Гетерозис
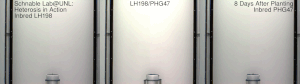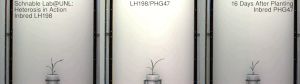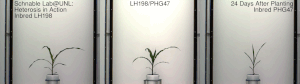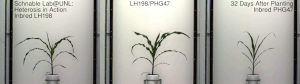
Гетерозис , гибридная сила или аутбридинг – это улучшение или усиление функции любого биологического качества у гибридного потомства. Потомство является гетеротичным, если его признаки усиливаются в результате смешения генетических вкладов его родителей. Гетеротическое потомство часто имеет черты, которые представляют собой нечто большее, чем простое сложение признаков родителей, и может быть объяснено менделевским или неменделевским наследованием . [ 1 ] Типичными гетерозисными/гибридными характеристиками, представляющими интерес для сельского хозяйства, являются более высокая урожайность, более быстрое созревание, стабильность, засухоустойчивость. [ 2 ] и т. д.
Определения
[ редактировать ]Предлагая термин «гетерозис» вместо старого термина «гетерозиготность» , Г.Х. Шулл стремился избежать ограничения этого термина эффектами, которые можно объяснить гетерозиготностью при менделевском наследовании. [ 3 ]
Физиологическая сила организма, проявляющаяся в быстроте его роста, росте и общей крепости, находится в положительной зависимости от степени несходства гамет, в результате слияния которых образовался организм... Чем многочисленнее различия между объединяющимися гаметами — у хотя бы в определенных пределах — тем больше в целом объем стимуляции… Эти различия не обязательно должны быть менделевскими по своему наследованию… Чтобы избежать предположения, что все генотипические различия, которые стимулируют деление клеток, рост и другие физиологические активности организма, являются Менделя в их наследии, а также для краткости выражения я предлагаю… принять слово «гетерозис».
Гетерозис часто обсуждается как противоположность инбредной депрессии , хотя различия в этих двух концепциях можно увидеть в эволюционных соображениях, таких как роль генетической изменчивости или влияние генетического дрейфа в небольших популяциях на эти концепции. Инбредная депрессия возникает, когда у родственных родителей рождаются дети с чертами , которые отрицательно влияют на их приспособленность, во многом из-за гомозиготности . В таких случаях ауткроссинг должен привести к гетерозису.
Не все ауткроссы приводят к гетерозису. Например, когда гибрид наследует от своих родителей не полностью совместимые черты, приспособленность может снизиться. Это форма аутбредной депрессии , последствия которой аналогичны инбредной депрессии. [ 4 ]
Генетические и эпигенетические основы
[ редактировать ]С начала 1900-х годов для объяснения силы гибридов были разработаны две конкурирующие генетические гипотезы, не обязательно взаимоисключающие. Совсем недавно эпигенетический компонент гибридной силы. также был установлен [ 5 ] [ 6 ]
Доминирование и сверхдоминирование
[ редактировать ]Когда популяция небольшая или инбредная, она имеет тенденцию терять генетическое разнообразие. Инбредная депрессия — это потеря приспособленности из-за потери генетического разнообразия. Инбредные линии, как правило, гомозиготны по рецессивным аллелям , которые являются умеренно вредными (или производят признак, нежелательный с точки зрения селекционера). С другой стороны, гетерозис или гибридная сила — это тенденция аутбредных линий превосходить обоих инбредных родителей по приспособленности.
