Нотч-белки
| Notch (LNR) домен | |||
|---|---|---|---|
 | |||
| Идентификаторы | |||
| Символ | Нотч | ||
| Пфам | PF00066 | ||
| ИнтерПро | ИПР000800 | ||
| УМНЫЙ | SM00004 | ||
| PROSITE | ПС50258 | ||
| Суперсемейство OPM | 462 | ||
| белок OPM | 5кзо | ||
| Мембраном | 19 | ||
| |||
Белки Notch представляют собой трансмембранных семейство белков типа 1 , которые образуют основной компонент сигнального пути Notch , который высоко консервативен у животных . Notch Внеклеточный домен опосредует взаимодействия с лигандами семейства DSL , позволяя ему участвовать в юкстакринной передаче сигналов . Внутриклеточный домен Notch действует как активатор транскрипции в комплексе с транскрипционными факторами семейства CSL . Члены этого семейства трансмембранных белков типа 1 имеют общие основные структуры, включая внеклеточный домен, состоящий из множества повторов, подобных эпидермальному фактору роста (EGF), и внутриклеточный домен активации транскрипции (TAD). Члены семейства Notch действуют во множестве различных тканей и играют роль в различных процессах развития, контролируя решения о судьбе клеток . Большая часть того, что известно о функции Notch, получено из исследований, проведенных на Caenorhabditis elegans ( C.elegans ) и Drosophila melanogaster . Человеческие гомологи также были идентифицированы, но детали функции Notch и взаимодействий с его лигандами в этом контексте недостаточно известны.
Открытие
[ редактировать ]Нотч был обнаружен у мутанта -дрозофилы в марте 1913 года в лаборатории Томаса Ханта Моргана . [2] Этот мутант появился после нескольких поколений вычеркивания и обратного скрещивания бисерокрылых мух с мухами дикого типа и впервые был охарактеризован Джоном С. Декстером. [3] Наиболее часто наблюдаемый фенотип у мух-мутантов Notch - это появление вогнутой зазубрины на самом дистальном конце крыльев, в честь которого назван ген , сопровождающееся отсутствием краевых щетинок. [4] [5] Было обнаружено, что этот мутант является сцепленной с полом доминантой в Х-хромосоме , которую можно наблюдать только у гетерозиготных женщин, поскольку он был смертельным для мужчин и гомозиготных женщин. [2] Первый аллель Notch был установлен в 1917 году CW Metz и CB Bridges. [6] В конце 1930-х годов исследования эмбриогенеза мух , проведенные Дональдом Ф. Поулсоном, дали первые указания на роль Нотча в развитии. [7] У самцов-мутантов Notch-8 отсутствовали внутренние зародышевые листки , энтодерма и мезодерма , что приводило к неспособности подвергнуться более позднему морфогенезу и эмбриональной летальности. Более поздние исследования раннего дрозофил нейрогенеза предоставили некоторые из первых указаний на роль Notch в передаче сигналов между клетками, поскольку нервная система у мутантов Notch развивалась путем принесения в жертву гиподермальных клеток. [8]
Начиная с 1980-х годов исследователи начали получать дальнейшее представление о функции Notch посредством генетических и молекулярных экспериментов. Генетический скрининг, проведенный на дрозофиле, привел к идентификации нескольких белков, которые играют центральную роль в передаче сигналов Notch, включая Enhancer of Split, [8] Главный разум, Дельта, [9] Подавитель бесшерстности (CSL), [10] и Серрейт. [11] В то же время ген Notch был успешно секвенирован. [12] [13] и клонирован, [14] [15] предоставив представление о молекулярной архитектуре белков Notch и приведя к идентификации гомологов Notch у Caenorhabditis elegans ( C. elegans ). [16] [17] [18] и, в конечном итоге, у млекопитающих .
