Саламандре
| Саламандры | |
|---|---|

| |
| Пятнистый саламандр , Амбистома Макулатум | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Сорт: | Амфибия |
| Клада : | Хвостат |
| Заказ: | Уродела Duméril , 1806 |
| Подчиненные | |
| |
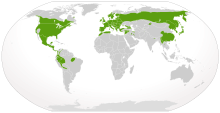
| Нативное распределение саламандров (в зеленом) | |
Саламандры -это группа амфибий , обычно характеризующихся их ящеристого внешнего вида, с тонкими телами, тупыми мордами, короткими конечностями, выступающими под прямым углом на тело, и наличием хвоста как у личинок, так и у взрослых. Все десять существующих семей Саламандре сгруппированы вместе под приказом Urodela из группы Caudata . [ 2 ] Уродела - это научный латинский термин, основанный на древнем греческом языке ὐ Δήλη : ourà dēlē «заметный хвост». Caudata - это латынь для «хвостовых», от Cauda : «Хвост».
Разнообразие саламандры является самым высоким в восточной части Северной Америки , особенно в Аппалачских горах ; Большинство видов встречаются в царстве Холрктики , с некоторыми видами, присутствующими в неотропической сфере . Саламандры никогда не имеют более четырех пальцев ног на передних ногах и пять на их задней ногах, [ 3 ] Но у некоторых видов меньше цифр, а другим не хватает задней конечности. Их проницаемая кожа обычно зависит от их среды обитания в воде или рядом с водой или в других прохладных, влажных местах. Некоторые виды саламандры полностью водные на протяжении всей своей жизни, некоторые с перерывами в воду, а другие - совершенно наземные, как взрослые.
Эта группа амфибий способна регенерировать потерянные конечности, а также другие поврежденные части их тела. Исследователи надеются обратить вспять регенеративные процессы для потенциальных медицинских применений человека, таких как лечение травмы головного и спинного мозга или предотвращение вредного рубца во время восстановления операции на сердце. [ 4 ] Замечательная способность саламандров регенерация ограничивается не только конечностями, но и распространяется на жизненно важные органы, такие как сердце, челюсть и части спинного мозга, показывая их уникальность по сравнению с различными типами позвоночных. Эта способность наиболее замечательна для возникновения без каких -либо рубцов. «Это сделало саламандры бесценным модельным организмом в научных исследованиях, направленных на понимание и достижение регенеративных процессов для медицинских достижений в биологии человека и животных. [ 5 ]
Члены семейства Salamandridae в основном известны как Newts , и не хватает канавки вдоль по бокам их тела, типичных для других групп. Кожа некоторых видов содержит мощный яд тетродотоксин ; Эти саламандры, как правило, медленно движутся и имеют яркую окраску, чтобы рекламировать их токсичность. Саламандры обычно откладывают яйца в воде и имеют водные личинки, но в их жизненном цикле возникают большие различия . Некоторые виды в суровых условиях воспроизводятся, пока все еще находятся в состоянии личинок.
Этимология
[ редактировать ]Слово саламандр происходит из старого французского саламандра из Латинской саламандры от греческого σαλαμάνΔρα : саламандра, неопределенного, возможно, до грика. [ 6 ] Греческое слово используется для пожарной саламандры . [ 7 ]
Описание
[ редактировать ]
Кожа не хватает весов, влажной и гладкой на ощупь, за исключением новых саламандрид, которые могут иметь бархатистую или бородавленную кожу, мокрой на ощупь. Кожа может быть милой или ярко окрашенной, демонстрируя различные узоры полос, стержней, пятен, пятен или точек. Мужские новые становятся драматично окрашенными в течение сезона размножения. Пещеры, жилые в темноте, не имеют пигментации и имеют полупрозрачный розовый или перламусный вид. [ 8 ]
Саламандры варьируются в размере от минутных саламандров , с общей длиной 27 мм ( 1 + 1 ~ 8 дюймов), включая хвост, к китайскому гигантскому саламандре , которая достигает 1,8 м (6 футов) и весит до 65 кг (145 фунтов). Все крупнейшие виды встречаются в четырех семействах гигантских саламандров , сирен , конго и протеиды , которые являются водными и облигационными педоморфами. [ 9 ] [ 10 ] Некоторые из крупнейших наземных саламандров, которые проходят полную метаморфозу, принадлежат к семейству тихоокеанских гигантов саламандров и намного меньше. [ 11 ] Большинство саламандров, однако, имеют длину от 10 до 20 см (4 и 8 дюймов). [ 12 ]
Багажник, конечности и хвост