Селекция растений и животных, включая гибридизацию, началась задолго до того, как появилось понимание основополагающих научных принципов. В начале 20 века, после того как законы Менделя стали понятны и приняты, генетики взялись объяснить превосходную силу роста многих гибридов растений. Были разработаны две конкурирующие гипотезы, не исключающие друг друга: [ 7 ]
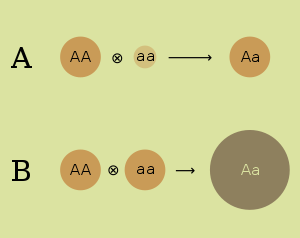
- Гипотеза доминирования . Гипотеза доминирования объясняет превосходство гибридов подавлением нежелательных рецессивных аллелей одного родителя доминантными аллелями другого. Он объясняет плохую продуктивность инбредных штаммов потерей генетического разнообразия, при этом штаммы становятся чисто гомозиготными по многим локусам. Гипотеза доминирования была впервые высказана в 1908 году генетиком Чарльзом Дэвенпортом . [ 8 ] Согласно гипотезе доминирования, ожидается, что вредные аллели будут поддерживаться в популяции случайных спариваний при балансе отбора и мутации, который будет зависеть от скорости мутаций, эффекта аллелей и степени, в которой аллели экспрессируются в гетерозиготах. [ 9 ]
- Гипотеза сверхдоминирования . Определенные комбинации аллелей, которые можно получить путем скрещивания двух инбредных линий, являются выгодными в гетерозиготе . Гипотеза сверхдоминирования объясняет преимущество гетерозигот выживанием многих аллелей, которые являются рецессивными и вредными у гомозигот. Он объясняет плохую производительность инбредных линий высоким процентом этих вредных рецессивов. Гипотеза сверхдоминирования была независимо разработана Эдвардом М. Истом (1908). [ 10 ] и Джордж Шулл (1908). [ 11 ] Ожидается, что генетическая изменчивость в сверхдоминантном локусе будет поддерживаться за счет балансирующего отбора. Высокая приспособленность гетерозиготных генотипов способствует сохранению аллельного полиморфизма в популяции. [ 9 ] Эту гипотезу обычно используют для объяснения устойчивости некоторых аллелей (наиболее известный аллель серповидноклеточного признака ), которые вредны для гомозигот. В обычных обстоятельствах такие вредные аллели были бы удалены из популяции в процессе естественного отбора. Как и гипотеза доминирования, она объясняет плохую продуктивность инбредных линий экспрессией таких вредных рецессивных аллелей.
Доминирование и сверхдоминирование имеют разные последствия для профиля экспрессии генов у людей. Если сверхдоминирование является основной причиной преимуществ гетерозиса в приспособленности, то у гетерозиготного потомства должна быть сверхэкспрессия определенных генов по сравнению с гомозиготными родителями. С другой стороны, если причиной является доминирование, у гетерозиготного потомства должно быть недостаточно экспрессировано меньше генов по сравнению с родителями. Более того, экспрессия любого данного гена должна быть сопоставима с той, которая наблюдается у наиболее приспособленных двух родителей. В любом случае ауткроссное скрещивание дает преимущество, заключающееся в маскировке вредных рецессивных аллелей в потомстве. Было предложено считать это преимущество основным фактором поддержания полового размножения среди эукариот, как это резюмировано в статье « Эволюция полового размножения» .
Историческая ретроспектива
[ редактировать ]Какой из двух механизмов является «главной» причиной гетерозиса, вызывает научные споры в области генетики . [ 12 ] Популяционный генетик Джеймс Кроу (1916–2012) в молодости считал, что чрезмерное доминирование является основным фактором, способствующим жизнеспособности гибридов. В 1998 году он опубликовал ретроспективный обзор развивающейся науки. [ 13 ] По словам Кроу, демонстрация нескольких случаев преимущества гетерозигот у дрозофилы и других организмов впервые вызвала большой энтузиазм по поводу теории сверхдоминирования среди ученых, изучающих гибридизацию растений. Но сверхдоминирование подразумевает, что урожайность инбредных линий должна снижаться по мере того, как инбредные линии отбираются для получения результатов гибридных скрещиваний, поскольку доля вредных рецессивов в инбредной популяции возрастает. За прошедшие годы эксперименты в области генетики растений доказали, что происходит обратное: урожайность увеличивается как у инбредных штаммов, так и у гибридов, что позволяет предположить, что одного доминирования может быть достаточно, чтобы объяснить превосходную урожайность гибридов. Во всей генетике зарегистрировано лишь несколько убедительных случаев сверхдоминирования. С 1980-х годов, по мере накопления экспериментальных данных, теория доминирования вернулась.