В начале 1990-х годов Notch все чаще считался рецептором ранее неизвестного межклеточного сигнального пути. [19] [20] при котором межклеточный домен Notch (NICD) транспортируется в ядро , где он действует как фактор транскрипции, напрямую регулируя гены-мишени. [21] [22] [23] Было обнаружено, что высвобождение NICD происходит в результате протеолитического расщепления трансмембранного белка под действием комплекса γ-секретазы каталитической субъединицы пресенилина . Это было значительное взаимодействие, поскольку пресенилин участвует в развитии болезни Альцгеймера. [24] Это и дальнейшие исследования механизма передачи сигналов Notch привели к исследованиям, которые в дальнейшем свяжут Notch с широким спектром заболеваний человека.
Структура
[ редактировать ]Дрозофила содержит единственный белок Notch C. , elegans содержат два резервных паралога выреза, Lin-12. [25] и ГПП-1, [18] [26] и у людей есть четыре варианта Notch: Notch 1-4. Хотя между гомологами существуют вариации, существует набор высококонсервативных структур, обнаруженных во всех белках семейства Notch. Белок можно в общих чертах разделить на внеклеточный домен Notch (NECD) и внутриклеточный домен Notch (NICD), соединенные вместе однопроходным трансмембранным доменом (TM).
NECD содержит 36 повторов EGF у дрозофилы . [13] 28–36 у людей и 13 и 10 у C. elegans Lin-12 и GLP-1 соответственно. [27] Эти повторы сильно модифицируются посредством O-гликозлиирования. [28] и было показано, что добавление специфических О-связанных гликанов необходимо для правильного функционирования. За повторами EGF следуют три богатых цистеином повторов Lin-12/Notch (LNR) и домен гетеродимеризации (HD). Вместе LNR и HD составляют негативную регуляторную область, прилегающую к клеточной мембране , и помогают предотвратить передачу сигналов в отсутствие связывания лиганда.
NICD действует как фактор транскрипции, который высвобождается после того, как связывание лиганда запускает его расщепление. Он содержит последовательность ядерной локализации (NLS), которая обеспечивает его транслокацию в ядро , где он образует транскрипционный комплекс вместе с несколькими другими факторами транскрипции. Попав в ядро, несколько анкириновых повторов и взаимодействие домена RAM между белками NICD и CSL образуют комплекс активации транскрипции. [29] У людей дополнительный домен PEST играет роль в деградации NICD. [30]
Функция
[ редактировать ]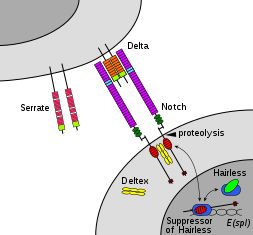
Члены семейства Notch играют роль во множестве процессов развития, контролируя решения о судьбе клеток. Сигнальная сеть Notch представляет собой эволюционно консервативный межклеточный сигнальный путь , который регулирует взаимодействия между физически соседними клетками. У дрозофилы взаимодействие notch с его связанными с клеткой лигандами (дельта, зубчатым) устанавливает межклеточный сигнальный путь, который играет ключевую роль в развитии. Этот белок действует как рецептор для мембраносвязанных лигандов и может играть множество ролей во время развития. [31] Дефицит может быть связан с двустворчатым аортальным клапаном . [32]
Есть доказательства, что активированные Notch 1 и Notch 3 способствуют дифференцировке клеток-предшественников в астроглию . [33] Notch 1, активируемый еще до рождения, индуцирует радиальной глии . дифференцировку [34] но постнатально индуцирует дифференцировку в астроциты . [35] Одно исследование показывает, что каскад Notch-1 активируется Рилином неустановленным способом. [36] Рилин и Notch1 сотрудничают в развитии зубчатой извилины . Согласно другому мнению, [37]
