Грапталит
| Грапталиты Временный диапазон:
Выживает в настоящее время через живой род Rhabdopleura . [ 2 ]
| |
|---|---|

| |
| Cryptograptus из силурийской Южной Америки. Образец в Музее Королевского Онтарио | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Hemichordata |
| Сорт: | Птеробранчия |
| Подкласс: | Graptolithina Бронн, 1849 |
| Подгруппы | |
| |
Грапталиты - это группа колониальных животных , члены подкласса Graptolithina в классе Pterobranchia . Эти организмы , кормящие фильтров, известны главным образом из окаменелостей , найденных из среднего кембрия ( Миолингиан , Вулиан ) через нижний каменноугольный ( Миссисипский ). [ 3 ] Возможный ранний грапталит, Chaunograptus , известен из среднего кембрия. [ 1 ] Недавние анализы предположили идею о том, что живой Pterobranch Rhabdopleura представляет собой существующий граптолит, который расходился от остальной части группы в кембрийском языке. [ 2 ]
Отопаемые граптолиты и Rhabdopleura имеют структуру колонии взаимосвязанных зооидов, размещенных в органических трубах (THECA), которые имеют базовую структуру сложенных полузащитных колец (Fuselli). Большинство потушенных граптолитов принадлежат к двум основным порядкам: похожим на кустах, сидячими дендроидами и планктонной , свободно плавающей отгрузкой . Эти заказы, скорее всего, развивались из инкрустирующих птеробранч, аналогичных Rhabdopleura . Из-за их широко распространенного изобилия, планктонного образа жизни и хорошо пролеченных эволюционных тенденций, в частности, граптолоиды являются полезными индексными окаменелостями для ордовикских и силурских периодов. [ 4 ]
Название Graptolite происходит от греческого graptos, означающего «написанный», и литос, означающий «рок», так как многие окаменелости для граптолитов напоминают иероглифы, написанные на скале. Линнеус изначально рассматривал их как « картины, напоминающие окаменелости », а не истинные окаменелости, хотя более поздние работники предполагали, что они связаны с гидрозоанами ; Теперь они широко признаны полуходатами . [ 4 ]
История
[ редактировать ]Название «Graptolite» происходит от рода Graptolilithus («Написание на скалах»), который использовался Линнеусом в 1735 году для неорганических минерализаций и индукций, которые напоминали фактические окаменелости. В 1768 году в 12 -м объеме Systema Naturae он включил G. Sagittarius и G. Scalaris , соответственно возможный ископаемый растение и возможный граптолит. В своем 1751 году Skånska Resa он включил фигуру «ископаемого или граптолита странного вида», который, как считается, является типом климакограпта (род бизеральных граптолитов).
Окаменелости для граптолитов позже были направлены на различные группы, в том числе другие разветвленные колониальные животные, такие как мжзильные жители («Мосские животные») и гидрозоанские . Термин Graptolithina был создан Бронном в 1849 году, который считал их представлять ортоконные головоногих. К середине 20-го века граптолиты были признаны уникальной группой, тесно связанной с живыми птеробранками в родах Rhabdopleura и Cephalodiscus , которые были описаны в конце 19-го века. Graptolithus , как род, был официально заброшен в 1954 году ICZN . [ 5 ]
Морфология
[ редактировать ]Структура колонии
[ редактировать ]
Каждая граптолитная колония происходит от начального человека, называемого Sucult Zooid, из которого будут развиваться последующие зооиды . Все они взаимосвязаны Столонами , настоящей колониальной системой, разделяемой Rhabdopleura , но не головокружением . Эти зооиды размещены в органической структуре, содержащей серию трубок, секретируемых железами на головозном щите . Структура колонии была известна из нескольких различных названий, включая коенециум (для живых птеробранков), рабдосомы (для ископаемых граптолитов) и чаще всего тубария (для обоих). Отдельные трубки, каждый из которых занят одним зооидом, известны как Theca . [ 4 ] Композиция тубария не известна, но разные авторы предполагают, что он сделан из коллагена или хитина . В некоторых колониях существует два размера Theca, более крупного автотеки и меньшего размера, и было высказано предположение, что эта разница связана с сексуальным диморфизмом зоопарков в колонии. [ 4 ]
В начале развития колонии труба расщепляется в переменное число ветвей (известных как Stypes ) и различные расположения Theca, особенности, которые важны для идентификации ископаемых граптолитов. Колонии могут быть классифицированы по их общему количеству рядов Theca (у бизеральных колоний есть два ряда, у универсального есть один), а количество начальных станов на колонию (многорамные колонии имеют много стип, у паухирамных колоний есть два или один). Каждая текальная трубка в основном состоит из двух серий сложенных полукруглых полусексов, известных как Fuselli (Sing: Fusellum). Фуселли напоминает линии роста при сохранении в окаменелостях, и эти два стека встречаются вдоль шва с зигзагообразным рисунком. Fuselli являются основным усиливающим компонентом трубки, хотя им помогает один или несколько дополнительных слоев более низкой ткани, коры.