Ворона написал:
Современная точка зрения... заключается в том, что гипотеза доминирования является основным объяснением упадка инбридинга и высокой урожайности гибридов. мало Статистических данных о вкладе сверхдоминирования и эпистаза . Но получают ли лучшие гибриды дополнительный импульс от сверхдоминирования или благоприятного эпистатического вклада, остается открытым вопросом. [ 13 ]
Эпигенетика
[ редактировать ]гетерозис . У растений установлен эпигенетический вклад в [ 6 ] и это также было зарегистрировано у животных. [ 14 ] МикроРНК (миРНК), открытые в 1993 году, представляют собой класс некодирующих малых РНК, которые подавляют трансляцию информационных РНК (мРНК) или вызывают деградацию мРНК. [ 15 ] У гибридных растений большинство микроРНК имеют неаддитивную экспрессию (она может быть выше или ниже, чем у родителей). [ 6 ] Это говорит о том, что малые РНК участвуют в росте, жизнеспособности и адаптации гибридов. [ 6 ]
Влияние «гетерозиса без гибридности» на размер растения было продемонстрировано на генетически изогенных триплоидных (автополиплоидных) растениях F1, где избыток отцовского генома триплоиды F1 демонстрируют положительный гетерозис, тогда как избыток F1 в материнском геноме демонстрирует отрицательные эффекты гетерозиса. [ 16 ] Такие результаты показывают, что эффекты гетерозиса с эпигенетической основой, зависящей от дозы генома, могут возникать у потомков F1, которые являются генетически изогенными (т.е. не имеют гетерозиготности). [ 16 ] [ 17 ] Было показано [ 5 ] Эта гибридная сила аллополиплоидного гибрида двух видов Arabidopsis была обусловлена эпигенетическим контролем в верхних областях двух генов, что вызвало серьезные последующие изменения в накоплении хлорофилла и крахмала. Механизм включает ацетилирование или метилирование определенных аминокислот в гистоне H3 , белке, тесно связанном с ДНК, который может либо активировать, либо подавлять ассоциированные гены.
Конкретные механизмы
[ редактировать ]Главный комплекс гистосовместимости у животных
[ редактировать ]Одним из примеров того, где определенные гены могут быть важны у позвоночных животных для гетерозиса, является главный комплекс гистосовместимости (MHC). Позвоночные животные наследуют несколько копий MHC класса I и MHC класса II от каждого родителя, которые используются в презентации антигена как часть адаптивной иммунной системы. Каждая копия генов способна связывать и представлять различный набор потенциальных пептидов Т-лимфоцитам . Эти гены очень полиморфны во всех популяциях, но более сходны в небольших и более тесно связанных популяциях. Скрещивание между более генетически отдаленными особями снижает вероятность наследования двух одинаковых или схожих аллелей, что позволяет представить более разнообразный спектр пептидов. Таким образом, это увеличивает вероятность того, что какой-либо конкретный патоген будет распознан, и означает, что, вероятно, будет распознано больше антигенных белков любого патогена, что дает больший диапазон активации Т-клеток и, следовательно, больший ответ. Это также означает, что иммунитет, приобретенный к возбудителю, направлен против более широкого спектра антигенов, а это означает, что возбудитель должен мутировать больше, прежде чем иммунитет будет потерян. Таким образом, гибриды менее подвержены патогенным заболеваниям и более способны бороться с инфекцией. Однако это может быть причиной аутоиммунных заболеваний. [ нужна ссылка ]
Растения
[ редактировать ]Скрещивание инбредных растений из разных гетеротических групп приводит к образованию энергичных гибридов F1 со значительно большим гетерозисом, чем у гибридов F1 от инбредных растений одной и той же гетеротической группы или образца. Гетеротические группы создаются селекционерами для классификации инбредных линий и могут постепенно улучшаться путем взаимного рекуррентного отбора.
Гетерозис используется для повышения урожайности, однородности и силы роста. Методы гибридной селекции используются при выращивании кукурузы , сорго , риса , сахарной свеклы , лука , шпината , подсолнечника , брокколи и для создания более психоактивного каннабиса .
Кукуруза (маис)
[ редактировать ]Почти вся полевая кукуруза ( кукуруза ), выращиваемая в большинстве развитых стран, демонстрирует гетерозис. Современные гибриды кукурузы значительно превосходят традиционные сорта и лучше реагируют на удобрения .
Гетероз кукурузы был широко продемонстрирован в начале 20-го века Джорджем Х. Шулом и Эдвардом М. Истом изобрел гибридную кукурузу после того, как доктор Уильям Джеймс Бил из Мичиганского государственного университета на основе работы, начатой в 1879 году по настоянию Чарльза Дарвина . Работа доктора Била привела к первому опубликованному в 1881 году отчету о полевом эксперименте , демонстрирующем силу гибридов на кукурузе. Юджина Дэвенпорта и Перри Холдена Эти различные пионеры ботаники и смежных областей показали, что скрещивание инбредных линий, полученных из южной вмятины и Северный кремень, соответственно, продемонстрировал значительный гетерозис и превзошел традиционные сорта той эпохи. Однако в то время такие гибриды нельзя было экономически производить в больших масштабах для использования фермерами. Дональд Ф. Джонс на в сельскохозяйственной экспериментальной станции Коннектикута Нью-Хейвене изобрел первый практический метод производства высокоурожайной гибридной кукурузы в 1914–1917 годах. Метод Джонса позволил получить гибрид двойного скрещивания, который требует двух этапов скрещивания четырех различных исходных инбредных линий. Более поздние работы селекционеров кукурузы позволили получить инбредные линии, обладающие достаточной силой для практического производства коммерческого гибрида за один этап - гибридов простого скрещивания. Гибриды одиночного скрещивания производятся всего от двух исходных родительских инбредных растений. Они, как правило, более энергичные и более однородные, чем более ранние гибриды двойного скрещивания. Процесс создания этих гибридов часто включает в себя расчесывание .