Лигандные взаимодействия
[ редактировать ]| Зубчатый/зубчатый белок | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | DSL | ||
| Пфам | PF01414 | ||
| ИнтерПро | ИПР026219 | ||
| Мембраном | 76 | ||
| |||
Передача сигналов Notch запускается посредством прямого контакта между клетками, опосредованного взаимодействиями между белком рецептора Notch в клетке, принимающей сигнал, и лигандом в соседней клетке, передающей сигнал. Эти однопроходные трансмембранные белки типа 1 относятся к семейству белков Delta/Serrate/Lag-2 (DSL), названному в честь трех канонических лигандов Notch. [19] Дельта и зубчатая встречаются у Drosophila, а Lag-2 — у C. elegans . Люди содержат 3 гомолога Delta, Delta-like 1 , 3 и 4 , а также два зубчатых гомолога, Jagged 1 и 2 . Белки Notch состоят из относительно короткого внутриклеточного домена и большого внеклеточного домена с одним или несколькими мотивами EGF и N-концевым мотивом связывания DSL. Было показано, что повторы 11-12 EGF во внеклеточном домене Notch необходимы и достаточны для транс -сигнальных взаимодействий между Notch и его лигандами. [38] Кроме того, повторы 24-29 EGF участвуют в ингибировании цис- взаимодействий между Notch и лигандами, совместно экспрессируемыми в одной и той же клетке. [39]
Протеолиз
[ редактировать ]Для того чтобы произошло сигнальное событие, белок Notch должен быть расщеплен в нескольких сайтах. У людей Notch сначала расщепляется в домене NRR фурином во время процессинга в транс-сети Гольджи, а затем презентируется на поверхности клетки в виде гетеродимера. [40] [41] Drosophila Notch не требует такого расщепления для передачи сигналов. [42] и есть некоторые данные, позволяющие предположить, что LIN-12 и GLP-1 расщепляются в этом сайте у C. elegans .
Выпуск NICD достигается после двух дополнительных событий расщепления Notch. Связывание Notch с лигандом DSL приводит к конформационным изменениям, которые обнажают сайт расщепления в NECD. Ферментативный протеолиз в этом сайте осуществляется протеазой семейства доменов дизинтегрина и металлопротеазы (ADAM). называется кузбанским Этот белок у дрозофилы . [43] [44] суп-17 у C. elegans , [45] и ADAM10 у людей. [46] [47] После протеолитического расщепления высвободившийся NECD эндоцитозируется в клетку, передающую сигнал, оставляя после себя лишь небольшую внеклеточную часть Notch. Этот усеченный белок Notch затем может быть распознан γ-секретазой, которая расщепляет третий сайт, обнаруженный в домене TM. [48]
Человеческие гомологи
[ редактировать ]Нотч-1
[ редактировать ]Нотч-2
[ редактировать ]Notch-2 ( белок-гомолог нейрогенного локуса notch 2 ) — это белок , который у человека кодируется NOTCH2 геном . [49]
NOTCH2 связан с синдромом Алажиля [50] и синдром Хайду-Чейни . [51]
Нотч-3
[ редактировать ]Нотч-4
[ редактировать ]См. также
[ редактировать ]Примечания
[ редактировать ]- ^ Вардар Д., Норт КЛ, Санчес-Иризарри К., Астер Дж.К., Блэклоу СК (июнь 2003 г.). «Структура ядерного магнитного резонанса прототипа повторяющегося модуля Lin12-Notch человеческого Notch1». Биохимия . 42 (23): 7061–7. дои : 10.1021/bi034156y . ПМИД 12795601 .
- ^ Jump up to: а б Морган TH, Бриджес CB (1916). Наследование, сцепленное с полом, у дрозофилы . Библиотеки НГУ. Вашингтон, Институт Карнеги в Вашингтоне.
- ^ Декстер Дж. С. (декабрь 1914 г.). «Анализ случая непрерывной изменчивости дрозофилы путем изучения ее связей» . Американский натуралист . 48 (576): 712–758. дои : 10.1086/279446 . hdl : 2027/nnc1.cu56096100 .