[ редактировать ]Взрослый саламандр обычно напоминает небольшую ящерицу, имеющую базальную тетраподную форму с цилиндрическим стволом, четырьмя конечностями и длинным хвостом. За исключением семейства Salamandridae, голова, тело и хвост имеют ряд вертикальных депрессий на поверхности, которые проходят от средней области до вентральной области и известны как канавки . Их функция, по -видимому, заключается в том, чтобы помочь сохранить кожу влажной, направляя воду над поверхностью тела. [ 13 ]

Некоторые водные виды, такие как сирены и амфиумы , имеют уменьшенные или отсутствующие задние конечности, придающие им вид, похожий на угря , но у большинства видов передние и задние конечности имеют примерно такую же длину и проект, едва поднимая ствол с земля. Ноги широкие с короткими цифрами, обычно четыре на передних ногах и пять на задней части. Саламандры не имеют когтей, а форма ноги варьируется в зависимости от среды обитания животного. Вопросы для лазания имеют удлиненные носки с квадратным наконечником, в то время как жители каменных жителей имеют большие ноги с короткими тупыми пальцами. ( У саламандры, заполняющей дерево Bolitoglossa sp .), Есть тарелки, похожие на перепонки, которые прилипают к гладким поверхностям путем всасывания, в то время как виды гидромантов с камнями из Калифорнии имеют ноги с мясистыми сетью и короткими цифрами и используют их хвосты в качестве дополнительной конечности. При подъеме, хвост поднимает заднюю часть тела, в то время как одна задняя нога движется вперед, а затем поворачивается на другую сторону, чтобы обеспечить поддержку по мере продвижения другой задней ноги. [ 14 ]
У личинок и водных саламандров хвост сплющивается в боковом направлении, имеет дорсальные и вентральные плавники, и в восторге от стороны в сторону протолкнуть животное через воду. В семьях Ambystomatidae и Salamandridae хвост мужчины, который больше, чем у женщин, используется во время объятий Amplexus , чтобы подтолкнуть пару со спариванием в уединенное место. У наземных видов хвост движется, чтобы уравновешивать животное по мере его работы, в то время как в древесной саламандре и других видах, использующих деревьев, он является претензий . Хвост также используется определенными саламандрами Plethodontid , которые могут прыгать, чтобы помочь выйти в воздух. [ 14 ] Хвост используется в ухаживании и в качестве органа для хранения для белков и липидов. Он также функционирует как защита от хищничества, когда он может быть наброшен на злоумышленника или аутотомизирован при захвате. В отличие от лягушек, взрослый саламандр может регенерировать конечности и хвост, когда они теряются. [ 14 ]
Кожа
[ редактировать ]
Кожа саламандров, как и другие амфибии, тонкая, проницаемая для воды, служит дыхательной мембраной и хорошо поставляется с железами. Он имеет высоко оправданные внешние слои, периодически обновляемые через процесс выпадения кожи, контролируемый гормонами из гипофиза и щитовидной железы. Во время линьки кожа изначально ломается вокруг рта, и животное движется вперед через разрыв, чтобы сбросить кожу. Когда передние конечности работали ясны, серия рябь тела подталкивает кожу в заднюю часть. Задние конечности извлекаются и толкают кожу дальше, прежде чем он в конечном итоге будет освобожден от трения, когда саламандре движется вперед, прижав хвост к земле. [ 15 ] Животное часто ест полученную кожу. [ 8 ]
Железы выпуска кожи в слизи , которые сохраняют влажную кожу, что является важным фактором дыхания кожи и терморегуляции. Липкий слой помогает защитить от бактериальных инфекций и плесени, уменьшает трение при плавании и делает животное скользким и трудным для хищников. Гранулярные железы, разбросанные на верхней поверхности, в частности, голова, спина и хвост, производят репеллентные или токсичные выделения. [ 15 ] Некоторые токсины саламандры особенно сильны. Шероховатый Newt ( Taricha Granulosa нейротоксина ) продуцирует тетродотоксин , наиболее токсичное непротеиновое вещество, известное. Обработка Newts не причиняет вреда, но проглатывание даже минутного фрагмента кожи смертельно. В испытаниях по кормлению, рыба, лягушки, рептилии, птицы и млекопитающие были признаны восприимчивыми. [ 16 ]