Самые ранние граптолиты появились в отчете окаменелостей во время кембрий, и, как правило, были сидячими животными, с колонией, прикрепленной к морскому дну. Несколько ранних семейных семей были инкрустирующими организмами, а колония развивалась горизонтально вдоль субстрата. Экспадиционный рабдоплевра попадает в эту категорию, с общей инкрустирующей колонией в сочетании с вертикальной вертикальной тека. Большинство прямых, дендритных или густых/вентиляционных граптолитов классифицируются как денроиды (заказ dendroidea). Их колонии были привязаны к жесткому субстрату с собственным весом через диск привязанности. Грапталиты с относительно небольшим количеством ветвей были получены из граптолитов Дендроида в начале периода ордовика . Эта последняя крупная группа, граптолоиды (порядка graptololoidea) были пелагическими и планктоническими , свободно дрейфующими через толщу воды. Они были успешной и плодотворной группой, являющимися самыми важными и широко распространенными макропланктоническими животными, пока они не умерли в начале Девонский период. Граптолиты Дендроида выжили до каменного периода.
Зоологии
[ редактировать ]
Зрелый зооид имеет три важных региона, доральный диск или головное щит, воротник и туловище. В воротнике находятся рта и задний проход (U-образная пищеварительная система) и руки находятся; Graptholitina имеет одну пару рук с несколькими парными щупальцами. Как нервная система , грапталиты имеют простой слой волокон между эпидермисом и базальной пластинкой, также имеют ганглион воротника , который порождает несколько нервных ветвей, аналогичных нейронной трубке аккордов. [ 6 ] Правильные окаменелости мягких частей граптолитов еще не найдены, и неизвестно, были ли у них пленки с глоточной жабкой или нет, нет, нет, нет, нет, [ 7 ] Но, основываясь на существующей Rhabdopleura , вполне вероятно, что зооиды Grapotlite имели такую же морфологию. [ 4 ]
Таксономия
[ редактировать ]С 1970 -х годов, в результате достижений в области электронной микроскопии , как правило, граптолиты, как правило, наиболее тесно связаны с Pterobranchs , редкой группой современных морских животных, принадлежащих к филам Hemichordata . [ 8 ] Сравнения проводятся с современными гемиходатами цефалодиска и рабдоплевры . Согласно недавним филогенетическим исследованиям, рабдоплериды помещаются в граптолитину. Тем не менее, они считаются семейством Incertae Sedis . [ 3 ]
С другой стороны, Cephalodiscida считается сестринским подклассом Graptolithina. Одним из основных различий между этими двумя группами является то, что виды цефалодисциды не являются колониальными организмами. В организмах Cephalodiscida нет общего канала, соединяющего все зооиды. Зооиды Cephalodiscida имеют несколько рук, в то время как зооиды Graptolithina имеют только одну пару рук. Другие различия включают тип раннего развития, гонады, наличие или отсутствие прорезей жабры и размер зооидов. В записи ископаемых, где сохраняются в основном тубарию (трубки), сложно различать группы.
