переворот оснований ДНК
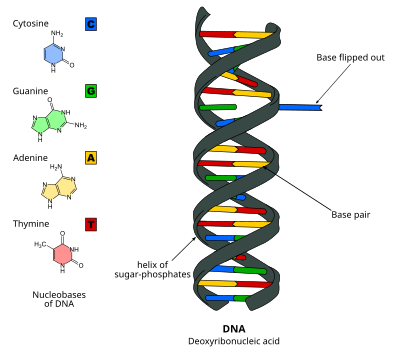
Переворот оснований ДНК , или переворот нуклеотидов , представляет собой механизм, при котором одно нуклеотидное основание, или нуклеиновое основание , поворачивается за пределы двойной спирали нуклеиновой кислоты. [ 1 ] , обрабатывающему нуклеиновую кислоту, Это происходит, когда ферменту необходим доступ к основанию для выполнения работы над ним, например, его вырезание для замены другим основанием во время репарации ДНК . Впервые это было обнаружено в 1994 году с помощью рентгеновской кристаллографии фермента метилтрансферазы , катализирующего метилирование цитозинового основания в ДНК. С тех пор было показано, что он используется различными ферментами во многих биологических процессах, таких как метилирование ДНК , различные механизмы репарации ДНК и репликация ДНК . Это также может происходить в двойных спиралях РНК. [ 2 ] или в промежуточных продуктах ДНК:РНК, образующихся во время транскрипции РНК .
Переворот оснований ДНК происходит путем разрыва водородных связей между основаниями и отделения основания от соседей. Это может происходить посредством активного процесса, когда фермент связывается с ДНК и затем способствует вращению основания, или пассивного процесса, когда основание самопроизвольно вращается, и это состояние распознается и связывается ферментом. Его можно обнаружить с помощью Рентгеновская кристаллография , ЯМР-спектроскопия , флуоресцентная спектроскопия или гибридизационные зонды .
Открытие
[ редактировать ]Переворот оснований впервые наблюдался в 1994 году, когда исследователи Климасаускас, Кумар, Робертс и Ченг использовали рентгеновскую кристаллографию, чтобы увидеть промежуточную стадию химической реакции метилтрансферазы, связанной с ДНК . [ 3 ] В качестве метилтрансферазы они использовали C5-цитозинметилтрансферазу Haemophilus haemolyticus (M. HhaI). Этот фермент распознает специфическую последовательность ДНК (5'-GCGC-3') и метилирует первое цитозиновое основание последовательности в ее положении C5. [ 3 ] При кристаллизации комплекса M. HhaI-ДНК они увидели, что целевое цитозиновое основание полностью вывернулось из двойной спирали и расположилось в активном сайте M. HhaI. Он удерживался на месте благодаря многочисленным взаимодействиям между M. HhaI и ДНК. [ 3 ]
Авторы предположили, что переворот оснований является механизмом, используемым многими другими ферментами, такими как хеликазы , ферменты рекомбинации , РНК-полимеразы , ДНК-полимеразы и топоизомеразы типа II . [ 3 ] За годы, прошедшие после этого открытия, было проведено много исследований, и было обнаружено, что переворот оснований — это механизм, используемый во многих биологических процессах, которые предлагают авторы. [ 4 ] [ 5 ] [ 6 ]
Механизм
[ редактировать ]
Нуклеотиды ДНК удерживаются вместе водородными связями , которые относительно слабы и легко разрываются. Переворот базы происходит за миллисекунды. [ 7 ] разрывая водородные связи между основаниями и отделяя основание от соседей. [ 8 ] Основание повернуто из двойной спирали на 180 градусов. [ 9 ] обычно через большую канавку , [ 10 ] и в активный центр фермента. Это открытие приводит к небольшим конформационным изменениям в остове ДНК. [ 11 ] которые быстро стабилизируются за счет увеличения взаимодействия фермента с ДНК. [ 12 ] Исследования, изучающие профили свободной энергии при перевороте оснований, показали, что барьер свободной энергии для переворота может быть снижен на 17 ккал/моль для M.HhaI в закрытой конформации . [ 10 ]
Существует два механизма переворота оснований ДНК: активный и пассивный. [ 13 ] При активном механизме фермент связывается с ДНК, а затем активно вращает основание, тогда как при пассивном механизме поврежденное основание сначала самопроизвольно вращается, а затем распознается и связывается ферментом. [ 8 ] Исследования продемонстрировали оба механизма: урацил-ДНК-гликозилаза действует по пассивному механизму. [ 8 ] а Tn10 транспозаза следует активному механизму. [ 14 ]