Зрелые взрослые некоторых видов саламандры имеют «брачную» железистую ткань в своих клоацах , у основания их хвостов, на головах или под подборами. Некоторые женщины выделяют химические вещества , возможно, из вентральной клоакальной железы, чтобы привлечь мужчин, но мужчины, похоже, не используют феромоны для этой цели. [ 17 ] В некоторых плетодонтах мужчины имеют заметные умственные железы на подбородке, которые прижимаются к ноздрам женщин во время ритуала ухаживания. Они могут функционировать, чтобы ускорить процесс спаривания, снижая риск его разрушения хищником или конкурирующим мужчиной. [ 18 ] Железная железа у основания хвоста в Plethodon Cinereus используется для маркировки фекальных пеллетов для провозглашения территориальной собственности. [ 17 ]
Чувства
[ редактировать ]

Запах
[ редактировать ]Занятие в Саламандре играет роль в техническом обслуживании территории, признании хищников и ритуалах ухаживания, но, вероятно, является второстепенным, чтобы увидеть во время выбора добычи и кормления. Саламандры имеют два типа сенсорных областей, которые реагируют на химию окружающей среды. Обонятельный эпителий в носовой полости поднимает воздухозабоченные и водные запахи, в то время как прилегающие органы из вомероназальных органов обнаруживают нелетуальные химические сигналы, такие как вкусы во рту. В Plethodonts сенсорный эпителий рвотероназальных органов распространяется на носоландские канавки , которые простираются от ноздрей до углах рта. Эти расширенные области, по -видимому, связаны с идентификацией предметов добычи, распознаванием конспецификов и идентификацией отдельных лиц. [ 19 ]
Зрение
[ редактировать ]Глаза большинства саламандров адаптированы в основном для видения ночью. У некоторых постоянно водных видов они уменьшены в размерах и имеют упрощенную структуру сетчатки , а у пещерных жителей, таких как Георгия Слепая саламандре , они отсутствуют или покрыты слоем кожи. У амфибийных видов глаза представляют собой компромисс и близки к воздуху и дальновидны в воде. Полностью наземные виды, такие как Fire Salamander, имеют более плотную линзу, которая может сосредоточиться на гораздо более широком диапазоне расстояний. [ 20 ] Чтобы найти свою добычу, саламандры используют трихроматическое цветовое зрение , распространяемое на ультрафиолетовый диапазон, на основе трех типов фоторецепторов , которые максимально чувствительны около 450, 500 и 570 нм. [ 21 ] Личинки и взрослые некоторых очень водных видов также имеют орган боковой линии , аналогичный рыбе, который может обнаружить изменения давления воды. [ 8 ]
Слушание
[ редактировать ]У всех саламандров не хватает полости среднего уха, барабанной перепонки и евстахийской трубки , но имеют систему Opercularis, такую как лягушки, и все еще способны обнаружить воздушный звук. [ 22 ] [ 23 ] Система opercularis состоит из двух оськсов: колумелла (эквивалентная станциям высших позвоночных ) , которые сливаются с черепом, и Operculum. Мышца Opercularis соединяет последнее с грудным поясом и поддерживается под напряжением, когда животное настороженно. [ 24 ] Система, по-видимому, способна обнаружить низкочастотные вибрации (500–600 Гц), которые могут быть подняты с земли на передних конечностях и переданы во внутреннее ухо. Это может служить для предупреждения животного подходящего хищника. [ 25 ]
Вокализация
[ редактировать ]Обычно считается, что саламандры не имеют голоса и не используют звук для общения так, как это делают лягушки; Однако в системе спаривания они общаются с помощью передачи сигналов феромона; Некоторые виды могут сделать тихой тикает. щелкнув, скрипит или звучит звуки, [ 26 ] Возможно, от открытия и закрытия клапанов в носу. У большинства саламандров не хватает голосового шнура, но в грязи (Necturus) и некоторых других видах присутствует гортань, а в тихоокеанском гигантском саламандре и некоторых других есть большая гортани и полосы, известные как вокалы Plicae. [ 27 ] Калифорнийский гигантский саламандр может производить кору или погремушку, и несколько видов могут скрипиться, сокращая мышцы в горле. Слушая саламандры может пискнуть, используя другой механизм; Он втягивает глаза в свою голову, вынуждая воздух изо рта. Энсатина Саламандр иногда издает шипящий звук, в то время как сирены иногда производят тихие клики и могут прибегнуть к слабым воплям, если его атаковывают. Подобное поведение клика наблюдалось в двух европейских Newts Lissotriton vulgaris и Ichthyosaura alpestris в их водной фазе. [ 26 ] Вокализация в Саламандре мало изучалась, и предполагается, что цель этих звуков - поражение хищников. [ 28 ]