| Филогения птеробранчии [ 3 ] | |||
|
Graptolithina включает в себя несколько небольших семей, а также два основных вымерших порядка, Dendroidea ( бентические граптолиты) и Graptololoidea ( планктические граптолиты). Последнее является наиболее разнообразным, в том числе 5 подчиненных, где наиболее различенными являются аксонофора (бизерные граптолиты и т. Д.). Эта группа включает в себя диплограптидов и неограптидов , группы, которые имели большое развитие во время ордовика. [ 3 ] Старые таксономические классификации рассматривают приказы Dendroidea, Tuboidea, Camaroidea, Crustoidea, Stolonoidea, Graptoloidea и Dithecoidea, но новые классификации встроены в Graptoloidea на разных таксономических уровнях.
Таксономия Граптолитины Малец (2014): [ 3 ] [ 4 ]
Подкласс Graptolithina Bronn, 1849
- Неопределенность
- Семейство Rhabdopleuridae Harmer, 1905
- Семейство † Cysticamaridae Bulman, 1955
- Семейство † Wimanicrustidae Bulman, 1970
- Семейство † Dithecodendridae Obut, 1964
- Семейство † Cyclograptidae Bulman, 1938
- Заказ † Dendroidea Nicholson, 1872
- Семья † Дендрограптид Роверс, 1897 г. во Фриле, 1897
- Семейство † acanthograptidae bulman, 1938
- Семья мастигограф †
- Заказ † Graptoloidea Lapworth, 1875 в Hopkinson & Lapworth, 1875 (планктические граптолиты)
- Suborder † Graptodendroidina Mu & Lin, 1981 в Lin (1981)
- Семейство † anisograptidae Bulman, 1950
- Подстановка † Sinograpta Maletz et al. , 2009
- Семейство † Sigmagraptidae Cooper & Fortey, 1982
- Семья † Sinograptidae mu, 1957
- Семейство † Abrograptidae mu, 1958
- Подборка † Dichograptina Lapworth, 1873
- Семейство † Dichograptidae Lapworth, 1873
- Семейство † Didymograptidae mu, 1950
- Семейство † Pterograptidae mu, 1950
- Семейство † Tetragraptidae Frech, 1897
- Подборка † Glossograptina Jaanusson, 1960
- Семейство † isograptidae Harris, 1933
- Семейство † Glossograptidae lapworth, 1873
- Suborder † Axonophora Frech, 1897 (бизерные граптолиты, а также ретиолитиды и монограптиды)
- InfraRORD † Diplograptina Lapworth, 1880
- Семья † Diplograptidae Lapworth, 1873
- Подсемейство † Diplograptinae Lapworth, 1873
- Подсемейство † Orthograptinae Mitchell, 1987
- Семейство † lasiograptidae lapworth, 1880e
- Семейство † Climacograptidae Frech, 1897
- Семейство † dicranograptidae lapworth, 1873
- Подсемейство † dicranograptinae lapworth, 1873
- Подсемейство † nemagraptinae lapworth, 1873
- Семья † Diplograptidae Lapworth, 1873
- Infraorder † Neograptina Štorch et al. , 2011
- Семейство † Normalograptidae štorch & Serpagli, 1993
- Семейство † neodiplograptidae melchin et al. , 2011
- Подсемейство † neodiplograptinae melchin et al. , 2011
- Подсемейство † Petalolithinae Bulman, 1955
- Суперсемейство † retiolitoidea lapworth, 1873
- Семейство † retiolitidae lapworth, 1873
- Подсемейство † Retiolitinae Lapworth, 1873
- Подсемейство † Plectograptinae Bouček & Münch, 1952
- Семейство † retiolitidae lapworth, 1873
- Суперсемейство † Monograptoidea lapworth, 1873
- Семейство † Dimorphograptidae elles & Wood, 1908
- Семейство † Monograptidae Lapworth, 1873
- InfraRORD † Diplograptina Lapworth, 1880
- Suborder † Graptodendroidina Mu & Lin, 1981 в Lin (1981)
Экология
[ редактировать ]
Грапталиты были основным компонентом ранних палеозойских экосистем, особенно для зоопланктона , потому что наиболее распространенные и разнообразные виды были планктоническими. Грапталиты были, скорее всего, кормушки для подвески и напрягали воду для пищи, такие как планктон. [ 9 ]
Выводя вывод по аналогии с современными птеробранками, они смогли мигрировать вертикально через толщу воды для эффективности питания и избежать хищников. С помощью экологических моделей и исследований фаций было отмечено, что, по крайней мере, для видов ордовиков, некоторые группы видов в значительной степени ограничены эпипелагической и мезопелагической зоной, от прибрежного до открытого океана. [ 10 ] Живая рабдоплера была обнаружена в глубоких водах в нескольких регионах Европы и Америки, но распространение может быть предвзято за счет усилий по отбору отбора проб; Колонии обычно встречаются в виде эпибионтов снарядов.
