Японский огнебрюхий тритон
| Японский огнебрюхий тритон Временной диапазон: Средний миоцен - настоящее время
| |
|---|---|

| |
| Самка японского огнебрюхого тритона | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Амфибия |
| Заказ: | Уродела |
| Семья: | Саламандриды |
| Род: | Кинопы |
| Разновидность: | C. пиррогастер
|
| Биномиальное имя | |
| Кинопс пиррогастер ( Бойе , 1826 г.)
| |
| |
| Синонимы [ 2 ] | |
|
Молге пиррогастер ( Бойе , 1826) | |

Японский огнебрюхий тритон или японская огнебрюхая саламандра ( Cynopspyrhogaster ) — вид тритона , эндемичный для Японии. Кожа на верхней части тела темная, а нижняя часть тела ярко-красная, хотя окраска варьируется в зависимости от возраста, генетики и региона. Взрослые особи имеют длину от 8 до 15 см (от 3,1 до 5,9 дюйма). Чтобы отпугивать хищников, японские огнебрюхие тритоны содержат высокий уровень тетродотоксина , нейротоксина, накапливающегося в основном с их пищей.
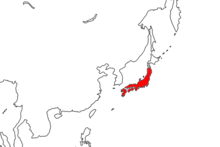
Вид встречается на многих японских островах , включая Хонсю , Сикоку и Кюсю . Их среда обитания включает как естественные, так и искусственные водоемы, а также леса и луга. Они размножаются с весны до начала лета, причем оба пола производят феромоны , когда готовы к спариванию. Яйца откладываются отдельно, вылупляются примерно через три недели. Они вырастают из личиночной формы в ювенильную за пять-шесть месяцев. Молодые особи питаются почвенной добычей, а взрослые особи поедают самых разных насекомых, головастиков и яйца своих собственных видов. У них есть несколько приспособлений , позволяющих избегать хищников, хотя то, какие они используют, зависит от того, где они живут. несколько аспектов их биологии Были изучены , в том числе их способность восстанавливать недостающие части тела .
Японский огнебрюхий тритон впервые отделился от своего ближайшего родственника в среднем миоцене , а затем разделился на четыре отдельные разновидности, каждая из которых имеет в основном отдельный ареал, хотя формально все четыре признаны составляющими один вид. В настоящее время их популяция сокращается, и они сталкиваются с угрозой болезней и торговли домашними животными . Их можно успешно содержать в неволе.
Этимология и таксономия
[ редактировать ]Вид был впервые научно описан немецким зоологом Генрихом Бойе в 1826 году как Molge Pyrrogaster . [ примечание 1 ] на основе образцов, привезенных из Японии в Европу. Он сравнил его с гладким тритоном , заявив, что принял бы первого за второго, если бы не знал, что он из Японии. Ни один из изученных им экземпляров не был полностью зрелым. [ 3 ] [ 4 ] Слово «пиррогастер» происходит от греческих слов purrhos ( букв . « огонь » ) и gastēr ( букв. « живот » ). [ 5 ] Salamandra subcristata была описана Коэнраадом Якобом Темминком и Германом Шлегелем в 1838 году и позже в том же году перенесена в Cynops швейцарским натуралистом Иоганном Якобом фон Чуди . [ 6 ] [ 2 ] а в 1850 году Cynops subcristata и Molgepyrrogaster были синонимизированы как Cynopspyrrogaster британским зоологом Джоном Эдвардом Греем . [ 7 ] [ 2 ] Исследование митохондриальной ДНК в 2001 году показало, что предполагаемые собратья Cynops , C. cyanurus и C. wolterstorffi , могут принадлежать к другому роду. [ 8 ]
Интегрированная таксономическая информационная система перечисляет шестнадцать синонимов Cynopspyrrogaster . [ 9 ] Общие названия вида: японский огнебрюхий тритон, [ 1 ] краснобрюхий тритон, [ 10 ] и японская огнепузая саламандра. [ 11 ] Исследования, изучающие морфологические и географические различия, ранее выявили шесть рас : Тохоку, Канто, Ацуми, промежуточную, Сасаяму и Хиросиму. [ 12 ] как подвид в 1969 году Один из них, сасаяма, был описан Робертом Мертенсом как Trituruspyrrogaster sasayamae , который теперь считается синонимом C.pyrrogaster . [ 2 ] Вместо этого современный молекулярный анализ подтверждает разделение C.pyrrogaster на четыре клады . [ 12 ] В частности, достоверность сасаяма и промежуточных рас никогда не была доказана, при этом одно исследование не обнаружило никаких поведенческих различий между двумя предполагаемыми формами. [ 13 ]
|
