : 26232. Бибкод : 2016NatSR...626232J . дои : 10.1038/srep26232 . ПМЦ 4876504 . ПМИД 27211319 .
- ^ Цзян, Д.; Мотани, Р.; Хао, В.; Шмитц, Л.; Риппель, О.; Сан, Ю.; Сан, З. (2008). «Новый примитивный ихтиозавр (Reptilia, Diapsida) из среднего триаса Паньсяна, Гуйчжоу, юго-западного Китая и его место в триасовом биотическом восстановлении» . Прогресс в естествознании . 18 (10): 1315–1319. дои : 10.1016/j.pnsc.2008.01.039 .
- ^ Jump up to: а б Мотани, Р.; Чен, XH; Цзян, Д.-Ю.; Ченг, Л.; Тинтори, А.; Риппель, О. (2015). «Питание ранних морских рептилий и быстрая эволюция гильдий, питающихся морскими четвероногими» . Научные отчеты . 5 : 8900. Бибкод : 2015NatSR...5E8900M . дои : 10.1038/srep08900 . ПМК 4354009 . ПМИД 25754468 .
- ^ Хусай, А. (2009). « Пахиостоз» у водных амниот: обзор» . Интегративная зоология . 4 (4): 325–340. дои : 10.1111/j.1749-4877.2009.00146.x . ПМИД 21392306 .
- ^ Талеви, М.; Фернандес, MS (2012). «Неожиданная гистология скелета ихтиозавра из средней юры Патагонии: значение для эволюции микроструктуры костей среди вторичных водных четвероногих» . Naturwissenschaften . 99 (3): 241–244. Бибкод : 2012NW.....99..241T . дои : 10.1007/s00114-012-0886-4 . ПМИД 22290413 . S2CID 2223683 .
- ^ Чен, Х.-Х.; Мотани, Р.; Ченг, Л.; Цзян, Д.-Ю.; Риппель, О. (2014). «Загадочная морская рептилия нанчангозавр из нижнего триаса провинции Хубэй, Китай, и филогенетическое сходство Hupehsuchia» . ПЛОС ОДИН . 9 (7): e102361. Бибкод : 2014PLoSO...9j2361C . дои : 10.1371/journal.pone.0102361 . ПМК 4094528 . ПМИД 25014493 .
- ^ Мотани, Р.; Цзян, Д.-Ю.; Тинтори, А.; Риппель, О.; Чен, Г.-Б.; Ты, Х. (2015). «Первые свидетельства наличия централий у ихтиоптеригий, подтверждающие предвзятость педоморфных признаков при филогенетической реконструкции морских рептилий». Журнал палеонтологии позвоночных . 35 (4): e948547. дои : 10.1080/02724634.2014.948547 . S2CID 83576278 .
- ^ Чен, ZQ; Бентон, MJ (2012). «Время и характер восстановления биотики после массового вымирания в конце пермского периода». Природа Геонауки . 5 (6): 375–383. Бибкод : 2012NatGe...5..375C . дои : 10.1038/ngeo1475 .
- ^ Лю, Дж.; Ху, SX; Риппель, О.; Цзян, Д.-Ю.; Бентон, MJ; Келли, штат Нью-Йорк; Эйчисон, Дж. К.; Чжоу, К.; Вэнь, В.; Хуанг, Дж.; Се, Т.; Льв, Т. (2014). «Гигантский нотозавр (Reptilia: Sauropterygia) из среднего триаса на юго-западе Китая и его значение для триасового восстановления биотики» . Научные отчеты . 4 : 7142. Бибкод : 2014NatSR...4E7142L . дои : 10.1038/srep07142 . ПМЦ 4245812 . ПМИД 25429609 .
- ^ Бентон, MJ; Чжан, К.; Ху, С.; Чен, З.-Ц.; Вэнь, В.; Лю, Дж.; Хуанг, Дж.; Чжоу, К.; Се, Т.; Тонг, Дж.; Чу, Б. (2013). «Исключительная биота позвоночных триаса Китая и расширение морских экосистем после массового пермо-триасового вымирания». Обзоры наук о Земле . 125 : 199–243. Бибкод : 2013ESRv..125..199B . doi : 10.1016/j.earscirev.2013.05.014 .
- ^ Jump up to: а б Стаббс, ТЛ; Бентон, MJ (2016). «Экоморфологическое разнообразие мезозойских морских рептилий: роль экологических возможностей и вымирания» . Палеобиология . 42 (4): 547–573. дои : 10.1017/pab.2016.15 . hdl : 1983/c3f9b0fe-6bc2-400c-a770-1d1ab6e8379e . S2CID 3722056 .
- ^ Келли, штат Нью-Йорк; Мотани, Р. (2015). «Трофическая конвергенция стимулирует морфологическую конвергенцию морских четвероногих» . Письма по биологии . 11 (1): 20140709. doi : 10.1098/rsbl.2014.0709 . ПМЦ 4321144 . ПМИД 25631228 .