Кроме того, исследования показали, что переворот оснований ДНК используется многими различными ферментами в различных биологических процессах, таких как метилирование ДНК , различные репарации ДНК механизмы , транскрипция РНК и репликация ДНК . [ 4 ] [ 5 ] [ 6 ]
Биологические процессы
[ редактировать ]Модификация и репарация ДНК
[ редактировать ]
ДНК может иметь мутации , которые приводят к повреждению основания цепи ДНК. Чтобы обеспечить генетическую целостность ДНК, ферментам необходимо восстанавливать любые повреждения. Существует много типов репарации ДНК . При восстановлении иссечения основания используется переворот основания, чтобы вывернуть поврежденное основание из двойной спирали. [ 5 ] и в карман специфичности гликозилазы , которая гидролизует гликозидную связь и удаляет основание. [ 15 ] ДНК-гликозилазы взаимодействуют с ДНК, переворачивая основания для определения несоответствия. Примером эксцизионной репарации оснований является ситуация, когда цитозиновое основание дезаминируется и становится основанием урацила. Это вызывает неправильное спаривание U:G, которое обнаруживается урацил-ДНК-гликозилазой . Основание урацила переворачивается в активный гликозилазный карман, где оно удаляется из цепи ДНК. [ 16 ] Переворот оснований используется для восстановления таких мутаций, как 8-оксогуанин (oxoG). [ 17 ] и димеры тимина, создаваемые УФ-излучением. [ 15 ] [ 18 ]
Репликация, транскрипция и рекомбинация
[ редактировать ]Репликация ДНК и транскрипция РНК используют переворот оснований. [ 5 ] ДНК-полимераза – это фермент, осуществляющий репликацию. Его можно рассматривать как руку, сжимающую одноцепочечную матрицу ДНК. [ 15 ] Когда матрица проходит через область ладони полимеразы, основания матрицы выворачиваются из спирали и от места связывания dNTP . [ 19 ] Во время транскрипции РНК-полимераза катализирует синтез РНК. Во время фазы инициации два основания элемента -10 выскакивают из спирали и попадают в два кармана РНК-полимеразы. Эти новые взаимодействия стабилизируют элемент -10 и способствуют разделению или плавлению нитей ДНК. [ 15 ] [ 20 ]
Переворот оснований происходит на последних стадиях рекомбинации . [ 21 ] RecA — белок, который способствует инвазии цепи [ 15 ] в ходе гомологичной рекомбинации . Переворот оснований был предложен как механизм, с помощью которого RecA может позволить одной цепи распознавать гомологию в дуплексной ДНК. [ 22 ] Другие исследования показывают, что он также участвует в рекомбинации V(D)J . [ 23 ]
Метилирование ДНК
[ редактировать ]
Метилирование ДНК — это процесс, при котором метильная группа добавляется либо к цитозину , либо к аденину . [ 24 ] Этот процесс вызывает активацию или инактивацию экспрессии генов , что приводит к регуляции генов в эукариотических клетках. Известно также, что процесс метилирования ДНК участвует в формировании некоторых типов рака . [ 25 ] [ 26 ] [ 27 ] Для того чтобы произошла эта химическая модификация, необходимо, чтобы целевое основание вывернулось из двойной спирали ДНК, чтобы позволить метилтрансферазам катализировать реакцию. [ 5 ]
Распознавание цели эндонуклеазами рестрикции
[ редактировать ]Эндонуклеазы рестрикции, также известные как ферменты рестрикции , представляют собой ферменты, которые расщепляют сахарофосфатный остов ДНК по определенным нуклеотидным последовательностям, длина которых обычно составляет от четырех до шести нуклеотидов. [ 28 ] Исследования, проведенные Хортоном и его коллегами, показали, что механизм, с помощью которого эти ферменты расщепляют ДНК, включает переворот оснований, а также изгиб ДНК и расширение малой бороздки . [ 29 ] В 2006 году Хортон и его коллеги были представлены данные рентгеновской кристаллографии , показывающие, что эндонуклеаза рестрикции HinP1I использует переворот оснований, чтобы распознать свою целевую последовательность. Известно, что этот фермент расщепляет ДНК по палиндромной тетрануклеотидной последовательности G↓CGC.