Cynopspyrhogaster отделился от своего близкого родственника C. ensicauda около 13,75 миллионов лет назад ( Mya ; во время среднего миоцена ). Общий предок этих двух видов жил на территории Евразийского материка , которая сегодня является Восточно-Китайским морем и центральными островами Рюкю . Земля, которая впоследствии стала Японскими островами, соединенными в то время с материком, вероятно, имела субтропический климат , что, возможно, заставило предков японских огнебрюхих тритонов мигрировать на север в поисках желаемой среды обитания. Со временем C.pyrrogaster разделился на четыре клады: северную, южную, западную и центральную. Первыми разошлись северные, около 9,68 млн лет назад, затем центральные около 8,23 млн лет назад и, наконец, южные и западные около 4,05 млн лет назад. Ареалы всех клад, кроме южной, сократились во время последнего ледникового периода , но впоследствии снова расширились. Исследование, в ходе которого они были идентифицированы, пришло к выводу, что эти четыре клады представляют собой отдельные таксономические единицы, хотя их точные взаимоотношения неясны. Он также отметил их крайние генетические различия, необычно большие для любого вида. [ 12 ] Ареалы центральной и западной разновидностей встречаются в Тюгоку на западе Японии, образуя гибридную зону (район, где две клады скрещиваются для получения гибридов). Центральный тип начал продвигаться на запад, что привело к смещению гибридной зоны. Ожидается, что в конечном итоге это приведет к разбавлению генома западной формы за счет увеличения гибридизации . [ 14 ]
Описание
[ редактировать ]
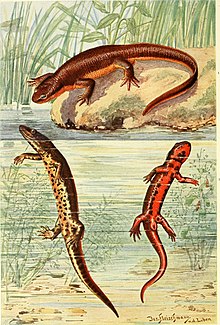
На верхней части тела тритона кожа темно-коричневая, приближающаяся к черной, и покрыта бородавчатыми бугорками. Низ живота и нижняя часть хвоста ярко-красные с черными пятнами. [ 4 ] Молодые особи имеют кремовую окраску вместо красной, хотя у большинства более крупных молодых особей присутствует немного красного цвета. [ 15 ] У взрослых особей с небольших островов, как правило, больше красного цвета на вентральной (брюшной) области, чем у особей с более крупных островов, иногда с очень маленькими пятнами или вообще без них. В целом у самцов больше красного цвета, чем у самок. [ 16 ] Самцов также можно отличить от самок по плоскому, широкому хвосту и вздутию в брюшной области. [ 17 ] Существует полностью красный вариант: считается, что эта окраска передается по наследству и рецессивна . Этот вариант не ограничивается какой-либо одной популяцией, а более распространен в западной половине Японии в целом. [ 18 ]
Сошниково -небные зубы, группа зубов в верхней задней части рта, расположены в два ряда. Язык сравнительно небольшой, составляет половину ширины рта. Ноздри расположены спереди (к голове), ближе друг к другу , чем к глазам, и при взгляде сверху почти не заметны. Пальцы ног самцов длиннее, чем у самок, хотя сами самки длиннее. Хвост плотно сжат, с плавниками как сверху, так и снизу. Гладкий гребень проходит от затылка до хвоста. [ 19 ] Полная длина тела взрослых особей составляет от 8 до 15 см (от 3,1 до 5,9 дюйма). [ 11 ] Длина морды и вентиляционных отверстий может составлять от 43,0 до 64,0 мм (1,69 и 2,52 дюйма) для самцов и от 48,5 до 75,0 мм (1,91 и 2,95 дюйма) для самок. Население более северных и возвышенных регионов, как правило, больше, чем население южных и более низких регионов. [ 20 ] Яйца имеют длину от 2,1 до 2,3 мм (от 0,083 до 0,091 дюйма). [ 17 ]
Распространение и среда обитания
[ редактировать ]Cynops пиррогастер является эндемиком Японии и встречается на нескольких островах архипелага , включая Хонсю , Сикоку и Кюсю . [ 1 ] В основном он обитает на крупных островах, тогда как его родственник C. ensicauda встречается на островах Рюкю. У него самый северный ареал среди всех Cynops видов ; все остальные виды, кроме вышеупомянутого C. ensicauda , произрастают в южном Китае. [ 12 ] также обитает интродуцированная популяция На Хатидзё-дзиме , которая, как полагают, произошла от жителей Сикоку. Считается, что их появление произошло в 1970-х годах, хотя точно, как это произошло, неизвестно. [ 21 ] Он был записан в США трижды, во Флориде и Массачусетсе. Каждый случай был либо побегом, либо преднамеренным выпуском, и ни одна популяция не была установлена. [ 11 ]
Из четырех клад северная встречается в округах Тохоку и Канто . Это не пересекается с ареалом центральной клады, которая встречается в Тюбу , северном Кансае и восточном Тюгоку. Ареал центрального региона немного перекрывается с ареалом западного, который встречается на юге Кинки, западном Тюгоку, Сикоку и центральном Кюсю. Западная клада также частично пересекается с южной кладой, которая встречается на западе и юге Кюсю. [ 12 ]