Гибриды кукурузы умеренного климата происходят от двух основных гетерозисных групп: «Iowa Stiff Stelk Synthetic» и нежестких стеблей. [ нужна ссылка ]
Рис ( Oryza sativa )
[ редактировать ]Гибридный рис выращивается во многих странах, включая Китай, Индию, Вьетнам и Филиппины. [ 18 ] По сравнению с инбредными линиями гибриды дают примерно на 20% больший урожай и занимают 45% посевных площадей риса в Китае. [ 19 ] Производство риса в Китае значительно выросло из-за интенсивного использования гибридного риса. В Китае в результате усилий был создан супергибридный сорт риса («LYP9») с производительностью около 15 тонн с гектара. В Индии также несколько сортов показали высокую жизнеспособность, в том числе «RH-10» и «Суручи 5401». [ нужна ссылка ]
Поскольку рис является самоопыляющимся видом, для создания гибридов из отдельных линий требуется использование линий с мужской стерильностью. Самый распространенный способ добиться этого — использовать линии с генетической мужской стерильностью, поскольку ручная кастрация не оптимальна для крупномасштабной гибридизации. [ 20 ] Первое поколение гибридного риса было выведено в 1970-х годах. Он опирается на три линии: линию цитоплазматической мужской стерильности (CMS) , линию поддержания и линию восстановления. [ 19 ] Второе поколение получило широкое распространение в 1990-х годах. [ 19 ] Вместо линии CMS он использует чувствительную к окружающей среде линию генной мужской стерильности (EGMS), стерильность которой может быть изменена в зависимости от света или температуры. [ 20 ] Это устраняет необходимость в обслуживающем персонале, делая процесс гибридизации и разведения более эффективным (хотя и требующим особого ухода). Линии второго поколения показывают увеличение урожайности на 5-10% по сравнению с линиями первого поколения. [ 20 ] Третье и нынешнее поколение использует линию с ядерной мужской стерильностью (NMS). Линии третьего поколения имеют рецессивный ген стерильности, и их выращивание более щадящее по отношению к линиям-контейнерам и условиям окружающей среды. Кроме того, трансгены присутствуют только в хранителе, поэтому гибридные растения могут извлечь выгоду из гибридной силы, не требуя специального надзора. [ 19 ]
Животные
[ редактировать ]Гибридное животноводство
[ редактировать ]Понятие гетерозиса применяется также в производстве товарного скота . У крупного рогатого скота в результате скрещивания черного ангуса и герефорда получается помесь, известная как « черный лысый ». У свиней «голубые задницы» получаются в результате скрещивания Хэмпшира и Йоркшира. Другие, более экзотические гибриды (два разных вида, поэтому генетически более несходные), такие как « бифало », представляющие собой гибриды крупного рогатого скота и бизона, также используются на специализированных рынках.
Птица
[ редактировать ]У птицы домашней сцепленные с полом гены были использованы для создания гибридов, в которых самцов и самок можно сортировать по цвету в возрасте одного дня. Для этого используются специфические гены, отвечающие за полоску и рост перьев на крыльях. Кресты такого типа создают то, что продается как черные секс-линки, красные секс-линки и различные другие кресты, известные под торговыми названиями.
Коммерческие бройлеры производятся путем скрещивания различных линий Уайт Рокс и Уайт Корниш, при этом Корниш обеспечивает крупный корпус, а Рокс обеспечивает быстрый прирост. Полученная гибридная сила позволяет получать однородных птиц с товарной массой тушки в возрасте 6–9 недель.
Аналогичным образом, гибриды различных линий белого леггорна используются для производства стад-несушек, которые обеспечивают большую часть белых яиц для продажи в Соединенных Штатах.