Экспериментальные подходы к обнаружению
[ редактировать ]Рентгеновская кристаллография
[ редактировать ]
Рентгеновская кристаллография — это метод измерения углов и интенсивности кристаллических атомов с целью определения атомной и молекулярной структуры интересующего кристалла. Кристаллографы затем могут создать трехмерное изображение, на котором можно определить положения атомов , химические связи , а также другие важные характеристики. [ 30 ] Климасаукас и его коллеги использовали эту технику для наблюдения первого явления переворота базы, при этом их экспериментальная процедура включала несколько этапов: [ 3 ]
- Очистка
- Кристаллизация
- Сбор данных
- Определение и уточнение структуры
Haemophilus haemolyticus Во время очистки метилтрансфераза была сверхэкспрессирована и очищена с использованием стадии обратной экстракции высоким содержанием соли для селективной солюбилизации M.HhaI с последующей быстрой белковой жидкостной хроматографией ( FPLC ), как это было сделано ранее Кумаром и его коллегами. [ 31 ] Авторы использовали анионообменную колонку Mono-Q для удаления небольшого количества белковых материалов и нежелательной ДНК перед стадией кристаллизации. После успешной очистки M.HhaI образец выращивали с использованием метода смешивания раствора, содержащего комплекс, при температуре 16 °C и метода диффузии пара в виде висячей капли для получения кристаллов. Затем авторы смогли собрать рентгеновские данные в соответствии с методом, использованным Ченгом и его коллегами в 1993 году. [ 32 ] Этот метод включал измерение интенсивности дифракции на детекторе FAST, где время экспозиции при повороте на 0,1° составляло 5 или 10 секунд. Для определения и уточнения структуры Климасаукас и его коллеги использовали молекулярную замену уточненной апо-структуры, описанную Ченгом и его коллегами в 1993 году. [ 32 ] где модели поиска X-PLOR , MERLOT и TRNSUM использовались для решения функций вращения и перевода. [ 33 ] [ 34 ] Эта часть исследования предполагает использование разнообразного программного обеспечения и компьютерных алгоритмов для определения структуры и характеристик интересующего кристалла.
ЯМР-спектроскопия
[ редактировать ]ЯМР-спектроскопия — это метод, который на протяжении многих лет использовался для изучения важных динамических аспектов переворота оснований. Этот метод позволяет исследователям определять физические и химические свойства атомов и других молекул, используя магнитные свойства атомных ядер . [ 35 ] Кроме того, ЯМР может предоставить разнообразную информацию, включая структуру, состояния реакции , химическое окружение молекул и динамику. [ 36 ] [ 37 ] Во время эксперимента по открытию переворота оснований ДНК исследователи использовали ЯМР-спектроскопию для изучения ферментативного переворота оснований метилтрансферазы HhaI. Для проведения этого эксперимента два остатка 5-фторцитозина были включены в мишень и контрольное положение с ДНК-субстратом, чтобы 19 Можно провести анализ химического сдвига F. Как только 19 Был проведен анализ химического сдвига F , затем был сделан вывод, что комплексы ДНК существуют с несколькими формами целевого 5-фторцитозина по пути переворота оснований. [ 38 ]
Флуоресцентная спектроскопия