Тритоны встречаются на высоте от 30 до 2020 м (от 98 до 6627 футов). Экосистемы, в которых они обитают, включают леса, луга, кустарники, водно-болотные угодья, озера, болота и возделываемые территории. Они также могут обитать в искусственных водоемах, таких как для аквакультуры . пруды [ 1 ]
Поведение и экология
[ редактировать ]
Размножение и жизненный цикл
[ редактировать ]Размножение происходит на рисовых полях, прудах, ручьях, заводях и ручьях. Самки принимают ухаживание самцов с весны до начала лета. [ 20 ] И самцы, и самки производят пептидные феромоны , чтобы привлечь противоположный пол, когда они готовы к спариванию. Самцы производят тип, известный как содефрин (от японского термина содефури , букв. « вымогательство » ); [ 22 ] у самок есть своя разновидность, названная иморином его первооткрывателями (от японского термина imo , букв. « любимая женщина » , и rin от содефрина ). Они высвобождаются из клоаки и являются первыми пептидными феромонами, идентифицированными у позвоночных , и первыми, идентифицированными у самок позвоночных соответственно. [ 10 ] [ 23 ]
Ухаживание начинается, когда самец приближается к самке, обнюхивая ее бока или клоаку. Затем самец подносит свой хвост к самке и быстро вибрирует им. В ответ самка толкает самца мордой в шею. В этот момент самец медленно удаляется, взмахивая хвостом, а самка следует за ним, касаясь хвоста мордой, когда оказывается достаточно близко. Затем самец откладывает от двух до четырех сперматофоров по одному, перемещаясь на несколько сантиметров после каждого, которые самка пытается подобрать своей клоакой, иногда безуспешно. [ 24 ] Самки откладывают яйца отдельно на подводные объекты, такие как листья и погруженные корни травы, оплодотворяемые один за другим из несущих их сперматофоров. За один сеанс они могут отложить до 40 яиц, а за сезон размножения — от 100 до 400 яиц. [ 24 ]

Молодь вылупляется из яиц примерно через три недели в виде плавающих жаберных личинок со спинными хвостовыми плавниками. За первые три месяца жизни они вырастают примерно на 3 см (1,2 дюйма). Через пять-шесть месяцев они перестают есть и претерпевают метаморфозы , теряют жабры и плавники и становятся молодыми. Молодые особи не могут оставаться погруженными в воду, как личинки, иначе они тонут. [ 25 ] [ 26 ] Тритоны на более низких высотах созревают быстрее, чем на более высоких. Самцы тритонов из высокогорных популяций, как правило, живут дольше после достижения зрелости, но их взрослый размер не такой большой, как у низинных тритонов. Были обнаружены дикие особи возрастом двадцать три года. [ 20 ]
Сперматогенез
[ редактировать ]Cynopspyrrogaster считается идеальной моделью позвоночных для изучения механизмов, участвующих в переходе от митоза к мейозу во время сперматогенеза . [ 27 ] У мужчин этот переход включает экспрессию PCNA , вспомогательного белка ДНК-полимеразы дельта, используемого в репликации и репарации ДНК . [ 27 ] В переходе также участвует DMC1 , белок, используемый в генетической рекомбинации . [ 27 ]
Диета
[ редактировать ]в неволе головастики Известно, что охотно поедают личинок комаров , артемий и дождевых червей . [ 25 ] Молодые особи часто поедают почвенные виды Collembola (ногохвостки) и Acari (клещи). [ 15 ] Было обнаружено, что взрослые особи одного субальпийского болота в горах Адзума в префектуре Фукусима любят как живую добычу, так и падаль . Они потребляют многие разновидности насекомых, например, представителей Odonata , к которым относятся стрекозы и стрекозы, личинки которых были обнаружены в желудках тритонов целиком, но только на кусках взрослых особей; Brachycera , подотряд Diptera (мухи); Hymenoptera , к которым относятся пилильщики, осы, пчелы и муравьи; и Coleoptera (жуки). Они также поедают головастиков Rhacophorus arboreus и яйца себе подобных. Состав их рациона меняется в зависимости от сезона и года, что позволяет предположить изменения в составе мелких животных внутри и вокруг прудов, в которых они обитают. [ 17 ] Аналогичные результаты были получены в пруду на территории кампуса Токийского столичного университета в Хатиодзи , Токио: желудки тритонов содержали насекомых самых разных отрядов , а также яйца особей сородичей. Как и раньше, в пищу употреблялись головастики лягушек, хотя они принадлежали к виду Rhacophorus schlegelii . [ 28 ]