Собаки
[ редактировать ]В 2013 году исследование показало, что смешанные породы живут в среднем на 1,2 года дольше, чем чистопородные. [ 21 ]
Джон Скотт и Джон Л. Фуллер провели детальное исследование чистокровных кокер-спаниелей, чистокровных басенджи и гибридов между ними. [ 22 ] Они обнаружили, что гибриды бегают быстрее, чем любой из родителей, возможно, из-за гетерозиса. Другие характеристики, такие как базальная частота сердечных сокращений, не выявили какого-либо гетерозиса — базальная частота сердечных сокращений собаки была близка к средней частоте сердечных сокращений ее родителей — возможно, из-за аддитивного воздействия нескольких генов. [ 23 ]
Иногда люди, работающие над программой разведения собак, не находят полезного гетерозиса. [ 24 ]
Несмотря на это, исследования не дают окончательных доказательств гибридной силы у собак. Во многом это связано с неизвестным наследием большинства используемых собак смешанных пород. Результаты сильно различаются: некоторые исследования показывают пользу, а другие обнаруживают, что собаки смешанных пород более склонны к генетическим заболеваниям. [ 25 ] [ 26 ] [ 27 ]
Птицы
[ редактировать ]В 2014 году исследование, проведенное Центром интегративной экологии Университета Дикина в Джилонге, штат Виктория, пришло к выводу, что внутривидовые гибриды между подвидами Platycercus elegans flaveolus и P. e. elegans малиновой розеллы ( P. elegans ) лучше боролись с болезнями, чем их чистые собратья. [ 28 ]
Люди
[ редактировать ]Все люди чрезвычайно генетически похожи друг на друга. [ 29 ] [ 30 ] [ 31 ] Майкл Мингрони предложил гетерозис в форме гибридной силы, связанной с историческим снижением уровня инбридинга, в качестве объяснения эффекта Флинна , устойчивого роста результатов тестов IQ во всем мире в течение 20-го века. [ нужна ссылка ] хотя обзор девяти исследований показал, что нет никаких доказательств того, что инбридинг влияет на IQ. [ 32 ]
Споры
[ редактировать ]Термин «гетерозис» часто вызывает путаницу и даже споры, особенно в селекции домашних животных , поскольку иногда (неверно) утверждается, что все гибридные растения и животные «генетически превосходят» своих родителей из-за гетерозиса. [ нужна ссылка ] . но с этим утверждением существуют две проблемы:
- Согласно статье, опубликованной в журнале Genome Biology , «генетическое превосходство» — это нечеткий термин и не общепринятая терминология в научной области генетики. [ 33 ] Соответствующий термин «приспособленность» четко определен, но его редко можно измерить напрямую. Вместо этого ученые используют объективные, измеримые величины, такие как количество семян, которые дает растение, скорость прорастания семян или процент организмов, доживающих до репродуктивного возраста. [ 34 ] С этой точки зрения гибридные растения и животные, демонстрирующие гетерозис, могут иметь «превосходные» черты, но это не обязательно означает какое-либо свидетельство явного «генетического превосходства». Использование термина «превосходство» является обычным явлением, например, в селекции сельскохозяйственных культур, где под ним понимают более урожайное и более устойчивое растение для сельского хозяйства. Такое растение может дать лучший урожай на ферме, но, скорее всего, ему будет трудно выжить в дикой природе, что делает его использование открытым для неправильной интерпретации. В генетике человека любой вопрос о «генетическом превосходстве» является еще более проблематичным из-за исторических и политических последствий любого такого утверждения. Некоторые могут даже зайти так далеко, что охарактеризуют это как сомнительное оценочное суждение в сфере политики, а не науки. [ 33 ]
- не все гибриды проявляют гетерозис (см. аутбридинговую депрессию ).
Примером неоднозначных оценочных суждений, навязываемых гибридам и гибридной силе, является мул . Хотя мулы почти всегда бесплодны, их ценят за сочетание выносливости и темперамента, которое отличается от их родителей-лошадей или ослов. Хотя эти качества могут сделать их «превосходными» для конкретных целей использования человеком, проблема бесплодия подразумевает, что эти животные, скорее всего, вымерли бы без вмешательства человека посредством животноводства , что сделает их «неполноценными» с точки зрения естественного отбора .
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Шнабле, Патрик С.; Спрингер, Натан М. (29 апреля 2013 г.). «Прогресс в понимании гетерозиса сельскохозяйственных растений» . Ежегодный обзор биологии растений . 64 (1): 71–88. doi : 10.1146/annurev-arplant-042110-103827 . ISSN 1543-5008 . ПМИД 23394499 .