[ редактировать ]Флуоресцентная спектроскопия — это метод, который используется для анализа образца с помощью флуоресцентного зонда. Сами по себе нуклеотиды ДНК не являются хорошими кандидатами для этого метода, поскольку они с трудом переизлучают свет при световом возбуждении. [ 39 ] Для обнаружения переворота оснований необходим флуоресцентный маркер. 2-Аминопурин — это основание, которое структурно похоже на аденин , но очень флуоресцирует при высвобождении из дуплекса ДНК. [ 40 ] Он обычно используется для обнаружения переворота оснований и имеет возбуждение при 305–320 нм и эмиссию при 370 нм, поэтому он хорошо отделен от возбуждений белков и ДНК. Другими флуоресцентными зондами, используемыми для изучения переворота оснований ДНК, являются 6MAP (4-амино-6-метил-7(8H)-птеридон). [ 41 ] и пирроло-C (3-[β-D-2-рибофуранозил]-6-метилпирроло[2,3-d]пиримидин-2(3H)-он). [ 42 ] [ 43 ] Флуоресцентная спектроскопия с временным разрешением также используется для получения более подробной картины степени переворота оснований, а также конформационной динамики, происходящей во время переворота оснований. [ 44 ]
Исследование гибридизации
[ редактировать ]Зонды гибридизации можно использовать для обнаружения переворота оснований. В этом методе используется молекула, последовательность которой комплементарна той последовательности, которую вы хотите обнаружить, и которая связывается с одноцепочечной ДНК или РНК. Для обнаружения переворота оснований было использовано несколько зондов гибридизации. Перманганат калия используется для обнаружения остатков тимина , которые были выброшены метилтрансферазами цитозин-C5 и аденин-N6 . [ 45 ] Хлороацетальдегид используется для обнаружения остатков цитозина , выброшенных ДНК-цитозин-5-метилтрансферазой HhaI (M. HhaI). [ 46 ]

См. также
[ редактировать ]- восстановление ДНК
- Базовый иссеченный ремонт
- репликация ДНК
- Транскрипция РНК
- Метилирование ДНК
- ДНК-метилтрансфераза
- Генетическая рекомбинация
- Гомологичная рекомбинация
- ДНК
- Эпигенетика
- Эпигеномика
Ссылки
[ редактировать ]- ^ Робертс, Р.Дж.; Ченг, X (1998). «Переворот базы». Ежегодный обзор биохимии . 67 (1): 181–198. doi : 10.1146/annurev.biochem.67.1.181 . ПМИД 9759487 .
- ^ Рейтер, Нью-Джерси; Блад, Х; Абильдгаард, Ф; Мясник, ЮВ (2004). «Динамика внутримолекулярной стволовой петли U6 РНК: конформационное изменение с переворачиванием оснований». Биохимия . 43 (43): 13739–47. дои : 10.1021/bi048815y . ПМИД 15504036 . S2CID 25391616 .
- ^ Перейти обратно: а б с д и Климасаускас, Саулюс; Кумар, Санджай; Робертс, Ричард Дж.; Ченг, Сяодун (январь 1994 г.). «Hhal-метилтрансфераза выворачивает целевое основание из спирали ДНК». Клетка . 76 (2): 357–369. дои : 10.1016/0092-8674(94)90342-5 . ПМИД 8293469 . S2CID 23161543 .
- ^ Перейти обратно: а б Браун, Том. «Книга нуклеиновых кислот» . ATDBio . Проверено 26 февраля 2014 г.
- ^ Перейти обратно: а б с д и Хуанг, Ню; Нилеш К. Банавали; Александр Д. МакКерелл (7 января 2003 г.). «Облегченный белком переворот оснований в ДНК цитозин-5-метилтрансферазой» . ПНАС . 100 (1): 68–73. Бибкод : 2003PNAS..100...68H . дои : 10.1073/pnas.0135427100 . ПМК 140885 . ПМИД 12506195 .
- ^ Перейти обратно: а б Грубмюллер, Гельмут. «Переворот базы ДНК» . Архивировано из оригинала 4 февраля 2017 года . Проверено 26 февраля 2014 г.
- ^ Бувье, Бенджамин; Грубмюллер, Хельмут (август 2007 г.). «Молекулярно-динамическое исследование медленного переворота оснований в ДНК с использованием конформационного наводнения» (PDF) . Биофизический журнал . 93 (3): 770–786. Бибкод : 2007BpJ....93..770B . doi : 10.1529/biophysj.106.091751 . ЧВК 1913169 . ПМИД 17496048 . Архивировано из оригинала (PDF) 9 августа 2017 г. Проверено 15 марта 2014 г.
- ^ Перейти обратно: а б с Ларивьер, Л. (23 июня 2004 г.). «Структурные доказательства пассивного механизма переворота оснований глюкозилтрансферазы» . Журнал биологической химии . 279 (33): 34715–34720. дои : 10.1074/jbc.M404394200 . ПМИД 15178685 .
- ^ Грожан, [под редакцией] Анри (2009). Ферменты модификации ДНК и РНК: структура, механизм, функции и эволюция . Остин, Техас: Landes Bioscience. ISBN 978-1-58706-329-9 . Архивировано из оригинала 7 апреля 2014 г. Проверено 10 марта 2014 г.