Хищники
[ редактировать ]
Тритоны на материковой части Японии ведут себя иначе, чем тритоны на небольших островах. Особи на небольших островах (например, на острове Фукуэ ) обычно используют маневр, называемый рефлексом ункен , при котором они подставляют нападающим свое ярко-красное подбрюшье . Поскольку их основными хищниками являются птицы, способные различать красный цвет, этот метод эффективен. В материковой Японии тритонам также следует избегать хищников-млекопитающих, которые не различают цвета так же хорошо, как птицы -охотники. Это приводит к тому, что эти группы населения меньше используют этот маневр, поскольку попытка его применения может привести к смерти. [ 16 ]
Против змей тритоны с острова Фукуэ, как правило, демонстрируют виляние хвостом, призванное привлечь внимание хищника к их сменному хвосту, а не к более важной голове; выходцы из префектуры Нагасаки на материковой части Японии обычно просто бегут. Змеи присутствуют в обеих областях. Такая разница в поведении, вероятно, связана с тем, что тритоны с материка приспособлены к бегству от охотников на млекопитающих, которых такая демонстрация с меньшей вероятностью отпугнет. [ 29 ]
Токсин
[ редактировать ]Дикие японские огнебрюхие тритоны содержат высокий уровень нейротоксина тетродотоксина ( ТТХ). [ 30 ] Этот токсин ингибирует активность натриевых каналов у большинства позвоночных, препятствуя хищничеству как птиц, так и млекопитающих. [ 29 ] Эксперименты показали, что токсин почти полностью получен из рациона тритона. Выращенные в неволе без источника ТТХ, молодые особи в возрасте от 36 до 70 недель не содержали обнаруживаемых уровней, но дикие экземпляры из той же исходной среды обитания имели высокую токсичность. У молодых тритонов, выращенных в неволе, все еще обнаруживалось некоторое количество ТТХ, которое, как предполагалось, было перенесено взрослыми самками в их яйца. [ 30 ] В последующем эксперименте, проведенном той же командой, выращенным в неволе тритонам давали пищу, содержащую нейротоксин. Они охотно потребляли мотыля с добавлением ТТХ, когда им предлагали, не проявляя никаких симптомов после проглатывания яда. Впоследствии его можно было обнаружить в их телах, что еще раз указывает на то, что источником токсина является пища. В их среде обитания не известны организмы, продуцирующие ТТХ, но их существование вполне вероятно, и это могло бы объяснить происхождение ТТХ у диких тритонов. [ 31 ]
Сохранение
[ редактировать ]
Международный союз охраны природы (МСОП) причислил его к находящемуся под угрозой исчезновения . Такая оценка была сделана в 2020 году. [ 1 ] сдвиг по сравнению с 2004 годом, когда он был оценен как вызывающий наименьшее беспокойство . [ 32 ] Он успешно размножается в австралийских зоопарках. [ 1 ] Одной из основных угроз, с которыми сталкивается C.pyrrogaster, является сбор для торговли домашними животными . МСОП заявляет, что эту торговлю необходимо немедленно прекратить. Их популяция сокращается, особенно вблизи районов проживания человека. [ 1 ]
Японские огнебрюхие тритоны с загадочными поражениями кожи на озере Бива в японской префектуре Сига оказались заражены инфекциями, вызванными одноклеточными эукариотами отряда Dermocystida . Поражения содержали кисты , наполненные спорами . Почти все поражения были внешними, хотя одно было обнаружено на печени . Во всем мире болезни являются одной из причин сокращения популяций амфибий. Есть опасения, что это заболевание может распространиться на другие близлежащие виды, включая Zhangixalus arboreus и Hynobius vandenburghi . [ 33 ]
Считалось, что этот сорт, встречавшийся исключительно на полуострове Ацуми , вымер в 1960-х годах. Затем, в 2016 году, трио исследователей обнаружили, что тритоны на Читинском полуострове, скорее всего, были одним и тем же вариантом из-за схожих морфологических особенностей. Обе группы предпочитают более прохладную температуру и имеют гладкие и мягкие тела, бледную спинную часть и желтоватую нижнюю сторону. Даже если эта форма еще жива, она находится под серьезной угрозой и вскоре будет уничтожена без немедленной защиты. [ 34 ]
Взаимодействие с людьми
[ редактировать ]Исследовать
