Рибофлавинсинтаза
| Рибофлавинсинтаза | |||
|---|---|---|---|
 | |||
| Идентификаторы | |||
| Номер ЕС. | 2.5.1.9 | ||
| Номер CAS. | 9075-82-5 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
| 6,7-диметил-8-рибитиллумазинсинтаза | |||
|---|---|---|---|
 | |||
| Идентификаторы | |||
| Символ | DMRL_синтаза | ||
| Пфам | PF00885 | ||
| ИнтерПро | ИПР002180 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 1рвв / СКОПе / СУПФАМ | ||
| |||
Рибофлавинсинтаза — фермент конечную , катализирующий реакцию рибофлавина биосинтеза . Он катализирует перенос четырехуглеродного звена с одной молекулы 6,7-диметил-8-рибитиллумазина на другую, что приводит к синтезу рибофлавина и 5-амино-6-рибитиламино-2,4( 1H , 3H )-пиримидиндион :
- (2) 6,7-диметил-8-рибитиллумазин → рибофлавин + 5-амино-6-рибитиламино-2,4(1H , 3H ) -пиримидиндион
Структура
[ редактировать ]Мономер рибофлавинсинтазы имеет молекулярную массу около 23 кДа. Каждый мономер содержит два бета-цилиндра и одну α-спираль на С-конце (остатки 186–206). Мономер сворачивается в псевдодвойную симметрию, что предсказывается сходством последовательностей между стволом N-конца (остатки 4-86) и стволом C-конца (остатки 101-184). [ 1 ] Граница между этими стволами двух разных субъединиц и есть место активного сайта . [ 3 ] Фермент разных видов принимает разные четвертичные структуры , содержащие до 120 субъединиц. [ 4 ]
Архейная рибофлавинсинтаза образуется в виде гомопентамера, тогда как эубактериальная , грибковая и растительная рибофлавинсинтаза существует в виде гомотримера. Их последовательности совершенно неродственны, архейный фермент паралогичен 6,7 -диметил-8-рибитиллумазинсинтазе . [ 3 ] Реакции, катализируемые этими двумя типами рибофлавинсинтазы, протекают через «энантиомерные» промежуточные продукты. [ 3 ]
Активный сайт
[ редактировать ]Две молекулы 6,7-диметил-8-рибитиллумазина (синтезированные люмазинсинтазой ) связаны водородом с каждым мономером, поскольку эти два домена топологически схожи. [ 5 ] Активный центр расположен на границе раздела подложек активного центра . моделируемые структуры димера между парами мономеров, и созданы [ 2 ] Только один из активных центров фермента катализирует образование рибофлавина одновременно, тогда как два других сайта обращены наружу и подвергаются воздействию растворителя . [ 1 ] 62 , Ala64 , Изображены аминокислотные остатки, участвующие в образовании водородной связи с лигандом, участвующие остатки могут включать Thr148, Met160, Ile162, Thr165, Val6, Tyr164, Ser146 и Gly96 в C-концевом домене, а также Ser41, Thr50, Gly Ser64, Val103, Cys48, His102 в N-концевом домене. [ 3 ]
-
![Водородная связь между субстратом и ферментом в С-концевом домене.[2]](//upload.wikimedia.org/wikipedia/commons/thumb/a/ab/Hydrogen_Bonding_C-Terminal_Domain.jpg/120px-Hydrogen_Bonding_C-Terminal_Domain.jpg) Водородная связь между субстратом и ферментом в С-концевом домене. [ 2 ]
Водородная связь между субстратом и ферментом в С-концевом домене. [ 2 ] -
![Водородная связь между субстратом и ферментом в N-концевом домене.[2]](//upload.wikimedia.org/wikipedia/commons/thumb/1/14/Hydrogen_Bonding_N-Terminal_Domain.jpg/120px-Hydrogen_Bonding_N-Terminal_Domain.jpg) Водородная связь между субстратом и ферментом в N-концевом домене. [ 2 ]
Водородная связь между субстратом и ферментом в N-концевом домене. [ 2 ]
![Водородная связь между субстратом и ферментом в С-концевом домене.[2]](http://upload.wikimedia.org/wikipedia/commons/thumb/a/ab/Hydrogen_Bonding_C-Terminal_Domain.jpg/120px-Hydrogen_Bonding_C-Terminal_Domain.jpg)
![Водородная связь между субстратом и ферментом в N-концевом домене.[2]](http://upload.wikimedia.org/wikipedia/commons/thumb/1/14/Hydrogen_Bonding_N-Terminal_Domain.jpg/120px-Hydrogen_Bonding_N-Terminal_Domain.jpg)
Механизм
[ редактировать ]Для катализа не кофакторы требуются . Кроме того, образование рибофлавина из 6,7-диметил-8-рибитиллумазина может происходить в кипящем водном растворе в отсутствие рибофлавинсинтазы. [ 6 ] Реакция следующая:
- (2) 6,7-диметил-8-рибитиллумазин → рибофлавин + 5-амино-6-рибитиламино-2,4(1H , 3H ) -пиримидиндион
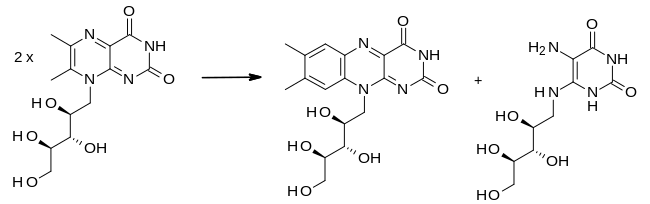