{{cite book}}:|first=имеет общее имя ( справка ) - ^ Перейти обратно: а б Хуанг, Н.; Банавали, Северная Каролина; МакКерелл, AD (27 декабря 2002 г.). «Облегченный белком переворот оснований в ДНК цитозин-5-метилтрансферазой» . Труды Национальной академии наук . 100 (1): 68–73. дои : 10.1073/pnas.0135427100 . ПМК 140885 . ПМИД 12506195 .
- ^ Джудиче, Э. (1 марта 2003 г.). «Открытие пары оснований в B-ДНК: пути свободной энергии для пар GC и AT на основе моделирования зонтичного отбора проб» . Исследования нуклеиновых кислот . 31 (5): 1434–1443. дои : 10.1093/нар/gkg239 . ПМК 149832 . ПМИД 12595551 .
- ^ Хуанг, Н.; Банавали, Северная Каролина; МакКерелл, AD (27 декабря 2002 г.). «Облегченный белком переворот оснований в ДНК цитозин-5-метилтрансферазой» . Труды Национальной академии наук . 100 (1): 68–73. дои : 10.1073/pnas.0135427100 . ПМК 140885 . ПМИД 12506195 .
- ^ О'Нил, Лорен. Переворот оснований в ДНК: обнаружение, структура и энергетика: Диссертация . ISBN 9780549590743 . Проверено 15 марта 2014 г. [ постоянная мертвая ссылка ]
- ^ Бишерур, Жюльен; Чалмерс, Рональд; Белинский, Аня-Катрин (10 июля 2009 г.). «Переворот базы при транспозиции Tn10: механизм активного переворота и захвата» . ПЛОС ОДИН . 4 (7): е6201. Бибкод : 2009PLoSO...4.6201B . дои : 10.1371/journal.pone.0006201 . ПМК 2705183 . ПМИД 19593448 .
- ^ Перейти обратно: а б с д и Университет, Джеймс Д. Уотсон, Лаборатория Колд-Спринг-Харбор, Таня А. Бейкер, Массачусетский технологический институт, Александр Ганн, Лаборатория Колд-Спринг-Харбор, Майкл Левин, Калифорнийский университет, Беркли, Ричард Лосик, Гарвард (2014). Молекулярная биология гена (Седьмое изд.). Бостон: Пирсон/CSH Press. ISBN 978-0-321-76243-6 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Крокан, Ханс Э; Драблёс, Финн; Слупфауг, Гейр (16 декабря 2002 г.). «Урацил в ДНК – возникновение, последствия и восстановление». Онкоген . 21 (58): 8935–8948. дои : 10.1038/sj.onc.1205996 . ПМИД 12483510 .
- ^ Банерджи, Анирбан; Ян, Вэй; Карплюс, Мартин; Вердин, Грегори Л. (31 марта 2005 г.). «Структура репарационного фермента, опрашивающего неповрежденную ДНК, позволяет распознавать поврежденную ДНК». Природа . 434 (7033): 612–618. Бибкод : 2005Natur.434..612B . дои : 10.1038/nature03458 . ПМИД 15800616 . S2CID 4426014 .
- ^ Фуксрайтер, Моника; Ло, Нин; Едловский, Пал; Саймон, Иштван; Осман, Роман (ноябрь 2002 г.). «Роль переворота оснований в специфическом распознавании поврежденной ДНК репарирующими ферментами». Журнал молекулярной биологии . 323 (5): 823–834. дои : 10.1016/S0022-2836(02)00999-3 . ПМИД 12417196 .
- ^ Патель, Премал Х.; Сузуки, Мотоши; Адман, Элинор; Синкай, Акео; Леб, Лоуренс А. (май 2001 г.). «Прокариотическая ДНК-полимераза I: эволюция, структура и механизм «переворота оснований» для отбора нуклеотидов». Журнал молекулярной биологии . 308 (5): 823–837. дои : 10.1006/jmbi.2001.4619 . ПМИД 11352575 . S2CID 16277925 .
- ^ Лим, ХМ; Ли, HJ; Рой, С.; Адхья, С. (4 декабря 2001 г.). «Мастер» в распарывании оснований при изомеризации промотора при связывании РНК-полимеразы» . Труды Национальной академии наук . 98 (26): 14849–14852. Бибкод : 2001PNAS...9814849L . дои : 10.1073/pnas.261517398 . ПМК 64947 . ПМИД 11734629 .