- ^ Мор О.Л. (май 1919 г.). «Изменения характера, вызванные мутацией целого участка хромосомы у дрозофилы» . Генетика . 4 (3): 275–82. дои : 10.1093/генетика/4.3.275 . ПМК 1200460 . ПМИД 17245926 .
- ^ Линдсли Д.Л., Зимм Г.Г. (2 декабря 2012 г.). Геном дрозофилы Melanogaster . Академическая пресса. ISBN 9780323139847 .
- ^ Мец CW, Бриджес CB (декабрь 1917 г.). «Несовместимость мутантных рас у дрозофилы» . Труды Национальной академии наук Соединенных Штатов Америки . 3 (12): 673–8. Бибкод : 1917PNAS....3..673M . дои : 10.1073/pnas.3.12.673 . ПМЦ 1091355 . ПМИД 16586764 .
- ^ Поулсон Д.Ф. (март 1937 г.). «Хромосомные дефекты и эмбриональное развитие дрозофилы Melanogaster» . Труды Национальной академии наук Соединенных Штатов Америки . 23 (3): 133–7. Бибкод : 1937ПНАС...23..133П . дои : 10.1073/pnas.23.3.133 . ПМЦ 1076884 . ПМИД 16588136 .
- ^ Jump up to: а б Леманн Р., Хименес Ф., Дитрих У., Кампос-Ортега Х.А. (март 1983 г.). «О фенотипе и развитии мутантов раннего нейрогенеза у Drosophila melanogaster». Архив биологии развития Вильгельма Ру . 192 (2): 62–74. дои : 10.1007/BF00848482 . ПМИД 28305500 . S2CID 25602190 .
- ^ Леманн Р., Дитрих У., Хименес Ф., Кампос-Ортега Х.А. (июль 1981 г.). «Мутации раннего нейрогенеза у дрозофилы». Архив биологии развития Вильгельма Ру . 190 (4): 226–229. дои : 10.1007/BF00848307 . ПМИД 28305572 . S2CID 21814447 .
- ^ Фортини М.Э., Артаванис-Цаконас С. (октябрь 1994 г.). «Супрессор белка безволосости участвует в передаче сигналов рецептора Notch». Клетка . 79 (2): 273–82. дои : 10.1016/0092-8674(94)90196-1 . ПМИД 7954795 . S2CID 40771329 .
- ^ Флеминг Р.Дж., Скоттгейл Т.Н., Дидерих Р.Дж., Артаванис-Цаконас С. (декабрь 1990 г.). «Ген Serrate кодирует предполагаемый EGF-подобный трансмембранный белок, необходимый для правильного эктодермального развития у Drosophila melanogaster» . Гены и развитие . 4 (12А): 2188–201. дои : 10.1101/gad.4.12a.2188 . ПМИД 2125287 .
- ^ Кидд С., Келли М.Р., Янг М.В. (сентябрь 1986 г.). «Последовательность локуса notch Drosophila melanogaster: связь кодируемого белка с факторами свертывания крови и роста млекопитающих» . Молекулярная и клеточная биология . 6 (9): 3094–108. дои : 10.1128/mcb.6.9.3094 . ПМК 367044 . ПМИД 3097517 .
- ^ Jump up to: а б Уортон К.А., Йохансен К.М., Сюй Т., Артаванис-Цаконас С. (декабрь 1985 г.). «Нуклеотидная последовательность нейрогенного локуса notch подразумевает генный продукт, который имеет гомологию с белками, содержащими EGF-подобные повторы» . Клетка . 43 (3, часть 2): 567–81. дои : 10.1016/0092-8674(85)90229-6 . ПМИД 3935325 .
- ^ Кидд С., Локетт Т.Дж., Янг М.В. (сентябрь 1983 г.). «Локус Notch Drosophila melanogaster». Клетка . 34 (2): 421–33. дои : 10.1016/0092-8674(83)90376-8 . ПМИД 6193889 . S2CID 36425372 .