Дыхание
[ редактировать ]Дыхание отличается среди различных видов саламандров и может включать жабры, легкие, кожу и мембраны рта и горла. Личиночные саламандры дышат в основном с помощью жабр , которые обычно являются внешними и перьями по внешнему виду. Вода втягивается через рот и вытекает через прорезь жабры. Некоторые неотенические виды, такие как Mudpuppy ( Necturus maculosus ), сохраняют свои жабры на протяжении всей своей жизни, но большинство видов теряют их при метаморфозе . Эмбрионы , которые подвергаются прямому развитию, имеют большие жабры , некоторых наземных саламандров без легких, таких как Энсатина которые находятся рядом с поверхностью яйца. [ 29 ]
При наличии у взрослых саламандров легкие сильно различаются среди разных видов по размеру и структуре. В водных видах холодных вод, таких как торрент саламандры ( Rhyacotriton ), легкие очень малы с гладкими стенами, в то время как виды, живущие в теплой воде с небольшим количеством растворенного кислорода, таких как меньшая сирена ( Siren Intermedia ), имеют большие легкие с созреваемыми поверхности. В саламандерах без легких ( семейство Plethodontidae и когтистые саламандры в семействе азиатских саламандров ), легких или жабр не присутствуют, и газовый обмен в основном происходит через кожу, известную как кожное дыхание , дополненные тканями, выстилающими рот. Чтобы облегчить это, у этих саламандров есть плотная сеть кровеносных сосудов прямо под кожей и во рту. [ 29 ] [ 30 ] [ 31 ]
В Amphiums метаморфоза является неполной, и они сохраняют одну пару жабрных прорези в качестве взрослых, с полностью функционирующими внутренними легкими. [ 32 ] Некоторые виды, в которых не хватает легких, дышат через жабры. В большинстве случаев это внешние жабры, видимые как пучки с обеих сторон головы. Некоторые наземные саламандры имеют легкие, используемые в дыхании, хотя они просты и похожи на мешковины, в отличие от более сложных органов, обнаруженных у млекопитающих . Многие виды, такие как OLM , имеют как легкие, так и жабры в качестве взрослых. [ 8 ]

В нектаре внешние жабра начинают образовываться как средство борьбы с гипоксией в яйце, когда яичный желток превращается в метаболически активную ткань. [ 33 ] Тем не менее, молекулярные изменения в Mudpuppy во время пост-эмбрионального развития в первую очередь из-за щитовидной железы предотвращают интернализацию внешних жабл, как видно в большинстве саламандров, которые подвергаются метаморфозе. [ 34 ] Внешние жабры, наблюдаемые в саламандерах, сильно отличаются от амфибий с интернализованными жабрами. В отличие от амфибий с интернализованными жабрами, которые обычно опираются на изменение давления в рамках промежуточных и глоточных полостей, чтобы обеспечить диффузию кислорода на жаберную занавес Респираторные поверхности постоянно контактируют с новой кислородной водой. [ 35 ] [ 36 ]
Кормление и диета
[ редактировать ]Саламандры - оппортунистические хищники . Как правило, они не ограничиваются конкретными продуктами, но питаются практически любым организмом разумного размера. [ 37 ] Большие виды, такие как японский гигант саламандре ( Andrias Japonicus ), едят крабов, рыб, мелких млекопитающих, амфибий и водных насекомых. [ 38 ] В исследовании небольших темных саламандров ( desmognathus ) в Аппалачских горах их диета включает в себя дождевые черви , мухи , жуки , личинки жуков, подходы листовые , весенние хвосты , мотыльки , пауки , кузнечики и клещи . [ 37 ] Иногда каннибализм происходит, особенно когда ресурсы короткие или время ограничены. Голочи в эфемерных бассейнах тигров в эфемерных бассейнах иногда прибегают к употреблению друг друга и, по -видимому, способны нацеливаться на не связанных с ними людей. [ 39 ] Взрослые Blackbelly Salamanders ( Desmognathus Quadramaculatus ) добывают взрослых и молодых других видов саламандров, в то время как их личинки иногда каннибализируют меньшие личинки. [ 40 ]

Большинство видов саламандры имеют маленькие зубы как в верхних, так и в нижних челюстях. В отличие от лягушек , даже личинки саламандров обладают этими зубами. [ 8 ] Хотя личиночные зубы формируются как заостренные шишки, зубы взрослых адаптированы, чтобы они могли легко понять добычу. Корона . , которая имеет два шин (BICUSPID), прикреплена к цветоножной волокнам коллагеновыми волокнами Сустав, образованный между двумя и цветоножными, является частично гибким, так как он может сгибаться внутрь, но не наружу. Когда борьба с добычей продвигается во рту саламандры, кончики зубов расслабляются и сгибаются в том же направлении, поощряя движение к горлу и сопротивляясь побегу добычи. [ 41 ] Многие саламандры имеют пятна зубов, прикрепленные к Вомеру и недобывающие кости в крыше рта, и они помогают сохранить добычу. Все виды зубов резоргируются и заменяются с интервалами на протяжении всей жизни животного. [ 42 ]