- ^ «Засухоустойчивые гибридные семена дают фермерам отсрочку от голода» . СИММИТ . 06.03.2019 . Проверено 19 января 2023 г.
- ^ Джордж Харрисон Шулл (1948). «Что такое гетерозис?» . Генетика . 33 (5): 439–446. дои : 10.1093/генетика/33.5.439 . ПМК 1209417 . ПМИД 17247290 .
- ^ Эдмандс, Сюзанна (15 ноября 2006 г.). «Между молотом и наковальней: оценка относительных рисков инбридинга и аутбридинга для сохранения и управления: ОТНОСИТЕЛЬНЫЕ РИСКИ ИНБРИДИНГА И АУТБРЕДИНГА» . Молекулярная экология . 16 (3): 463–475. дои : 10.1111/j.1365-294X.2006.03148.x . ПМИД 17257106 . S2CID 457825 .
- ^ Jump up to: а б Ни З., Ким ЭД, Ха М и др. (январь 2009 г.). «Измененные циркадные ритмы регулируют силу роста у гибридов и аллополиплоидов» . Природа . 457 (7227): 327–31. Бибкод : 2009Natur.457..327N . дои : 10.1038/nature07523 . ПМЦ 2679702 . ПМИД 19029881 .
- ^ Jump up to: а б с д Баранвал В.К., Миккилинени В., Зер У.Б., Тьяги А.К., Капур С. (ноябрь 2012 г.). «Гетеозис: новые идеи о гибридной силе» . Дж. Эксп. Бот 63 (18): 6309–14. дои : 10.1093/jxb/ers291 . ПМИД 23095992 .
- ^ Кроу, Джеймс Ф. (1948). «Альтернативные гипотезы гибридной силы» . Генетика . 33 (5): 477–487. дои : 10.1093/генетика/33.5.477 . ПМК 1209419 . ПМИД 17247292 .
- ^ Давенпорт CB (1908). «Дегенерация, альбинизм и инбридинг» . Наука . 28 (718): 454–5. Бибкод : 1908Sci....28..454D . дои : 10.1126/science.28.718.454-b . ПМИД 17771943 . S2CID 29068462 .
- ^ Jump up to: а б Карр, Дэвид Э.; Дудаш, Мишель Р. (29 июня 2003 г.). «Современные подходы к генетической основе инбредной депрессии у растений» . Философские труды Королевского общества B: Биологические науки . 358 (1434): 1071–1084. дои : 10.1098/rstb.2003.1295 . ISSN 0962-8436 . ПМК 1693197 . ПМИД 12831473 .
- ^ Восток ЭМ (1908 г.). «Инбридинг кукурузы». Отчеты Сельскохозяйственной экспериментальной станции Коннектикута за 1907 год : 419–428.
- ^ Шулль Г.Х. (1908). «Композиция кукурузного поля». Отчеты Американской ассоциации заводчиков : 296–301.
- ^ Бирхлер Дж.А.; Оже ДЛ; Риддл, Северная Каролина (2003). «В поисках молекулярных основ гетерозиса» . Растительная клетка . 15 (10): 2236–2239. дои : 10.1105/tpc.151030 . ПМК 540269 . ПМИД 14523245 .
- ^ Jump up to: а б Кроу, Джеймс Ф. (1998). «90 лет назад: начало гибридной кукурузы» . Генетика . 148 (3): 923–928. дои : 10.1093/генетика/148.3.923 . ПМК 1460037 . ПМИД 9539413 .
- ^ Хан З., Мтанго Н.Р., Патель Б.Г., Сапиенца С., Латам К.Э. (октябрь 2008 г.). «Гибридная энергия и трансгенерационное эпигенетическое воздействие на ранний фенотип эмбриона мыши» . Биол. Репродукция . 79 (4): 638–48. дои : 10.1095/biolreprod.108.069096 . ПМЦ 2844494 . ПМИД 18562704 .
- ^ Чжоу Ю, Фергюсон Дж, Чанг Дж. Т., Клюгер Ю (2007). «Интер- и внутрикомбинаторная регуляция факторами транскрипции и микроРНК» . БМК Геномика . 8 : 396. дои : 10.1186/1471-2164-8-396 . ПМК 2206040 . ПМИД 17971223 .
- ^ Jump up to: а б Форт, Антуан; Райдер, Питер; Маккеун, Питер С.; Вейнен, Крис; Аартс, Марк Г.; Сюльпис, Ронан; Спиллейн, Чарльз (1 января 2016 г.). «Дезагрегация полиплоидии, дозировка родительского генома и вклад гибридности в гетерозис у Arabidopsis thaliana» . Новый фитолог . 209 (2): 590–599. дои : 10.1111/nph.13650 . ISSN 1469-8137 . ПМИД 26395035 .