[ редактировать ]Японские огнебрюхие тритоны служат очень полезным модельным организмом в лабораторных условиях, но после метаморфоза за ними становится труднее ухаживать. Эксперимент, проведенный при поддержке Японского общества содействия науке, показал, что тиомочевина (TU) может предотвратить этот процесс, позволяя животным оставаться в своей преметаморфозной форме в течение двух лет, сохраняя при этом способность метаморфизироваться после удаления. из решения ТУ. Это не оказало никакого влияния на их регенерационные способности. [ 25 ]
Японские огнебрюхие тритоны вырабатывают мотилин — пептид, стимулирующий желудочно-кишечные сокращения , обнаруженный у многих позвоночных. Он создается в верхних отделах тонкой кишки и поджелудочной железе . Открытие последнего было первым случаем наблюдения мотилина поджелудочной железы. Орган также вырабатывает инсулин . Эти результаты представляют собой первое открытие мотилина у земноводных, что позволяет предположить, что он играет для них такую же роль, как и для птиц и млекопитающих. Существование панкреатического мотилина также указывает на другую, неизвестную функцию. [ 35 ]
Этот вид, как и другие амфибии Urodele , способен отращивать недостающие части тела , включая конечности с функциональными суставами и нижнюю челюсть . [ 36 ] [ 37 ] Когда происходит этот процесс, регенерированная ткань имеет тенденцию повторять форму неповрежденной ткани. [ 36 ] Он также способен вырастить недостающие линзы : на это уходит тридцать дней в личинке и восемьдесят дней во взрослом состоянии. Разница во времени обусловлена исключительно размером глаза, а регенеративная способность не меняется; открытие этого факта противоречило популярному утверждению, что молодые животные регенерируют быстрее, чем взрослые. [ 38 ]
В плену
[ редактировать ]Цинопс пиррогастер можно содержать в неволе. Доктор ветеринарной медицины Лианна МакЛеод описала их как «не требующих особого ухода», отметив, что тритоны в неволе любят мотыля, артемию, стеклянную креветку , дафнию , а для более крупных особей — гуппи . [ 39 ]
Примечания
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г Группа специалистов МСОП по амфибиям SSC (2021 г.). « Цинопс пиррогастер » . Красный список исчезающих видов МСОП . 2021 : e.T59444A177224976. doi : 10.2305/IUCN.UK.2021-1.RLTS.T59444A177224976.en . Проверено 12 ноября 2021 г.
- ^ Jump up to: а б с д и Фрост, Даррел Р. (2023). « Цинопс пиррогастер (Бойе, 1826)» . Виды амфибий мира: онлайн-справочник . дои : 10.5531/db.vz.0001 . Архивировано из оригинала 2 мая 2021 года . Проверено 8 января 2023 г.
- ^ Окен, Лоренц (1826). «Исида фон Окен» . Экспедиция Исиды (на немецком языке): 203–204. Архивировано из оригинала 10 января 2023 года . Проверено 8 января 2023 г.
- ^ Jump up to: а б Бойе, Генрих (1827). Характеристики некоторых японских земноводных (на голландском языке). Джо. Ф. Снеллеман. стр. 30–31. дои : 10.5962/bhl.title.46817 . OCLC 727216017 . Архивировано из оригинала 28 октября 2022 года . Проверено 28 октября 2022 г.
- ^ Джоблинг, Джеймс А. (2010). Словарь научных названий птиц Хелма [электронный ресурс]: от аалге до зусии . Кристофер Хелм. п. 326. ИСБН 978-1-4081-3326-2 . Проверено 13 ноября 2022 г.
- ^ Чуди, Иоганн Якоб фон; Агассис, Луи (1838). Классификация батрахов: с учетом ископаемых животных этого отдела рептилий / Дж. Я. Чуди (на немецком языке). Университет Невшателя . п. 94. дои : 10.5962/bhl.title.4883 . OCLC 964903266 . Архивировано из оригинала 28 октября 2022 года . Проверено 28 октября 2022 г.
- ^ Грей, Джон Эдвард (1850). Каталог образцов амфибий в коллекции Британского музея. Часть II. Батрахия градиентиа и др . Отпечатано по заказу попечителей. п. 25. дои : 10.5962/bhl.title.64048 . OCLC 3183646 . Архивировано из оригинала 28 октября 2022 года . Проверено 28 октября 2022 г.
- ^ Чан, Лорен М.; Самудио, Келли Р.; Уэйк, Дэвид Б. (декабрь 2001 г.). «Взаимоотношения родов саламандридов Paramesotriton , Pachytriton и Cynops на основе последовательностей митохондриальной ДНК» . Копейя . 2001 (4): 997–1009. doi : 10.1643/0045-8511(2001)001[0997:ROTSGP]2.0.CO;2 . ISSN 0045-8511 . S2CID 46994906 . Архивировано из оригинала 28 октября 2022 года . Проверено 28 октября 2022 г.
- ^ « Цинопс пиррогастер » . itis.gov . Архивировано из оригинала 30 октября 2022 года . Проверено 30 октября 2022 г.