Земная саламандре ловит свою добычу, выбивая липкий язык в действии, которое занимает менее чем на полсекунды. У некоторых видов язык прикреплен спереди к полу рта, в то время как в других он монтируется на цветоноселе. Он оказывается липким от выделения слизи из желез в его кончике и на крыше рта. [ 43 ] Высокоскоростная кинематография показывает, как тигровая саламандра ( Ambystoma tigrinum ) позиционирует себя с ее мордой близко к ее добыче. Затем его рот широко растет, нижняя челюсть остается неподвижной, а язык вытушивает и изменяет форму, когда он стреляет вперед. Выпяченный язык имеет центральную депрессию, и обод этого рушится внутрь, когда цель поражена, задерживая добычу в корыте, наполненной слизью. Здесь он удерживается, в то время как шея животного согнута, язык втягивается и закрывается челюсти. Большая или устойчивая добыча сохраняется зубами, в то время как повторные выступы и втягивания языка втягивают его. Шлота включает в себя альтернативное сокращение и расслабление мышц в горле, которым помогает депрессия глазных яблок в крышу во рту. [ 44 ] Многие саламандры без легких из семейства Plethodontidae имеют более сложные методы кормления. Мышцы, окружающие сжиму поджоидной кости для хранения эластичной энергии в пружинной соединительной ткани, и фактически «выстрелили» под горную кость изо рта, тем самым удлиняя язык. [ 45 ] [ 46 ] Мышцы, которые возникают в области таза и вставки в язык, используются, чтобы отводить язык, а подкоим подложку к их первоначальным положениям. [ 47 ]
Водной саламандре не хватает мышц на языке и захватывает свою добычу совершенно по -разному. Он захватывает продукт, охватывает его зубами и принимает своего рода инерционное кормление. Это включает в себя бросок головы, резко вытягивая воду в рту и за его пределами, и щелкает челюсти, все из которых, как правило, рвут и мацепируют добычу, которая затем проглатывается. [ 47 ]
Несмотря на то, что они часто питаются медленно движущимися животными, такими как улитки , креветки и черви , сирениды уникальны среди саламандров для развития видообразования в отношении травоядных, таких как концы челюсти, похожие на клюв и обширные кишечники. Они питаются водорослями и другими мягкими растениями в дикой природе и легко едят предлагаемый салат . [ 48 ]
Защита
[ редактировать ]Саламандры имеют тонкие шкуры и мягкие тела, и движутся довольно медленно, и на первый взгляд может показаться уязвимым для оппортунистического хищничества. Однако у них есть несколько эффективных линий защиты. Слизистое покрытие на влажной коже затрудняет их понимание, а слизистое покрытие может иметь оскорбительный вкус или быть токсичным. При нападении хищника саламандре может позиционировать себя, чтобы заставить главного ядовитого железы встретиться с агрессором. Часто они находятся на хвосте, которые могут быть покатаны или выдвинуты и выгнуты над спиной животного. Жертва хвоста может быть достойной стратегией, если саламандр убегает со своей жизнью, и хищник учится избегать этого вида саламандре в будущем. [ 49 ]
Апосематизм
[ редактировать ]
Было показано, что секреция кожи саламандры Tiger ( Ambystoma tigrinum ), питаемые крысам, вызывают отвращение к вкусу, и крысы избежали презентационной среды, когда она была им снова предложена. [ 50 ] Пожарная саламандра ( Саламандра Саламандра ) имеет хребет с большими гранулированными железами вниз по позвоночнику, которые способны впрыгивать тонкую реактивную реакцию токсичной жидкости у своего нападающего. Проведя его тело, он может точно направлять спрей на расстояние до 80 см (30 дюймов). [ 51 ]
Иберийский ребристый Newt ( Pleurodeles Waltl ) имеет еще один метод сдерживания агрессоров. Его кожа источает ядовитую, вязкую жидкость и в то же время, Newt вращает свои резко заостренные ребра через угол между 27 и 92 ° и принимает завышенную осанку. Это действие заставляет ребра проколоть стенку тела, каждое ребро, выступающее через оранжевую бородавку, расположенную в боковом ряду. Это может обеспечить апонемтический сигнал, который делает шипы более заметными. Когда опасность прошла, ребра убираются, а кожа заживает. [ 52 ]