Их локомоция была относительно массы воды, в которой они жили, но точные механизмы (такие как турбулентность, плавучесть , активное плавание и т. Д.) Еще не ясны. Одно предложение, выдвинутое Мелчином и Демонтом (1995), предположило, что движение грапталита было аналогично современным свободно плавающим животным с тяжелыми жилищными сооружениями. В частности, они сравнили граптолиты с «морскими бабочками» ( Thecostomata ), небольшими плавающими Pteropod улитками . Согласно этому предложению, граптолиты проходили через греблю или плавание посредством волнообразного движения парных мышечных придатков , разработанных из головокружительного щита или кормления щупальц. У некоторых видов текальная апертура, вероятно, была настолько ограничена, что гипотеза придатков невозможна. С другой стороны, плавучесть не поддерживается каким-либо дополнительным механизмом контроля тканей или газа, а активное плавание требует много энергетических отходов, которые, скорее, будут использованы для конструкции тубария. [ 10 ]
Есть еще много вопросов, касающихся передвижения грапталита, но все эти механизмы являются возможными альтернативами в зависимости от вида и его среды обитания. Для бентических видов, которые жили, прикрепленные к осадкам или любому другому организму, это не было проблемой; Зооиды были в состоянии двигаться, но ограничены в трубке. Хотя это движение зооиды возможно как у планктических, так и у бентических видов, оно ограничено Столоном , но особенно полезно для кормления. Используя свои руки и щупальца, которые близки к рту, они фильтруют воду, чтобы поймать любые частицы пищи. [ 10 ]
Жизненный цикл
[ редактировать ]Изучение биологии развития граптолитины было возможным благодаря открытию вида R. Compacta и R. Normani на мелководье; Предполагается, что окаменелости граптолитов имели аналогичное развитие, как и их существующие представители. Жизненный цикл включает в себя два события, онтогенью и астогеню, где основное различие состоит в том, происходит ли развитие в отдельном организме или в модульном росте колонии.