- ^ Jump up to: а б Накада, Томоаки; Тойода, Фумиё; Мацуда, Кохэй; Накакура, Такаши; Хасунума, Итару; Ямамото, Кадзутоши; Оноуэ, Сатоми; Ёкосука, Макото; Кикуяма, Сакаэ (25 января 2017 г.). «Иморин: феромон сексуальной привлекательности самок краснобрюхих тритонов ( Cynopspyrrogaster )» . Научные отчеты 7 (1): 41334. Бибкод : 2017НацСР... 741334Н дои : 10.1038/srep41334 . ISSN 2045-2322 . ПМК 5264602 . ПМИД 28120945 .
- ^ Jump up to: а б с « Цинопс пиррогастер » . usgs.gov . Архивировано из оригинала 28 октября 2022 года . Проверено 27 октября 2022 г.
- ^ Jump up to: а б с д и ж Томинага, Ацуши; Мацуи, Масафуми; Ёсикава, Нацухико; Нисикава, Канто; Хаяси, Терутаке; Мисава, Ясучика; Танабэ, Синго; Ота, Хидетоши (март 2013 г.). «Филогения и историческая демография Cynopspyrrogaster (Amphibia: Urodela): Таксономические отношения и изменения в распространении, связанные с климатическими колебаниями» . Молекулярная филогенетика и эволюция . 66 (3): 654–667. дои : 10.1016/j.ympev.2012.10.015 . ПМИД 23103571 . Проверено 28 октября 2022 г.
- ^ Тагами, Масатака; Хори, Чикако; Каваи, Тосимаса; Сакабе, Ай; Симада, Томохико (2015). «Брачное поведение Cynopspyrrogaster из префектур Гифу и Айти, Центральная Япония, в неволе» . Современная герпетология . 34 (1): 12–18. дои : 10.5358/hsj.34.12 . S2CID 86006031 . Архивировано из оригинала 30 октября 2022 года . Проверено 30 октября 2022 г.
- ^ Томинага, Ацуши; Мацуи, Масафуми; Ёсикава, Нацухико; Это, Коширо; Нисикава, Канто (16 марта 2018 г.). «Геномное смещение и сдвиг гибридной зоны у японского огнебрюхого тритона» . Журнал наследственности 109 (3): 232–242. doi : 10.1093/jhered/esx085 . ПМИД 29566204 . Архивировано 29 октября. из оригинала Получено 29 октября.
- ^ Jump up to: а б Мацуи, Куми; Мочида, Кодзи; Накамура, Масахиса (июль 2003 г.). «Пищевые привычки молоди японского тритона Cynopspyrrogaster » . Зоологическая наука . 20 (7): 855–859. дои : 10.2108/zsj.20.855 . ПМИД 12867714 . S2CID 40530531 .
- ^ Jump up to: а б Мочида, Кодзи (23 июня 2009 г.). «Параллельная географическая мозаика морфологических и поведенческих апосематических признаков тритона Cynopspyrrogaster (Urodela: Salamandridae): параллельная вариация апосематических признаков» . Биологический журнал Линнеевского общества . 97 (3): 613–622. дои : 10.1111/j.1095-8312.2008.01182.x .
- ^ Jump up to: а б с Ихара, Садао (февраль 2014 г.). «Пищевые привычки взрослого японского тритона» . Современная герпетология . 33 (1): 38–45. дои : 10.5358/hsj.33.38 . ISSN 1345-5834 . S2CID 83534797 . Архивировано из оригинала 30 октября 2022 года . Проверено 30 октября 2022 г.
- ^ Мацуи, Куми; Маруноути, Джунсукэ; Накамура, Масахиса (2003). «Красные варианты японского тритона Cynopspyrrogaster (Amphibia: Salamandridae): обзор записей и наблюдения в неволе о наследственности окраски» . Современная герпетология . 22 (1): 37–42. дои : 10.5358/hsj.22.37 . S2CID 89239218 . Архивировано из оригинала 30 октября 2018 года . Проверено 30 октября 2022 г.
- ^ Стейнегер, Леонхард (1907). «Герпетология Японии и сопредельной территории» . Бюллетень Национального музея США (58): 16. doi : 10.5479/si.03629236.58.i . Архивировано из оригинала 30 октября 2022 года . Проверено 30 октября 2022 г.
- ^ Jump up to: а б с Очи, Осаму; Маруноути, Джунсукэ; Уэда, Хироаки (1 января 2000 г.). «Изменения в возрасте и размере гнездящихся популяций японских тритонов Cynopspyrhogaster на разных высотах » . Земноводные-рептилии . 21 (3): 381–396. дои : 10.1163/156853800507444 . ISSN 1568-5381 . Архивировано из оригинала 29 октября 2022 года . Проверено 29 октября 2022 г.