Камуфляж и мимика
[ редактировать ]Хотя многие саламандры имеют загадочные цвета, чтобы быть незаметными, другие сигнализируют о своей токсичности благодаря своей яркой окраске . Желтый, оранжевый и красный - это цветы, которые обычно используются, часто с черным для большего контраста. Иногда позы животных, если они подвергаются нападению, раскрывая вспышку предупреждающего оттенка на его нижней стороне. Красный EFT, ярко окрашенная наземная форма ювенильной формы восточного Ньюта ( Notophthalmus viridescens ), очень ядовита. Его избегают птиц и змей, и он может выжить в течение 30 минут после проглатывания (позже извергнуто). [ 53 ] Красная саламандре ( Pseudotriton Ruber ) - это приемлемый вид с аналогичной окраской для красного EFT. Хищники, которые ранее питались на нем, было показано, чтобы избежать его после встречи с красными эфтами, пример мимики Бейтсиана . [ 53 ] Другие виды демонстрируют аналогичную мимику. В Калифорнии приятный желтоглазый саламандр ( Ensatina Eschscholtzii ) напоминает токсичную Калифорнийскую Ньют ( Taricha Torosa ) и грубого кожа Newt ( Taricha Granulosa ), тогда как в других частях его диапазона он загадочен. [ 54 ] Существует корреляция между токсичностью калифорнийских видов саламандре и суточными привычками: относительно безвредные виды, такие как калифорнийский стройный саламандре ( Batrachoseps Attenuatus ), являются ночными и едят змеи, в то время как в калифорнийском Ньюте есть много больших ядовитых желез на его коже, джорт,, джонки,, джонки,, джонки,, джонки, динтуальные, джонки, динтурские, джонки, джори и избегается змеями. [ 55 ]
Аутотомия
[ редактировать ]Некоторые виды саламандры используют аутотомию хвоста, чтобы избежать хищников. Хвост падает и парят на некоторое время после атаки, и саламандр либо убегает, либо остается настолько, чтобы не быть замеченным, пока хищник отвлекается. Хвост отрастает со временем, и саламандры регулярно регенерируют другие сложные ткани, включая линзу или сетчатку глаза. Всего за несколько недель после потери кусочки конечности саламандр прекрасно реформирует недостающую структуру. [ 56 ]
Распределение и среда обитания
[ редактировать ]Саламандры откладываются от других амфибий во время середины и позднего перм и первоначально были похожи на современных членов Cryptobranchoidea . Их сходство с ящерицами является результатом симплсуоморфии , их общее сохранение примитивного плана тела тетрапод, но они не более тесно связаны с ящерицами, чем с млекопитающими. Их ближайшие родственники - это лягушки и жабы, в рамках Батрахии .
Самая старая известная общая группа ( Caudata ) Саламандра-это триассурус из триаса Кыргызстана . [ 57 ] Дальнейшие окаменелости саламандры известны из среднего юрского Англии периода , [ 58 ] Шотландия , [ 59 ] Китай , [ 60 ] и Казахстан . [ 61 ] Самая старая известная саламандр короны ( Urodela но недавние анализы предполагают, что это Valdotriton от поздней юры Испании ) остается неопределенным , . [ 59 ]
Саламандры встречаются только в холрктике и неотропических регионах, не достигающих к югу от Средиземного бассейна , Гималаев или в Южной Америке в бассейне Амазонки . Они не простираются к северу от линии арктических деревьев , с самыми северными азиатскими видами Salamandrella Keyserlingii , которые могут пережить долгосрочное замораживание при -55 ° C, [ 62 ] Происходящие в сибирских лиственных лесах саха и самых северных видах в Северной Америке, Ambystoma Loterale , не достигая дальше на север, чем Лабрадор и Тарича Гранулеза, не за пределами Аляски . [ 63 ] У них было исключительно Лауразианское распределение, пока Болитоглосса не вторглась в Южную Америку из Центральной Америки, вероятно, к началу раннего миоцена , около 23 миллионов лет назад. [ 64 ] Они также жили на Карибских островах во время ранней эпохи миоцена , подтвержденную открытием палеоптодонских латинолье . [ 65 ] найден в ловушке в Янтаре в Доминиканской Республике . Окаменелости позвонков, извлеченных из места окаменелостей Мургона, были предварительно приписаны на саламандре, [ 66 ] хотя его истинная идентичность оспаривается. Если позвонки действительно принадлежат саламандре, они будут представлять собой единственные саламандры в Австралии .