- ^ Артаванис-Цаконас С., Мускавич М.А., Едвобник Б. (апрель 1983 г.). «Молекулярное клонирование Notch, локуса, влияющего на нейрогенез у Drosophila melanogaster» . Труды Национальной академии наук Соединенных Штатов Америки . 80 (7): 1977–81. Бибкод : 1983PNAS...80.1977A . дои : 10.1073/pnas.80.7.1977 . ПМЦ 393735 . ПМИД 6403942 .
- ^ Гринвальд I (февраль 1987 г.). «Локус lin-12 Caenorhabditis elegans». Биоэссе . 6 (2): 70–3. дои : 10.1002/bies.950060207 . ПМИД 3551950 . S2CID 19253030 .
- ^ Присс-младший, Шнабель Х., Шнабель Р. (ноябрь 1987 г.). «Локус glp-1 и клеточные взаимодействия у ранних эмбрионов C. elegans». Клетка . 51 (4): 601–11. дои : 10.1016/0092-8674(87)90129-2 . ПМИД 3677169 . S2CID 6282210 .
- ^ Jump up to: а б Остин Дж., Кимбл Дж. (ноябрь 1987 г.). «glp-1 необходим в зародышевой линии для регуляции выбора между митозом и мейозом у C. elegans». Клетка . 51 (4): 589–99. дои : 10.1016/0092-8674(87)90128-0 . ПМИД 3677168 . S2CID 31484517 .
- ^ Jump up to: а б Артаванис-Цаконас С., Мацуно К., Фортини М.Э. (апрель 1995 г.). «Нетч-сигнализация». Наука . 268 (5208): 225–32. Бибкод : 1995Sci...268..225A . дои : 10.1126/science.7716513 . ПМИД 7716513 .
- ^ Гринвальд I, генеральный менеджер «Рубина» (январь 1992 г.). «Изменить ситуацию: роль межклеточных взаимодействий в установлении отдельных идентичностей эквивалентных клеток». Клетка . 68 (2): 271–81. дои : 10.1016/0092-8674(92)90470-w . ПМИД 1365402 . S2CID 11901508 .
- ^ Шретер Э.Х., Кисслингер Дж.А., Копан Р. (май 1998 г.). «Передача сигналов Notch-1 требует индуцированного лигандом протеолитического высвобождения внутриклеточного домена». Природа . 393 (6683): 382–6. Бибкод : 1998Natur.393..382S . дои : 10.1038/30756 . ПМИД 9620803 . S2CID 4431882 .
- ^ Копан Р., Най Дж.С., Вайнтрауб Х. (сентябрь 1994 г.). «Внутриклеточный домен мышиного Notch: конститутивно активированный репрессор миогенеза, направленный на основную область спираль-петля-спираль MyoD». Разработка . 120 (9): 2385–96. дои : 10.1242/dev.120.9.2385 . ПМИД 7956819 .
- ^ Струл Г., Фицджеральд К., Гринвальд I (июль 1993 г.). «Внутренняя активность внутриклеточных доменов Lin-12 и Notch in vivo». Клетка . 74 (2): 331–45. дои : 10.1016/0092-8674(93)90424-о . ПМИД 8343960 . S2CID 27966283 .
- ^ Шеррингтон Р., Рогаев Е.И., Лян Ю., Рогаева Е.А., Левеск Г., Икеда М. и др. (июнь 1995 г.). «Клонирование гена, несущего миссенс-мутации при ранней семейной болезни Альцгеймера». Природа . 375 (6534): 754–60. Бибкод : 1995Natur.375..754S . дои : 10.1038/375754a0 . ПМИД 7596406 . S2CID 4308372 .
- ^ Гринвальд И.С., Штернберг П.В., Хорвиц Х.Р. (сентябрь 1983 г.). «Локус lin-12 определяет судьбу клеток Caenorhabditis elegans». Клетка . 34 (2): 435–44. дои : 10.1016/0092-8674(83)90377-x . ПМИД 6616618 . S2CID 40668388 .