Жизненный цикл начинается с планктонной планктонную планула , похожей на личинки , произведенную сексуальным размножением, которая впоследствии становится Sucult Zooid , которая начинает колонию. В Rhabdopleura колонии несут мужчины и женские зооиды, но оплодотворенные яйца инкубируются в женской тубарии, и остаются там, пока они не станут личинками, способными плавать (через 4–7 дней), чтобы успокоиться, чтобы начать новую колонию. Каждая личинка окружает себя в защитном коконе, где происходит метаморфоз к зооиде (7–10 дней) и прикрепляется к задней части тела, где в конечном итоге развивается стебель. [ 4 ]
Развитие является косвенным и лецитотрофным , а личинки ресничны и пигментированы, с глубокой депрессией на вентральной стороне. [ 11 ] [ 6 ] Астогенью происходит, когда колония растет путем бесполого воспроизведения из кончика постоянного терминала зооида, за которым новые зооиды выделены из стебля, тип почковатого, называемого моноподиальным . Вполне возможно, что у окаменелостей граптолита терминальная зооида не была постоянной, потому что новые зооиды, образованные из кончика последней, другими словами, симпподиального начинающегося. Эти новые организмы разбивают дыру в стене тубария и начинают секретировать свою собственную трубку. [ 4 ]
Граптолиты в эволюционном развитии
[ редактировать ]
В последние годы живые граптолиты использовались в качестве модели гемихордата для исследований Evo-Devo , как и их сестринская группа, Acorn Worms . Например, граптолиты используются для изучения асимметрии в гемиходатах, особенно потому, что их гонады, как правило, расположены случайным образом с одной стороны. В Rhabdopleura Normani яичко и расположено асимметрично, и, возможно, другие структуры, такие как ламелла для полости рта гонопор . [ 12 ] Значение этих открытий состоит в том, чтобы понять раннюю асимметрию левого правого позвоночного из-за того, что аккорды являются родственной группой гемихордов, и, следовательно, асимметрия может быть особенностью, которая развивалась на ранней стадии дейтерии . Поскольку местоположение структур не установлено строго, также в некоторых энтеропнесах , вероятно, асимметричные состояния в полуходовцах не находятся в сильном развитии или эволюционном ограничении. На происхождение этой асимметрии, по крайней мере, для гонадов, возможно, влияет направление базального наращивания в тубарии некоторыми внутренними биологическими механизмами в птеробранках или исключительно факторами окружающей среды. [ 12 ]
Hedgehog (HH), высоко консервативный ген, вовлеченный в паттерн развития нейронного развития, был проанализирован в Hemichordates, принимая Rhabdopleura в качестве представителя Pterobranch. Было обнаружено, что ген ежа у Pterobranchs экспрессируется по -другому по сравнению с другими гемиходатами в качестве Enteropneust Saccoglossus Kowalevskii . изменяется в R. compacta путем введения аминокислотного треонина (t) в N-конце Важный консервативный мотив глицин-цистеин-фенилаланина (GCF) в месте автокаталитического расщепления в генах HH Существует замена серина (ы) для глицина (G). Эта мутация снижает эффективность аутопротеолитического расщепления и, следовательно, сигнальной функции белка. Не ясно, как этот уникальный механизм произошел в эволюции и влияния, которые он оказывает в группе, но, если он сохранялся в течение миллионов лет, он подразумевает функциональное и генетическое преимущество. [ 13 ]
Геологическая значимость
[ редактировать ]Сохранение
[ редактировать ]
Грапталиты являются обычными окаменелостями и имеют мировое распределение. Они чаще всего встречаются в сланцах и грязевых стадах, где окаменелости морского слоя редки, этот тип породы, образованный от отложений, откладываемых в относительно глубокой воде с плохим нижним циркуляцией, был дефицит в кислороде и не имел мусорщиков. Мертвые планктические граптические граптолиты, потопленные на морское дно, в конечном итоге стали бы захоронены в осадке и, таким образом, были хорошо сохранены.
Эти колониальные животные также обнаруживаются в известняках и бухте , но, как правило, эти скалы были отложены в условиях, которые были более благоприятными для жизни в дне, включая мусорщиков, и, несомненно, большинство останков граптолита здесь обычно съели другие животные.

Окаменелости часто обнаруживаются, сплющенные вдоль плоскости постельных принадлежностей камней, в которых они встречаются, хотя могут быть обнаружены в трех измерениях, когда они заполнены железным пиритом или другими минералами. Они варьируются по форме, но чаще всего являются викинтирующими: дендритные или ветвирующие (такие как диктионема ), пистса -похожие или « настройка вилки » (например, Didymograptus murchisoni ). Их останки могут быть приняты за ископаемые растения случайным наблюдателем, как это было в случае с первыми описаниями граптолитов.
Граптолиты обычно сохраняются как черная углеродная пленка на поверхности скалы или в виде светло -серых глиняных пленок в тектонически искаженных породах. Окаменелость также может показаться растянутым или искаженным. Это связано с слоями, в которых находится грапталит, сложенный и уплотненный. Иногда их может быть трудно увидеть, но, наклонив образец на свет, они показывают себя как блестящая маркировка. Пиритизированные граптолитовые окаменелости также обнаружены.