Есть около 760 живых видов саламандры. [ 67 ] [ 68 ] Одна треть известных видов саламандры встречается в Северной Америке. Самая высокая концентрация из них находится в области Аппалачских гор, где, как полагают, Plethodontidae возникли в горных ручьях. Здесь зоны растительности и близость к воде имеют большее значение, чем высота. Только виды, которые приняли более наземный способ жизни, смогли рассеять в другие населенные пункты. Северная слизистая саламандре ( глютиноз Plethodon ) имеет широкий диапазон и занимает среду обитания, похожую на южную серую саламандре ( Plethodon Metcalfi ). Последнее ограничено немного более холодными и более влажными условиями в северо-северо- бухте северных , но какой-то неизвестный фактор, по-видимому, предотвращает сосуществование двух видов. [ 37 ]
Один вид, саламандра Андерсона , является одним из немногих видов живых амфибий, которые встречаются в солоноватой или соленой воде. [ 69 ]
Размножение и развитие
[ редактировать ]
Многие саламандры не используют вокализации, [ 70 ] И у большинства видов полы выглядят одинаково, поэтому они используют обонятельные и тактильные сигналы для выявления потенциальных товарищей, и половой отбор происходит . Феромоны играют важную роль в процессе и могут быть произведены брюшной железой у мужчин, а также клоакальными железами и кожей у обоих полов. Иногда мужчины рассматриваются по исследуемым потенциальным товарищам со своими мордами. В Старом Свете, Triturus spp., Мужчины сексуально диморфны и демонстрируют перед женщинами. Визуальные сигналы также считаются важными у некоторых видов Plethodont . [ 71 ]
За исключением наземных видов в трех семействах Plethodontidae , Ambystomatidae и Salamandridae , саламандры спариваются в воде. [ 72 ] Коричневое спаривание варьируется от ухаживания между одним мужчинами и женщинами до взрывной группы разведения. [ 73 ] В Clade Salamandroidea , которая составляет около 90% всех видов, оплодотворение является внутренним. [ 74 ] Как правило, саламандры с внутренним оплодотворением имеют косвенную перенос сперматозоидов, но у саламандре сардинского ручья , саламандре корсиканского ручья , кавказской саламандре и саламандре пиренана , мужчина передает свою сперму непосредственно в женскую клока. [ 75 ] [ 76 ] [ 77 ] Для вида с косвенным переносом сперматозоидов мужчина откладывает сперматофор на земле или в воде в соответствии с видами, и самка поднимает это своим вентиляционным отверстием. Сперматофор имеет пакет сперматозоидов, поддерживаемый на конической желатиновой основе, и часто сложное поведение ухаживания участвует в его осаждении и сборе. Оказавшись внутри Cloaca, сперматозоиды перемещаются в сперматеку , одну или несколько камер на крыше клоаки, где они хранятся в течение иногда длительных периодов, пока яйца не будут отложены. В азиатских саламандерах гигантские саламандры и Sirenidae , которые являются наиболее примитивными группами, оплодотворение является внешним. В репродуктивном процессе, сходном с процессом типичных лягушек, мужчина выпускает сперму на массу яиц. У этих саламандров также есть мужчины, которые проявляют родительскую помощь , которая в противном случае встречается только у женщин с внутренним оплодотворением. [ 71 ] [ 78 ]
Происходят три различных вида осаждения яиц. Ambystoma и Taricha Spp. Появится большое количество маленьких яиц в тихих прудах, где многие большие хищники маловероятны. Большинство сумеречных саламандров ( desmognathus ) и гигантского гиганта тихоокеанских саламандров ( дикамптодон ) лежат меньшие партии яиц среднего размера в скрытой площадке в текущей воде, и они обычно охраняются взрослыми, обычно самка. Многие из тропических саламандров ( Bolitoglossa ) и саламандров без легких (Plethodontinae) откладывают небольшое количество больших яиц на суше в хорошо скрытом месте, где они также охраняются матерью. [ 71 ] Некоторые виды, такие как пожарные саламандры ( саламандры ), являются овавивипарусными , а самка сохраняет яйца внутри ее тела до тех пор, пока они не вылуплены, либо в личинки, которые будут осаждены в корпусе воды, или в полностью сформированные несовершеннолетние. [ 8 ]