На границе раздела субстрата между парами мономеров фермент удерживает две молекулы 6,7-диметил-8-рибитиллумазина в положении посредством водородных связей, катализируя реакцию дисмутации . [ 6 ] Кроме того, был предложен кислотно-основной катализ аминокислотными остатками. His102/Thr148 Конкретные остатки могут включать диаду в качестве основания для депротонирования метильной группы C7a . В диаде His102 происходит из N-цилиндра, а Thr148 - из C-цилиндра, что подчеркивает важность близости двух субъединиц фермента на ранних стадиях реакции. [ 7 ] Также было высказано предположение, что нуклеофилом является один из следующих консервативных остатков: Ser146, Ser41, Cys48 или Thr148 или вода в некатализируемой реакции. [ 1 ] В исследованиях роли Cys48 как возможного нуклеофила не было определено, происходит ли нуклеофильное замещение посредством реакции SN1 или SN2 . [ 7 ]
Во время реакции дисмутации между двумя молекулами 6,7-диметил-8-рибитиллумазина происходит обмен четырехуглеродной единицы. В ходе реакции образуется пентациклическая молекула, которая затем распадается на рибофлавин и 5-амино-6-рибитиламино-2,4(1H , 3H ) -пиримидиндион. Интересно, что архейная рибофлавинсинтаза создает «энантиомерный» промежуточный продукт в отличие от эубактериальной, грибковой и растительной рибофлавинсинтазы, где атака одной молекулы происходит с противоположной стороны по сравнению с другим ферментом. [ 3 ] Точный механизм образования циклического аддукта неизвестен. Также неизвестно, как именно протекает этот механизм, если он не катализируется ферментом. [ 3 ]
Производство лекарств
[ редактировать ]Ученые выдвинули гипотезу, что ферменты, участвующие в пути биосинтеза рибофлавина, в том числе рибофлавинсинтаза, могут быть использованы для разработки антибактериальных препаратов для лечения инфекций, вызванных грамотрицательными бактериями и дрожжами . Эта гипотеза основана на неспособности грамотрицательных бактерий, таких как E. coli и S. typhimurium , поглощать рибофлавин из внешней среды. [ 3 ] [ 8 ] Поскольку грамотрицательным бактериям необходимо вырабатывать собственный рибофлавин, ингибирование рибофлавинсинтазы или других ферментов, участвующих в этом пути, может быть полезным инструментом при разработке антибактериальных препаратов.
рибофлавинсинтазы Наиболее мощным ингибитором является 9- D- рибитил-1,3,7-тригидропурин-2,6,8-трион со K i значением 0,61 мкМ. Считается, что он действует посредством конкурентного ингибирования с помощью 6,7-диметил-8-рибитиллумазина. [ 8 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д ПДБ : 1i8d ; Ляо Д.И., Ваврзак З., Калабрезе Х.К., Виитанен П.В., Джордан Д.Б. (май 2001 г.). «Кристаллическая структура рибофлавинсинтазы» . Структура . 9 (5): 399–408. дои : 10.1016/S0969-2126(01)00600-1 . ПМИД 11377200 .
- ^ Перейти обратно: а б с д ПДБ : 1кзл ; Герхардт С., Шотт А.К., Кайрис Н., Кушман М., Илларионов Б., Эйзенрайх В., Бахер А., Хубер Р., Штайнбахер С., Фишер М. (октябрь 2002 г.). «Исследование механизма реакции рибофлавинсинтазы: рентгенокристаллическая структура комплекса с 6-карбоксиэтил-7-оксо-8-рибитиллумазином» . Структура . 10 (10): 1371–81. дои : 10.1016/S0969-2126(02)00864-X . ПМИД 12377123 .
- ^ Перейти обратно: а б с д и ж г Фишер М., Бахер А. (июнь 2008 г.). «Биосинтез витамина B2: Структура и механизм рибофлавинсинтазы». Арх. Биохим. Биофиз . 474 (2): 252–65. дои : 10.1016/j.abb.2008.02.008 . ПМИД 18298940 .
- ^ «ЭК 2.5.1.9» . ebi.ac.uk. Проверено 28 ноября 2021 г.
- ^ Фишер М., Шотт А.К., Кемтер К., Файхт Р., Рихтер Г., Илларионов Б., Эйзенрайх В., Герхардт С., Кушман М., Штайнбахер С., Хубер Р., Бахер А. (декабрь 2003 г.). «Рибофлавинсинтаза Schizosaccharomyces pombe. Динамика белков, выявленная с помощью экспериментов по возмущению белков ЯМР 19F» . БМК Биохим . 4:18 . дои : 10.1186/1471-2091-4-18 . ПМК 337094 . ПМИД 14690539 .
- ^ Перейти обратно: а б Бахер А., Эберхардт С., Фишер М., Кис К., Рихтер Г. (2000). «Биосинтез витамина В2 (рибофлавина)». Анну. Преподобный Нутр . 20 : 153–67. дои : 10.1146/annurev.nutr.20.1.153 . ПМИД 10940330 .
- ^ Перейти обратно: а б Чжэн Ю.Дж., Джордан Д.Б., Ляо Ди (август 2003 г.). «Исследование промежуточного продукта реакции в активном центре рибофлавинсинтазы». Биоорг. Хим . 31 (4): 278–87. дои : 10.1016/S0045-2068(03)00029-4 . ПМИД 12877878 .
- ^ Перейти обратно: а б Кушман М., Ян Д., Кис К., Бахер А. (декабрь 2001 г.). «Разработка, синтез и оценка 9-D-рибитил-1,3,7-тригидро-2,6,8-пуринетриона, мощного ингибитора рибофлавинсинтазы и люмазинсинтазы». Дж. Орг. Хим . 66 (25): 8320–7. дои : 10.1021/jo010706r . ПМИД 11735509 .
Внешние ссылки
[ редактировать ]- Рибофлавин + синтаза Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)