Известным местом для ископаемых граптолитов в Британии является залив Абердедди , Дайфед , Уэльс , где они встречаются в скалах с периода ордовика . Места в южных возвышенных районах Шотландии, границ озера и валлийских границ также дают богатые и хорошо сохранившиеся граптолитовые фауны. Знаменитое местоположение грапталита в Шотландии-это Линн DOB с видами из граничного ордовика-силурского. Поскольку группа имела широкое распространение, окаменелости также в нескольких частях Соединенных Штатов, Канады, Австралии, Германии и Китая, среди прочего.
Стратиграфия
[ редактировать ]Граптолитовые окаменелости имеют предсказуемое сохранение, широкое распространение и постепенное изменение по геологическим временным шкалам . Это позволяет им использоваться для датирования слоев камней по всему миру. [ 8 ] Они являются важными индексными окаменелостями для знакомства с палеозойскими породами, поскольку они быстро развивались со временем и образуют множество различных отличительных видов. Геологи могут разделить скалы ордовикских и силурийских периодов на биозоны грапталита; Обычно это составляет менее миллиона лет. Всемирный ледниковый период в конце ордовика устранял большинство граптолитов, кроме неограптинов. Диверсификация от неограптинов, которые пережили ордовикское оледенение, началась около 2 миллионов лет спустя. [ 14 ]
Великое событие биорадификации ордовиков ( GOBE ) повлияло на изменения в морфологии колоний и Thecae, что привело к новым группам, таким как планктическая GraptoloDiodea. Позже, некоторые из величайших вымираний, которые повлияли на группу, были гирнантианами в ордовике и Лундгрене в силурийском, где популяции граптолитов были резко уменьшены (см. Также эффект Лилипута ). [ 4 ] [ 15 ]
Разнообразие граптолитов было значительно сокращено во время события Sedgwickii в аэрониане . [ 16 ] Это событие было засвидетельствовано в таких местах, как сегодняшняя Канада, Ливия, а также в формировании Аргентины в Ла -Чилке (тогда часть Гондваны ). [ 16 ]
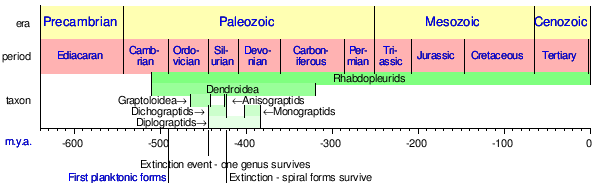 | |
| Диапазоны таксонов граптолита. | |
Исследователи
[ редактировать ]Ниже приведен выбор исследователей граптолита и птеробранша: [ 4 ]
- Иоахим Барранде (1799–1883)
- Ханнс Бруно Гейниц (1814–1900)
- Джеймс Холл (1811–1898)
- Фредерик М'Кей (1817–1899)
- Генри Эллейн Николсон (1844–1899)
- Джон Хопкинсон (1844–1919)
- Свен Леонхард Тёрнквист (1840–1920)
- Свен Аксель Туллберг (1852–1886)
- Герхард Холм (1853–1926)
- Карл Виман (1867–1944)
- Томас Сержант Холл (1858–1915)
- Александр Роберт Кейбл (1884–1963)
- Ноэль Бенсон (1885–1957)
- Уильям Джон Харрис (1886–1957)
- Дэвид Эван Томас (1902–1978)
- MU Enzhi (1917–1987)
- Ли Джиджин (1928–2013)
- Vladimir Nikolayevich Beklemishev (1890–1962)
- Майкл Сарс (1805–1869)
- Джордж Оссиан Сарс (1837–1927)
- Уильям Кармайкл М'Интош (1838-1931)
- Нэнси Кирк (1916–2005)
- Роман Козловски (1889–1977)
- Jörg Maletz
- Денис Эб Бейтс
- Альфред С. Ленц
- Крис Б. Кэмерон
- Адам Урбанек
- Дэвид К. Лойделл
- Герман Дэгер (1929–1992)
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а беременный Малец Дж. (2014). Hemichordata (Pterobranchia, Enteropneusta) и ископаемые записи. Палеогеография, палеоклиматология, палеоэкология , 398: 16-27.