- ^ Душинская, Дорота; Маккеун, Питер С.; Юнгер, Томас Э.; Петрашевская-Богель, Анна; Гилен, Дэнни; Спиллейн, Чарльз (1 апреля 2013 г.). «Плодородность гамет и изменение количества семяпочек у самоопыляющихся реципрокных гибридных триплоидных растений F1 передаются по наследству и демонстрируют эпигенетические эффекты родительского происхождения» . Новый фитолог . 198 (1): 71–81. дои : 10.1111/nph.12147 . ISSN 1469-8137 . ПМИД 23368793 .
- ^ Ву, Шеллен X. (01 июля 2021 г.). «Юань Лунпин (1930–2021)» . Природа . 595 (7865): 26. Бибкод : 2021Natur.595...26W . дои : 10.1038/d41586-021-01732-2 . ISSN 0028-0836 . S2CID 235633772 .
- ^ Jump up to: а б с д Ляо, Чанкан; Ян, Вэй; Чен, Чжуфэн; Се, Банда; Дэн, Син Ван; Тан, Сяоянь (июнь 2021 г.). «Инновации и развитие технологии гибридного риса третьего поколения» . Журнал «Урожай» . 9 (3): 693–701. doi : 10.1016/j.cj.2021.02.003 . S2CID 233623160 .
- ^ Jump up to: а б с Ким, Ю-Джин; Чжан, Дабин (январь 2018 г.). «Молекулярный контроль мужской фертильности для селекции гибридов сельскохозяйственных культур» . Тенденции в науке о растениях . 23 (1): 53–65. doi : 10.1016/j.tplants.2017.10.001 . ПМИД 29126789 .
- ^ О'Нил, генеральный директор; Церковь, ДБ; МакГриви, доктор медицинских наук; Томсон, ПК; Бродбелт, округ Колумбия (2013). «Продолжительность жизни и смертность домашних собак в Англии» (PDF) . Ветеринарный журнал . 198 (3): 638–43. дои : 10.1016/j.tvjl.2013.09.020 . ПМИД 24206631 .
- ^ Спейди, Тайрон С.; Острандер, Элейн А. (2008). «Поведенческая генетика собак: выявление фенотипов и определение генов» . Американский журнал генетики человека . 82 (1): 10–8. дои : 10.1016/j.ajhg.2007.12.001 . ПМК 2253978 . ПМИД 18179880 .
- ^ Джон Пол Скотт и Джон Л. Фуллер. «Генетика и социальное поведение собаки» . 1965. с. 307 и с. 313.
- ^ Пер Дженсен. «Поведенческая биология собак» . 2007. с. 179
- ^ Николас, Фрэнк В. (2016). «Гибридная сила у собак?» . Ветеринарный журнал . 214 : 77–83. дои : 10.1016/j.tvjl.2016.05.013 . ПМИД 27387730 . Проверено 29 июля 2020 г.
- ^ Николас, JAC (2012). «Обзор офтальмологических отклонений у лабрадуделя в Великобритании» . Ветеринарный журнал . 170 (15): 390. дои : 10.1136/vr.100361 . ПМИД 22278634 . S2CID 5932838 . Проверено 29 июля 2020 г.
- ^ Шарки, Лаура (2020). «Гибридная энергия у собак» . Функциональный собачий коллектив . Проверено 29 июля 2020 г.
- ^ Australian Geographic (сентябрь 2014 г.). «Гибридные птицы лучше борются с болезнями, чем чистокровные» .
- ^ Хоукс, Джон (2013). Значение геномов неандертальцев и денисовцев в эволюции человека . Том. 42. Ежегодные обзоры. стр. 433–449, 438. doi : 10.1146/annurev-anthro-092412-155548 . ISBN 978-0-8243-1942-7 . ISSN 0084-6570 .
Общая эволюционная история живых людей привела к высокому родству среди всех ныне живущих людей, о чем свидетельствует, например, очень низкий индекс фиксации (FST ) среди ныне живущих человеческих популяций.
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Барбуджани, Гвидо ; Колонна, Винченца (15 сентября 2011 г.). «Генетическая основа биоразнообразия человека: обновленная информация» . В Захосе, Фрэнк Э.; Хабель, Ян Кристиан (ред.). Горячие точки биоразнообразия . Спрингер. стр. 97–119. дои : 10.1007/978-3-642-20992-5_6 . ISBN 978-3-642-20992-5 . Проверено 23 ноября 2013 г.