- ^ Остин Дж., Кимбл Дж. (август 1989 г.). «Анализ транскриптов glp-1 и lin-12, гомологичных генов, необходимых для клеточных взаимодействий во время развития C. elegans». Клетка . 58 (3): 565–71. дои : 10.1016/0092-8674(89)90437-6 . ПМИД 2758467 . S2CID 8514017 .
- ^ Гринвальд I (декабрь 1985 г.). «lin-12, гомеотический ген нематод, гомологичен набору белков млекопитающих, который включает эпидермальный фактор роста» . Клетка . 43 (3, часть 2): 583–90. дои : 10.1016/0092-8674(85)90230-2 . ПМИД 3000611 .
- ^ Шао Л., Луо Ю., Молони DJ, Халтивангер Р. (ноябрь 2002 г.). «О-гликозилирование повторов EGF: идентификация и первоначальная характеристика УДФ-глюкозы: протеин-О-глюкозилтрансфераза» . Гликобиология . 12 (11): 763–70. дои : 10.1093/гликоб/cwf085 . ПМИД 12460944 .
- ^ Тамура К., Танигучи И., Миногучи С., Сакаи Т., Тун Т., Фурукава Т., Хондзё Т. (декабрь 1995 г.). «Физическое взаимодействие между новым доменом рецептора Notch и фактором транскрипции RBP-J каппа/Su(H)». Современная биология . 5 (12): 1416–23. дои : 10.1016/S0960-9822(95)00279-X . hdl : 2433/202204 . ПМИД 8749394 . S2CID 18442572 .
- ^ Венг А.П., Феррандо А.А., Ли В., Моррис Дж.П., Сильверман Л.Б., Санчес-Иризарри С. и др. (октябрь 2004 г.). «Активирующие мутации NOTCH1 при остром Т-клеточном лимфобластном лейкозе человека». Наука . 306 (5694): 269–71. Бибкод : 2004Sci...306..269W . дои : 10.1126/science.1102160 . ПМИД 15472075 . S2CID 24049536 .
- ^ «Ген Энтрез: гомолог 1 NOTCH1 Notch, связанный с транслокацией (дрозофила)» .
- ^ МакКеллар С.Х., Тестер DJ, Ягубян М., Маджумдар Р., Акерман М.Дж., Сундт Т.М. (август 2007 г.). «Новые мутации NOTCH1 у пациентов с заболеванием двустворчатого аортального клапана и аневризмами грудной аорты» . Журнал торакальной и сердечно-сосудистой хирургии . 134 (2): 290–6. дои : 10.1016/j.jtcvs.2007.02.041 . ПМИД 17662764 .
- ^ Танигаки К., Ногаки Ф., Такахаси Дж., Таширо К., Курока Х., Хондзё Т. (январь 2001 г.). «Notch1 и Notch3 инструктивно ограничивают мультипотентные нейрональные клетки-предшественники, реагирующие на bFGF, астроглиальной судьбой». Нейрон . 29 (1): 45–55. дои : 10.1016/S0896-6273(01)00179-9 . hdl : 2433/150564 . ПМИД 11182080 . S2CID 17047028 .
- ^ Гаяно Н., Най Дж. С., Фишелл Дж. (май 2000 г.). «Идентификация радиальной глии обеспечивается передачей сигналов Notch1 в переднем мозге мышей» . Нейрон . 26 (2): 395–404. дои : 10.1016/S0896-6273(00)81172-1 . ПМИД 10839358 . S2CID 15861936 .
- ^ Чемберс CB, Пэн Й., Нгуен Х., Гайано Н., Фишелл Г., Най Дж.С. (март 2001 г.). «Пространственно-временная избирательность ответа на сигналы Notch1 у предшественников переднего мозга млекопитающих» . Разработка . 128 (5): 689–702. дои : 10.1242/dev.128.5.689 . ПМИД 11171394 .