- ^ Фан, Ж.-Х.; Шен, С.-З.; Эрвин, Д.Х.; Сэдлер, премьер-министр; Маклауд, Н.; Ченг, Ц.-М.; Хоу, X.-D.; Ян, Дж.; Ван, X.-D.; Ван, Ю.; Чжан, Х.; Чен, X.; Ли, Г.-Х.; Чжан, Ю.-К.; Ши, Ю.-К.; Юань, Д.-Х.; Чен, К.; Чжан, Л.-Н.; Ли, К.; Чжао, Ю.-Ю. (2020). «Краткая информация о биоразнообразии морских беспозвоночных от кембрия до раннего триаса» . Наука . 367 (6475): 272–277. Бибкод : 2020Sci...367..272F . doi : 10.1126/science.aax4953 . ПМИД 31949075 . S2CID 210698603 .
- ^ Эмбри, А.Ф. (1997). «Глобальные границы последовательности триаса и их идентификация в осадочном бассейне Западной Канады» . Бюллетень канадской нефтяной геологии . 45 (4): 415–433.
- ^ Патон, Монтана; Иванов А.В.; Фиорентини, ML; Макнотон, Нью-Джерси; Мудровская И.; Резницкий Л.З.; Демонтерова, Е.И. (2010). «Позднепермские и раннетриасовые магматические импульсы в Ангаро-Тасеевой синклинали, Южно-Сибирских траппах и их возможное влияние на окружающую среду». Российская геология и геофизика . 51 (9): 1012–1020. Бибкод : 2010РуГГ...51.1012П . дои : 10.1016/j.rgg.2010.08.009 .
- ^ Такахаши, С.; Ямасаки, СИ; Огава, К.; Кайхо, К.; Цучия, Н. (2015). «Окислительно-восстановительные условия в Панталассе конца раннего триаса». Палеогеография, Палеоклиматология, Палеоэкология . 432 : 15–28. Бибкод : 2015PPP...432...15T . дои : 10.1016/j.palaeo.2015.04.018 .
- ^ Дик, генеральный директор; Максвелл, Э.Э. (2015). «Эволюция и вымирание ихтиозавров с точки зрения количественного экокосмического моделирования» . Письма по биологии . 11 (7): 20150339. doi : 10.1098/rsbl.2015.0339 . ПМЦ 4528449 . ПМИД 26156130 .
- ^ Сандер, премьер-министр; Чен, X.; Ченг, Л.; Ван, X. (2011). «Короткорылый беззубый ихтиозавр из Китая предполагает позднетриасовую диверсификацию ихтиозавров, питающихся всасыванием» . ПЛОС ОДИН . 6 (5): e19480. Бибкод : 2011PLoSO...619480S . дои : 10.1371/journal.pone.0019480 . ПМК 3100301 . ПМИД 21625429 .
- ^ Мотани, Р.; Джи, К.; Томита, Т.; Келли, Н.; Максвелл, Э.; Цзян, Д.-Ю.; Сандер, премьер-министр (2013). «Отсутствие всасывающего питания ихтиозавров и его последствия для триасовой мезопелагической палеоэкологии» . ПЛОС ОДИН . 8 (12): е66075. Бибкод : 2013PLoSO...866075M . дои : 10.1371/journal.pone.0066075 . ПМЦ 3859474 . ПМИД 24348983 .
- ^ Вандевалле, П.; Сентен, П.; Шардон, М. (1995). «Структуры и движения щечных и глоточных челюстей в связи с питанием Diplodus sargus ». Журнал биологии рыб . 46 (4): 623–656. дои : 10.1111/j.1095-8649.1995.tb01101.x . hdl : 2268/249857 .
- ^ Крофтс, SB; Саммерс, AP (2014). «Как лучше разбить улитку: влияние формы зуба на раздавливающую нагрузку» . Журнал интерфейса Королевского общества . 11 (92): 20131053. doi : 10.1098/rsif.2013.1053 . ПМЦ 3899874 . ПМИД 24430124 .
- ^ Мазухова Н.; Умбанховар, ПБ; Гольдман, Д.И. (2013). «Наземное передвижение робота, напоминающего морскую черепаху, с помощью ласт». Биоинспирация и биомиметика . 8 (2): 026007. Бибкод : 2013BiBi....8b6007M . дои : 10.1088/1748-3182/8/2/026007 . ПМИД 23612858 . S2CID 14888521 .
- ^ Фиш, FE (2000). «Биомеханика и энергетика водных и околоводных млекопитающих: от утконоса до кита». Физиологическая и биохимическая зоология . 73 (6): 683–698. дои : 10.1086/318108 . ПМИД 11121343 . S2CID 49732160 .
- ^ Тителл, Эд; Боражжани, И.; Сотиропулос, Ф.; Бейкер, ТВ; Андерсон, Э.Дж.; Лаудер, Г.В. (2010). «Распутывание функциональной роли морфологии и движения при плавании рыб» . Интегративная и сравнительная биология . 50 (6): 1140–1154. дои : 10.1093/icb/icq057 . ПМЦ 2981590 . ПМИД 21082068 .