- ^ Томинага, Ацуши; Мейер-Рохов, В. Бенно; Окамото, Таку; Курияма, Такео; Нисикава, Канто; Мацуи, Масафуми (февраль 2016 г.). «Происхождение и генетическое единообразие интродуцированной популяции Cynopspyrrogaster (амфибия: Urodela) на острове Хатидзёдзима» . Современная герпетология . 35 (1): 64–68. дои : 10.5358/hsj.35.64 . S2CID 87404627 . Архивировано 30 октября. из оригинала Получено 30 октября.
- ^ Ван Бокслаер, Инес; Маекс, Марго; Трир, Даг; Янссенсвиллен, Сунита; Янссенс, Рик; Вандеберг, Вим; Пруст, Пол; Боссайт, Фрэнки (3 марта 2016 г.). «Помимо содефрина: доказательства существования многокомпонентной феромонной системы у модельного тритона Cynopspyrhogaster (Salamandridae)» . Научные отчеты . 6 (1): 21880. Бибкод : 2016NatSR...621880V . дои : 10.1038/srep21880 . ISSN 2045-2322 . ПМК 4776240 . ПМИД 26935790 .
- ^ Кикуяма, С.; Тойода, Ф.; Омия, Ю.; Мацуда, К.; Танака, С.; Хаяши, Х. (1995). «Содефрин: пептидный феромон, привлекающий самок, в клоакальных железах тритона» . Наука . 267 (5204): 1643–1645. Бибкод : 1995Sci...267.1643K . дои : 10.1126/science.7886452 . ISSN 0036-8075 . JSTOR 2886746 . ПМИД 7886452 . S2CID 38424857 . Архивировано из оригинала 29 октября 2022 года . Проверено 29 октября 2022 г.
- ^ Jump up to: а б Куцуки, Такако; Хасэгава, Эйсуке (сентябрь 2016 г.). «Женские предпочтения как в поведении, так и в морфологических особенностях самца японского тритона Cynopspyrhogaster » . Журнал этологии . 34 (3): 337–342. дои : 10.1007/s10164-016-0480-x . S2CID 11330345 . Архивировано из оригинала 30 октября 2022 года . Проверено 30 октября 2022 г.
- ^ Jump up to: а б с Чиба, Чикаго; Ямада, Шута; Танака, Хибики; Инаэ-Чиба, Майко; Миура, Томоя; Халл-Оукс, Мартин Майкл; Ёсикава, Таро; Инами, Ватару; Мизуно, Аки; Ислам, доктор медицинских наук. Рафикуль; Хан, Вендже; Ясумуро, Хирофуми; Мацумото, Микико; Такаянаги, Мияко (1 мая 2012 г.). «Подавление метаморфоза: альтернативный протокол выращивания тритона Cynopspyrrogaster » . Зоологическая наука . 29 (5): 293–298. дои : 10.2108/zsj.29.293 . ПМИД 22559962 . S2CID 22637590 . Получено 30 октября.
- ^ Халл-Оукс, Мартин Майкл; Ямада, Шута; Миура, Томоя; Накамура, Кента; Хейнс, Трейси; Маки, Нобуясу; Дель Рио-Цонис, Катя; Цонис, Riot A; Чиба, Чикаго (май 2011 г.). «Экспрессия экзогенных генов у тритонов путем трансгенеза» . Протоколы природы . 6 (5): 600–608. дои : 10.1038/nprot.2011.334 . ПМИД 21527918 . S2CID 5305751 . Архивировано 18 ноября. из оригинала Получено 30 октября.
- ^ Jump up to: а б с Ядзава Т., Ямамото Т., Накаяма Ю., Хамада С., Абе С. Преобразование из митоза в мейоз: морфология и экспрессия ядерного антигена пролиферирующих клеток (PCNA) и Dmc1 во время сперматогенеза тритона. Разница в росте разработчиков. 2000 декабрь;42(6):603-11. дои: 10.1046/j.1440-169x.2000.00544.x. PMID 11142682
- ^ Хикари, Накагава; Хо, Кусано (2007). Пищевые привычки краснобрюхих тритонов (Cynopspyrrogaster) в Минамиосаве, городе Хатиодзи, Токио Хикару Накагава, Тамоцу Кусано . Бюллетень Герпетологического общества (на японском языке). 2007 (1): 1–5. дои : 10.14880/hrghsj1999.2007.1 . Архивировано из оригинала 31 октября 2022 года . Проверено 31 октября 2022 г. (требуется подписка)
- ^ Jump up to: а б Мочида, Кодзи; Мори, Акира (29 ноября 2021 г.). «Противохищное поведение тритонов ( Cynopspyrrogaster ) против змей» . ПЛОС ОДИН . 16 (11): e0258218. Бибкод : 2021PLoSO..1658218M . дои : 10.1371/journal.pone.0258218 . ПМЦ 8629279 . ПМИД 34843491 .