- ^ Волошин Олег Н.; Камерини-Отеро, Р. Дэниел (сентябрь 2004 г.). «Возвращение к синаптическому комплексу» . Молекулярная клетка . 15 (6): 846–847. doi : 10.1016/j.molcel.2004.09.010 . ПМИД 15383274 .
- ^ Фольта-Стогнев, Э; О'Мэлли, С; Гупта, Р; Андерсон, Канзас; Раддинг, КМ (24 сентября 2004 г.). «Обмен парами оснований ДНК, который совпадает с распознаванием гомологии, поддерживаемой белком RecA E. coli» . Молекулярная клетка . 15 (6): 965–75. doi : 10.1016/j.molcel.2004.08.017 . ПМИД 15383285 .
- ^ Бишерур, Дж.; Лу, К.; Рот, Д.Б.; Чалмерс, Р. (31 августа 2009 г.). «Переворот оснований при рекомбинации V (D) J: понимание механизма образования шпильки, правила 12/23 и координации двухцепочечных разрывов» . Молекулярная и клеточная биология . 29 (21): 5889–5899. дои : 10.1128/MCB.00187-09 . ПМЦ 2772739 . ПМИД 19720743 .
- ^ Клозе, Роберт Дж.; Адриан П. Берд (2006). «Метилирование геномной ДНК: метка и ее медиаторы». Тенденции биохимических наук . 31 (2): 89–97. дои : 10.1016/j.tibs.2005.12.008 . ISSN 0968-0004 . ПМИД 16403636 .
- ^ Накао, М. (2001). «Эпигенетика: взаимодействие метилирования ДНК и хроматина». Джин . 278 (1–2): 25–31. дои : 10.1016/s0378-1119(01)00721-1 . ПМИД 11707319 .
- ^ Пласс, С; Солоуэй, П.Д. (2002). «Метилирование ДНК, импринтинг и рак» . Eur J Hum Genet . 10 (1): 6–16. дои : 10.1038/sj.ejhg.5200768 . ПМИД 11896451 .
- ^ Эстеллер, М; Герман, Дж.Г. (2002). «Рак как эпигенетическое заболевание: метилирование ДНК и изменения хроматина в опухолях человека». Дж. Патол . 196 (1): 1–7. дои : 10.1002/путь.1024 . ПМИД 11748635 . S2CID 35380651 .
- ^ «Биология и активность эндонуклеаз рестрикции» . Архивировано из оригинала 18 апреля 2014 г. Проверено 3 апреля 2014 г.
- ^ Хортон, Джон Р.; Чжан, Син; Маунус, Роберт; Ян, Чжэ; Уилсон, Джеффри; Робертс, Ричард; Ченг, Сяодун (2006). «Никкинг ДНК эндонуклеазой HinP1I: изгиб, переворачивание основания и незначительное расширение канавок» . Исследования нуклеиновых кислот . 34 (3): 939–948. дои : 10.1093/nar/gkj484 . ПМЦ 1363774 . ПМИД 16473850 .
- ^ Рентгеновская кристаллография
- ^ Кумар, С; Ченг, X; Пфлуграт, JW; Робертс, Р.Дж. (1992). «Очистка, кристаллизация и предварительный рентгеноструктурный анализ комплекса M.HhaI-AdoMet». Биохимия . 31 (36): 8648–8653. дои : 10.1021/bi00151a035 . ПМИД 1390649 .
- ^ Перейти обратно: а б Ченг, X; Кумар, С; Посфаи, Дж; Пфлуграт, JW; Робертс, Р.Дж. (1993). «Кристаллическая структура ДНК-метилтрансферазы HhaI в комплексе с S-аденозил-L-метионином». Клетка . 74 (2): 299–307. дои : 10.1016/0092-8674(93)90421-л . ПМИД 8343957 . S2CID 54238106 .
- ^ Brunger AT (1992) «X-PLOR, версия 3.1: система рентгеновской кристаллографии и ЯМР» (Нью-Хейвен, Коннектикут: издательство Йельского университета)
- ^ Фицджеральд, PMD (1988). «МЕРЛОТ, комплексный пакет компьютерных программ для определения кристаллической структуры методом молекулярного замещения». Дж. Прил. Кристаллогр . 21 (3): 273–288. дои : 10.1107/s0021889887012299 .