- ^ Боражжани, И.; Сотиропулос, Ф. (2010). «О роли формы и кинематики в гидродинамике плавания самоходного корпуса/хвостового плавника» . Журнал экспериментальной биологии . 213 (1): 89–107. дои : 10.1242/jeb.030932 . ПМИД 20008366 . S2CID 27365252 .
- ^ Гутарра, С.; Мун, Британская Колумбия; Рахман, Айова; Палмер, К.; Лаутеншлагер, С.; Бримакомб, Эй Джей; Бентон, MJ (2019). «Влияние эволюции плана тела на гидродинамическое сопротивление и энергетические потребности ихтиозавров при плавании» . Труды Королевского общества Б. 286 (1898): 20182786. doi : 10.1098/rspb.2018.2786 . ПМК 6458325 . ПМИД 30836867 .
- ^ Jump up to: а б Джи, К.; Тинтори, А.; Цзян, Д.; Мотани, Р. (2017). «Новые виды Thylacocephala (Arthropoda) из спата (нижний триас) Чаоху, провинция Аньхой, Китай» ПалЗ . 91 (2): 171–184. дои : 10.1007/ s12542-017-0347-7 S2CID 132045789 .
- ^ Jump up to: а б Мотани, Р.; Цзян, Д.-Ю.; Тинтори, А.; Джи, К.; Хуанг, Ж.-Д. (2017). «Дивергенция мезозойских морских рептилий до и после массового вымирания, продиктованная зависимостью темпов эволюции от временного масштаба» . Труды Королевского общества Б. 284 (1854): 20170241. doi : 10.1098/rspb.2017.0241 . ПМЦ 5443947 . ПМИД 28515201 .
- ^ Фу, В.; Цзян, Д.; Монтаньес, И.; Мейлерс, С.; Мотани, Р.; Тинтори, А. (2016). «Эксцентриситет и наклонность темпов углеродного цикла в раннем триасе и последствия для восстановления экосистем после вымирания» . Научные отчеты . 6 : 27793. Бибкод : 2016NatSR...627793F . дои : 10.1038/srep27793 . ПМЦ 4904238 . ПМИД 27292969 .
- ^ Ли, М.; Огг, Дж.; Чжан, Ю.; Хуанг, К.; Хиннов, Л.; Чен, ZQ; Цзоу, З. (2016). «Астрономическая настройка конца пермского вымирания и эпохи раннего триаса Южного Китая и Германии». Письма о Земле и планетологии . 441 : 10–25. Бибкод : 2016E&PSL.441...10L . дои : 10.1016/j.epsl.2016.02.017 .
- ^ Джи, К.; Чжан, К.; Цзян, Д.Ю.; Бучер, Х.; Мотани, Р.; Тинтори, А. (2015). «Контроль возраста аммоноидей морских рептилий раннего триаса из Чаоху (Южный Китай)». Палеомир . 24 (3): 277–282. дои : 10.1016/j.palwor.2014.11.009 .
- ^ Ли, С.Ю.; Тонг, Дж. Н.; Лю, Кентукки; Ван, Ф.Дж.; Хо, ГГ (2007). «Циклическое отложение нижнего триаса в Чаоху, провинция Аньхой, Китай». Палеогеография, Палеоклиматология, Палеоэкология . 252 (1–2): 188–199. Бибкод : 2007PPP...252..188S . дои : 10.1016/j.palaeo.2006.11.043 .
- ^ Цзян, Д.-Ю.; Мотани, Р.; Тинтори, А.; Риппель, О.; Чен, Г.-Б.; Хуанг, J.-D.; Чжан, Р.; Вс, З.-Я.; Джи, К. (2014). «Ранний триасовый эозавроптериг Majiashanosaurus diskcocoracoidis , gen. et sp. nov. (Reptilia, Sauropterygia), из Чаоху, провинция Аньхой, Китайская Народная Республика». Журнал палеонтологии позвоночных . 34 (5): 1044–1052. дои : 10.1080/02724634.2014.846264 . S2CID 129828376 .
- ^ Солнце, З.; Тинтори, А.; Цзян, Д.; Мотани, Р. (2013). «Новый перлейдид из спатия (оленекского яруса, раннего триаса) Чаоху, провинция Аньхой, Китай». Rivista Italiana di Paleontologia e Stratigrafia . 119 (3): 275–285. дои : 10.13130/2039-4942/6040 .
- ^ Тинтори, А.; Хуанг, Ж.-Д. Цзян, Д.-Ю.; Вс, З.-Я.; Мотани, Р.; Чен, Г. (2014). «Новый Saurichthys (Actinopterygii) из спатия (ранний триас) Чаоху (провинция Аньхой, Китай)» Итальянский журнал палеонтологии и стратиграфии . 120 (2): 157–164. дои : 10.13130/2039-4942/6057 .