- ^ Jump up to: а б Кудо, Юта; Тиба, Чикафуми; Коноки, Кейичи; Чо, Юко; Ёцу-Ямасита, Мари (июль 2015 г.). «Подтверждение отсутствия тетродотоксина и его аналогов у молоди японского огнебрюхого тритона Cynopspyrrogaster , выращенного в неволе из яиц в лаборатории с помощью HILIC-LC-MS» . Токсикон . 101 : 101–105. doi : 10.1016/j.токсикон.2015.05.008 . ПМИД 25986913 . Проверено 27 октября 2022 г.
- ^ Кудо, Юта; Тиба, Чикафуми; Коноки, Кейичи; Чо, Юко; Ёцу-Ямасита, Мари (октябрь 2017 г.). «Диетическое введение тетродотоксина и его предполагаемых промежуточных продуктов биосинтеза выращенному в неволе нетоксичному японскому огнебрюхому тритону Cynopspyrrogaster » . Токсикон . 137 : 78–82. дои : 10.1016/j.токсикон.2017.07.016 . ПМИД 28734983 . Проверено 28 октября 2022 г.
- ^ «Японский огнебрюхий тритон» . iucnredlist.org . Архивировано из оригинала 26 января 2021 года . Проверено 30 октября 2022 г.
- ^ Кавахара, Го; Такаяма, Юта; Сугияма, Макото; Икадай, Хироми; Хашимото, Осаму (2022). «Дермоцистидная инфекция у японского четырехбрюхого тритона Cynopspyrrogaster » . Журнал ветеринарной медицины . 84 (10): 1410–1416. дои : 10.1292/jvms.22-0233 . ПМЦ 9586028 . ПМИД 36047163 . S2CID 251945249 . Архивировано 29 октября. из оригинала Получено 29 октября.
- ^ Симада, Томохико; Маэда, Сиота; Сакакибара, Масаки (февраль 2016 г.). «Морфологическое исследование Cynopspyrrogaster с Читинского полуострова: повторное открытие «вымершей» расы Ацуми, эндемической для полуостровных регионов префектуры Айти, Центральная Япония» . Современная герпетология . 35 (1): 38–52. дои : 10.5358/hsj.35.38 . ISSN 1345-5834 . S2CID 88355202 . Архивировано 30 октября. из оригинала Получено 30 октября.
- ^ Мацумото, Мио; Такеми, Шота; Сакаи, Такафуми; Саката, Ичиро (июль 2022 г.). «Идентификация мотилина у японского огнебрюхого тритона» . Общая и сравнительная эндокринология . 323–324: 114031. doi : 10.1016/j.ygcen.2022.114031 . ПМИД 35331740 . S2CID 247653195 .
- ^ Jump up to: а б Цуцуми, Рио; Иноуэ, Такеши; Ямада, Сигэхито; Агата, Киёкадзу (февраль 2015 г.). «Реинтеграция регенерировавших и сохранившихся тканей при регенерации суставов у тритона Cynopspyrrogaster » . Регенерация . 2 (1): 26–36. дои : 10.1002/reg2.28 . ISSN 2052-4412 . ПМЦ 4895332 . ПМИД 27499865 .
- ^ Куросака, Хироши; Такано-Ямамото, Теруко; Ямасиро, Такаси; Агата, Киёкадзу (февраль 2008 г.). «Сравнение молекулярных и клеточных событий во время регенерации нижней челюсти тритона ( Cynopspyrrogaster ) и западноафриканской шпорцевой лягушки ( Xenopustropicis )» . Динамика развития . 237 (2): 354–365. дои : 10.1002/dvdy.21419 . ПМИД 18161063 . S2CID 41117859 .
- ^ Иноуэ, Такеши; Иноуэ, Ре; Бег, Рио; Тада, Кикуо; Урата, Юко; Мичибаяси, Чиаки; Такемура, Шота; Агата, Киёкадзу (октябрь 2012 г.). «Хрусталик регенерирует посредством аналогичных процессов и сроков у взрослых особей и личинок тритона Cynopspyrhogaster » . Динамика развития . 241 (10): 1575–1583. дои : 10.1002/dvdy.23854 . ПМИД 22930574 . S2CID 6513165 . Архивировано 30 октября. из оригинала Получено 30 октября.
- ^ Маклеод, Лианна. «Огнебрюхий тритон: профиль вида» . thesprucepets.com . Архивировано из оригинала 30 октября 2022 года . Проверено 30 октября 2022 г.