- ^ Jump up to: а беременный Mitchell, CE, Melchin, MJ, Cameron, CB & Maletz, J. (2013) Филогенетический анализ показывает, что Rhabdopleura является существующим граптолитом. Lethaia , 46: 34–56.
- ^ Jump up to: а беременный в дюймовый и Малец, Джёрг (2014). «Классификация Pterobranchia (Cephalodiscida и Graptolithina)» . Бюллетень из героя . 89 (3): 477–540. doi : 10.3140/bull.geosci.1465 . ISSN 1214-1119 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж Mallet, Jörg (2017). Граплолит палеобиология . Wiley-Blackwell. ISBN 9781118515617 .
- ^ Bulman, M. (1970) в Teichert, C. (ed.). Трактат о палеонтологии беспозвоночных . Часть V. graptolithina, с разделами на Enteropneusta и Pterobranchia . (2 -е издание). Геологическое общество Америки и Университет Канзасской прессы, Боулдер, Колорадо и Лоуренс, Канзас, XXXII + 163 стр.
- ^ Jump up to: а беременный Сато А., Бишоп, Дж. И Холланд, П. (2008). Биология развития птеробранха полуходов: история и перспективы. Genesis , 46: 587-591.
- ^ Основы палеонтологии беспозвоночных: макрофоссили
- ^ Jump up to: а беременный Форти, Ричард А. (1998). Жизнь: естественная история первых четырех миллиардов лет жизни на земле . Нью -Йорк: Альфред А. Кнопф. п. 129
- ^ "Грапталиты" . Samnoblemuseum.ou.edu . Получено 2018-12-28 .
- ^ Jump up to: а беременный в Купер Р., Ригби С. , Лойделл Д. и Бейтс Д. (2012) Палеоэкология Graptoloidea. Земля-научные обзоры , 112 (1): 23-41.
- ^ Röttinger, E. & Lowe, C. (2012) Эволюционные перекрестки в биологии развития: полуходы. Разработка , 139: 2463-2475.
- ^ Jump up to: а беременный Sato, A. & Holland, P. (2008). Асимметрия в гемихордном доме птеробранке и эволюция паттерна левого правого. Динамика развития , 237: 3634 –3639)
- ^ Sato, A., White-Cooper, H., Doggett, K. & Holland, P. 2009. Вырожденная эволюция гена хеджихога в линии гемихордата. Труды Национальной академии наук , 106 (18): 7491-7494.
- ^ BAPST, D., Bullock, P., Melchin, M., Sheets, D. & Mitchell, C. (2012) Граптолоидное разнообразие и неравенство стали отделенными во время массового вымирания ордовика. Труды Национальной академии наук , 109 (9): 3428-3433.
- ^ Urbanek, Adam (1993). «Биотические кризисы в истории верхних силурийских граптолоидов: палеобиологическая модель». Историческая биология . 7 : 29–50. doi : 10.1080/10292389309380442 .
- ^ Jump up to: а беременный Лопес, Фернандо Энрике; Кауфманн, Синтия (2023). «Новое понимание силурской граптолитной биостратиграфии формирования La Chilca, секции Poblete Norte, центральных предварительных регистраций Сан -Хуана, Аргентина: замена фауны и палеоэкологические последствия» . Андская геология . 50 (2): 201. doi : 10.5027/andgeov50n2-3617 .
Внешние ссылки
[ редактировать ]- Классификация Graptolilithoidea - Граптолиты и Птеробранки
- Подкаст на граптолитах Дэвида Бапста - Palaeocast
- Галерея граптолитов Майкла П. Климеца - Грапталиты
- Что такое ископаемые граптолиты и почему они полезны в геологии? - YouTube
- Написание на скалах - Геологический музей Стивена Хуи
| Базы данных управления авторитетом : национальный |
|---|