Масштабные усилия по детальному изучению генома человека позволили получить колоссальное количество генетических данных. Хотя мы до сих пор не можем понять молекулярные основы большинства сложных признаков, включая многие распространенные заболевания, теперь у нас есть более четкое представление о степени генетического сходства между человеком и другими видами приматов. Мы также знаем, что люди генетически очень близки друг к другу, даже больше, чем любые другие приматы, что большая часть нашего генетического разнообразия объясняется индивидуальными различиями внутри популяций и что лишь небольшая часть генетической изменчивости видов приходится на разные популяции. и их географические группы.
- ^ Рамачандран, Сохини ; Тан, Хуа; Гутенкунст, Райан Н.; Бустаманте, Чарльз Д. (2010). «Генетика и геномика структуры человеческой популяции». В Спайчере, Майкл Р.; Антонаракис, Стилианс Э.; Мотульский, Арно Г. (ред.). Генетика человека Фогеля и Мотульского . Гейдельберг: Springer Scientific. стр. 100-1 589–615. дои : 10.1007/978-3-540-37654-5_22 . ISBN 978-3-540-37653-8 . S2CID 89318627 .
Большинство исследований популяционной генетики человека начинаются со цитирования основополагающей статьи Ричарда Левонтина 1972 года, носящей название этого подраздела [29]. Учитывая центральную роль, которую эта работа сыграла в нашей области, мы начнем с ее краткого обсуждения и вернемся к ее выводам на протяжении всей главы. ... Ключевой вывод статьи заключается в том, что 85,4% всех наблюдаемых генетических вариаций приходится на каждую группу. То есть он сообщил, что подавляющее большинство генетических различий обнаруживается внутри популяций, а не между ними. ... Его открытие воспроизводится в исследовании за исследованием вплоть до настоящего времени: два случайных человека из любой группы (которая может быть континентом или даже местным населением) почти так же различны, как любые два случайных человека со всего мира.
- ^ Камин, Леон Дж. (1980). «Инбредная депрессия и IQ» . Психологический вестник . 87 (3): 469–478. дои : 10.1037/0033-2909.87.3.469 . ISSN 1939-1455 . ПМИД 7384341 .
- ^ Jump up to: а б Риш Н., Бурхард Э., Зив Э., Тан Х (июль 2002 г.). «Категоризация людей в биомедицинских исследованиях: гены, раса и болезни» . Геном Биол . 3 (7): комментарий 2007. doi : 10.1186/gb-2002-3-7-comment2007 . ПМК 139378 . ПМИД 12184798 .
- ^ Веллер С.Г., Сакаи А.К., Тай Д.А., Том Дж., Рэнкин А.Е. (ноябрь 2005 г.). «Инбредная депрессия и гетерозис в популяциях Schiedea viscosa, очень самоопыляющегося вида» . Дж. Эвол. Биол . 18 (6): 1434–44. дои : 10.1111/j.1420-9101.2005.00965.x . ПМИД 16313456 .
Дальнейшее чтение
[ редактировать ]- Баккер, Винфридус (2006). «Повышенная жизнеспособность гибридов приносит пользу племенному животноводству и бройлерам» (PDF) . Кобб Фокус (2). Архивировано из оригинала (PDF) 17 декабря 2008 г.
- Бирчлер Дж. А., Оже Д. Л., Риддл, Северная Каролина (октябрь 2003 г.). «В поисках молекулярной основы гетерозиса» . Растительная клетка . 15 (10): 2236–9. дои : 10.1105/tpc.151030 . ПМК 540269 . ПМИД 14523245 .
- Техническая записка NOAA NMFS NWFSC-30: Генетические последствия заброса неместных заводских рыб в естественные популяции: инбредная депрессия и аутбредная депрессия
- Мак Грегор, SE (2009) [1976]. «Введение: Гибридная сила растений и ее связь с опылением насекомыми» (PDF) . Опыление культурных растений насекомыми . Справочник по сельскому хозяйству №. 496. Служба сельскохозяйственных исследований, USDA OCLC 243509268 .
- «Гибриды и семейные реликвии» Университета Иллинойса. — статья из журнала Home Hort Hints
- Ройбал, Дж. (1 июля 1998 г.). « Ранчстар ». Говядина (beefmagazine.com).
- «Секс-Линкс» — по птицеводству; на FeatherSite