- ^ Кейлани С., Сугая К. (июль 2008 г.). «Рилин индуцирует фенотип радиальной глии в нервных клетках-предшественниках человека путем активации Notch-1» . Биология развития BMC . 8 (1): 69. дои : 10.1186/1471-213X-8-69 . ПМЦ 2447831 . ПМИД 18593473 .
- ^ Сиббе М., Фёрстер Э., Басак О., Тейлор В., Фротшер М. (июль 2009 г.). «Рилин и Notch1 сотрудничают в развитии зубчатой извилины» . Журнал неврологии . 29 (26): 8578–85. doi : 10.1523/JNEUROSCI.0958-09.2009 . ПМК 6665659 . ПМИД 19571148 .
- ^ Ребай И., Флеминг Р.Дж., Фехон Р.Г., Чербас Л., Чербас П., Артаванис-Цаконас С. (ноябрь 1991 г.). «Специфические повторы EGF Notch опосредуют взаимодействие с Delta и Serrate: значение Notch как многофункционального рецептора». Клетка . 67 (4): 687–99. дои : 10.1016/0092-8674(91)90064-6 . ПМИД 1657403 . S2CID 12643727 .
- ^ де Селис Дж. Ф., Брэй С. Дж. (март 2000 г.). «Домен Abruptex Notch регулирует негативные взаимодействия между Notch, его лигандами и Fringe» . Разработка . 127 (6): 1291–302. дои : 10.1242/dev.127.6.1291 . ПМИД 10683181 .
- ^ Блаумюллер СМ, Ци Х, Загурас П, Артаванис-Цаконас С (июль 1997 г.). «Внутриклеточное расщепление Notch приводит к образованию гетеродимерного рецептора на плазматической мембране» . Клетка . 90 (2): 281–91. дои : 10.1016/s0092-8674(00)80336-0 . ПМИД 9244302 . S2CID 16544864 .
- ^ Логит Ф, Бессия С, Бру С, ЛеБей О, Жаррио С, Сейда Н.Г., Исраэль А (июль 1998 г.). «Рецептор Notch1 конститутивно расщепляется фуриноподобной конвертазой» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (14): 8108–12. Бибкод : 1998PNAS...95.8108L . дои : 10.1073/pnas.95.14.8108 . ЧВК 20937 . ПМИД 9653148 .
- ^ Кидд С., Либер Т. (июль 2002 г.). «Расщепление фурином не является обязательным условием для функции Notch дрозофилы» . Механизмы развития . 115 (1–2): 41–51. дои : 10.1016/s0925-4773(02)00120-x . ПМИД 12049766 . S2CID 10249987 .
- ^ Рук Дж., Пан Д., Сюй Т., Рубин Г.М. (август 1996 г.). «KUZ, консервативный белок металлопротеаза-дезинтегрин, играющий две роли в нейрогенезе дрозофилы». Наука . 273 (5279): 1227–31. Бибкод : 1996Sci...273.1227R . дои : 10.1126/science.273.5279.1227 . ПМИД 8703057 . S2CID 10984937 .
- ^ Пан Д, генеральный менеджер «Рубина» (июль 1997 г.). «Кузбаниан контролирует протеолитическую обработку Notch и опосредует латеральное торможение во время нейрогенеза дрозофилы и позвоночных» . Клетка . 90 (2): 271–80. дои : 10.1016/s0092-8674(00)80335-9 . ПМИД 9244301 . S2CID 18819481 .
- ^ Вэнь С., Мецштайн М.М., Гринвальд I (декабрь 1997 г.). «SUP-17, белок ADAM Caenorhabditis elegans, родственный Drosophila KUZBANIAN, и его роль в передаче сигналов LIN-12/NOTCH». Разработка . 124 (23): 4759–67. дои : 10.1242/dev.124.23.4759 . ПМИД 9428412 .