- ^ ЯМР-спектроскопия
- ^ Герон, М. и Дж. Лерой. 1995. Исследования кинетики пар оснований методом ЯМР-измерения протонного обмена. В ядерном магнитном резонансе и нуклеиновых кислотах. Academic Press, Сан-Диего, Калифорния.
- ^ Лейон, М.; Граслунд, А. (1992). «Влияние последовательности и длины на имино-протонный обмен и кинетику открытия пар оснований в дуплексах ДНК-олигонуклеотидов» . Нуклеиновые кислоты Рез . 20 (20): 5339–5343. дои : 10.1093/нар/20.20.5339 . ПМЦ 334339 . ПМИД 1331987 .
- ^ Климасаукас, Салюс и Зита Люткевичюте. «Экспериментальные подходы к изучению переворота оснований ДНК». Ферменты модификации ДНК и РНК: структура, механизм, функции и эволюция. Landes Bioscience, 2009. 37–50. Веб. 16 марта 2014 г. < https://www.landesbioscience.com/pdf/04GrosjeanKlimasauskas.pdf. Архивировано 7 апреля 2014 г. в Wayback Machine >.
- ^ Грожан, [под редакцией] Анри (2009). Ферменты модификации ДНК и РНК: структура, механизм, функции и эволюция (PDF) . Остин, Техас: Landes Bioscience. п. 43. ИСБН 978-1-58706-329-9 .
{{cite book}}:|first=имеет общее имя ( справка ) - ^ Хольц, Б. (15 февраля 1998 г.). «2-Аминопурин как флуоресцентный зонд для переворота оснований ДНК метилтрансферазами» . Исследования нуклеиновых кислот . 26 (4): 1076–1083. дои : 10.1093/нар/26.4.1076 . ПМК 147370 . ПМИД 9461471 .
- ^ Ян, К; Мацика, С; Стэнли, Р.Дж. (6 сентября 2007 г.). «6MAP, флуоресцентный аналог аденина, представляет собой зонд переворота оснований ДНК-фотолиазой». Журнал физической химии Б. 111 (35): 10615–25. дои : 10.1021/jp071035p . ПМИД 17696385 . S2CID 4998287 .
- ^ Ян, К; Стэнли, Р.Дж. (май – июнь 2008 г.). «Степень деформации ДНК в комплексах ДНК-фотолиаза-субстрат: исследование флуоресценции в состоянии раствора». Фотохимия и фотобиология . 84 (3): 741–9. дои : 10.1111/j.1751-1097.2007.00251.x . ПМИД 18086248 . S2CID 44506405 .
- ^ Берри, Дэвид А.; Юнг, Ки-Ён; Уайз, Дин С.; Серсель, Энтони Д.; Пирсон, Уильям Х.; Маки, Хью; Рэндольф, Джон Б.; Сомерс, Роберт Л. (март 2004 г.). «Пирроло-dC и пирроло-C: флуоресцентные аналоги цитидина и 2'-дезоксицитидина для изучения олигонуклеотидов». Буквы тетраэдра . 45 (11): 2457–2461. дои : 10.1016/j.tetlet.2004.01.108 .
- ^ Нили, Р.К.; Тамулайтис Г.; Чен, К.; Кубала, М.; Сикснис, В.; Джонс, AC (8 сентября 2009 г.). «Флуоресцентные исследования с временным разрешением переворота нуклеотидов ферментами рестрикции» . Исследования нуклеиновых кислот . 37 (20): 6859–6870. дои : 10.1093/нар/gkp688 . ПМК 2777440 . ПМИД 19740769 .
- ^ Серва, С. (1 августа 1998 г.). «Химическое отображение остатков тимина, вызванных ДНК-метилтрансферазами» . Исследования нуклеиновых кислот . 26 (15): 3473–3479. дои : 10.1093/нар/26.15.3473 . ПМЦ 147733 . ПМИД 9671807 .
- ^ Даухотит, Д.; Люткявичюте, З.; Тамулайтис Г.; Климасаускас, С. (15 апреля 2008 г.). «Химическое картирование цитозинов, ферментативно вывернутых из спирали ДНК» . Исследования нуклеиновых кислот . 36 (10): е57. дои : 10.1093/нар/gkn200 . ПМЦ 2425465 . ПМИД 18450817 .