- ^ Ховард Л., Глинн П. (1995). «Мембранассоциированная металлопротеиназа, распознаваемая путем характерного расщепления основного белка миелина: анализ и выделение». Протеолитические ферменты: аспарагиновая и металлопептидазы . Методы энзимологии. Том. 248. стр. 388–95. дои : 10.1016/0076-6879(95)48025-0 . ISBN 9780121821494 . ПМИД 7545777 .
- ^ Ланн К.А., Фан Икс, Дейли Б., Миллер К., Заводный П.Дж., Нарула С.К., Ланделл Д. (январь 1997 г.). «Очистка ADAM 10 из бычьей селезенки в виде конвертазы TNF-альфа» . Письма ФЭБС . 400 (3): 333–5. дои : 10.1016/s0014-5793(96)01410-x . ПМИД 9009225 . S2CID 83810622 .
- ^ Струл Г., Адачи А. (сентябрь 2000 г.). «Требования к пресенилин-зависимому расщеплению notch и других трансмембранных белков» . Молекулярная клетка . 6 (3): 625–36. дои : 10.1016/s1097-2765(00)00061-7 . ПМИД 11030342 .
- ^ Ларссон С., Ларделли М., Уайт И., Лендал У. (ноябрь 1994 г.). «Гены NOTCH1, 2 и 3 человека расположены в положениях хромосом 9q34, 1p13-p11 и 19p13.2-p13.1 в областях транслокации, связанной с неоплазией». Геномика . 24 (2): 253–8. дои : 10.1006/geno.1994.1613 . ПМИД 7698746 .
- ^ Самеджима Х, Тории С, Косаки Р, Куросава К, Ёшихаси Х, Муроя К, Окамото Н, Ватанабе Ю, Кошо Т, Кубота М, Мацуда О, Гото М, Изуми К, Такахаси Т, Косаки К (2007). «Скрининг мутаций синдрома Алажилля в генах JAG1 и NOTCH2 с использованием денатурирующей высокоэффективной жидкостной хроматографии». Генетическое тестирование . 11 (3): 216–27. дои : 10.1089/gte.2006.0519 . ПМИД 17949281 .
- ^ Симпсон М.А., Ирвинг М.Д., Асилмаз Э., Грей М.Дж., Дафу Д., Элмсли Ф.В., Мансур С., Холдер С.Е., Брэйн К.Э., Бертон Б.К., Ким К.Х., Паули Р.М., Афтимос С., Стюарт Х., Ким К.А., Холдер-Эспинасс М., Робертсон С.П., Дрейк В.М., Трембат RC (март 2011 г.). «Мутации в NOTCH2 вызывают синдром Хайду-Чейни, заболевание, характеризующееся тяжелой и прогрессирующей потерей костной массы». Природная генетика . 43 (4): 303–5. дои : 10.1038/ng.779 . ПМИД 21378985 . S2CID 205357391 .
Ссылки
[ редактировать ]- Tax FE, Yeargers JJ, Thomas JH (март 1994 г.). «Последовательность C. elegans lag-2 обнаруживает домен клеточной сигнализации, общий с Delta и Serrate of Drosophila». Природа . 368 (6467): 150–4. Бибкод : 1994Natur.368..150T . дои : 10.1038/368150a0 . ПМИД 8139658 . S2CID 4345508 .
- Линдселл С.Э., Шаубер С.Дж., Боултер Дж., Вайнмастер Дж. (март 1995 г.). «Jagged: лиганд млекопитающих, который активирует Notch1» . Клетка . 80 (6): 909–17. дои : 10.1016/0092-8674(95)90294-5 . ПМИД 7697721 . S2CID 11720367 .
- Келли М.Р., Кидд С., Дойч В.А., Янг М.В. (ноябрь 1987 г.). «Мутации, изменяющие структуру кодирующих последовательностей, подобных эпидермальному фактору роста, в локусе Notch дрозофилы». Клетка . 51 (4): 539–48. дои : 10.1016/0092-8674(87)90123-1 . ПМИД 3119223 . S2CID 34338340 .