В умеренных регионах воспроизведение обычно является сезонным, и саламандры могут мигрировать на местах размножения. Мужчины обычно прибывают первыми, а в некоторых случаях создают территории . Как правило, следуют личиночной стадии, в которой организм полностью вод. Говага имеет три пары внешних жабр, без век, длинное тело, сплюснутый в боковом хвосте с дорсальными и вентральными плавниками, а также у некоторых видов конечностей или конечностей. Личинки пруда могут иметь пару балансировщиков, подобных стержням по обе стороны от головы, длинные жабры и широкие плавники. Личинки типа ручья являются более тонкими с короткими жаберными нитями-в Rhyacotriton и Onychodactylus, и некоторые виды в Batrachuperus, жабры и жабры чрезвычайно уменьшены, чрезвычайно уменьшены. [ 79 ] Более узкие плавники и без балансировщиков, но вместо этого уже развиваются задние конечности, когда они вылупится. [ 80 ] Головастики плотоядные , а личиночная стадия может длиться от дней до многих лет, в зависимости от видов. Иногда эта стадия полностью обходит, и яйца большинства саламандров без легких (Plethodontidae) развиваются непосредственно в миниатюрные версии взрослого без промежуточной личинки. [ 81 ]
К концу стадии личинки у головастиков уже есть конечности, и метаморфоза происходят нормально. В Саламандре это происходит в течение короткого периода времени и включает в себя закрытие прорезей жабры и потерю структур, таких как жабры и хвостовые плавники, которые не требуются в качестве взрослых. В то же время развиваются веки, рот становится шире, появляется язык и образуются зубы. Водная личинка появляется на земле как земного взрослого. [ 82 ]

Не все виды саламандров следуют по этому пути. Neoteny , также известный как педорфоза, наблюдалась во всех семействах саламандре и может быть повсеместно возможна во всех видах саламандре. В этом состоянии человек может сохранять жабры или другие юношеские черты, достигая репродуктивной зрелости. Изменения, которые происходят при метаморфозе, находятся под контролем гормонов щитовидной железы , а в облигатных неотенах, таких как аксолотль ( Ambystoma mexicanum ), ткани, по -видимому, не реагируют на гормоны. У других видов изменения могут не быть вызваны недостаточной механизмом гипоталамуса-гипофизарно-оборота, который может возникнуть, когда условия в наземной среде слишком раздражительны. [ 82 ] Это может быть связано с холодными или дико колеблющимися температурами, засушкой, отсутствием пищи, отсутствием укрытия или недостаточным йодом для образования гормонов щитовидной железы. Генетика также может сыграть роль. личинки саламандров тигров ( Ambystoma tigrinum Например, ) развиваются вскоре после вылупления и в сезонных бассейнах быстро подвергаются метаморфозу. Другие личинки, особенно в постоянных бассейнах и более теплом климате, могут не подвергаться метаморфозу до полной взрослой по размеру. Другие популяции в более холодном климате могут вообще не метаморфоза вообще и становятся сексуально зрелыми, в то время как в их личиночных формах. Neoteny позволяет видам выжить, даже когда наземная среда слишком резкая для взрослых, чтобы процветать на суше. [ 80 ]
Сохранение
[ редактировать ]
Общее снижение живых видов амфибий было связано с грибковым болезнью читридиомикоза . Более высокая доля видов саламандры, чем лягушек или кейцилиан, находится в одной из категорий риска, установленной МСОП . Саламандры показали значительное уменьшение количества за последние несколько десятилетий 20 -го века, хотя не было обнаружено прямой связи между грибом и снижением населения. [ 83 ] МСКН предпринял дальнейшие усилия в 2005 году, поскольку они создали План действий по сохранению амфибий (ACAP), за которым впоследствии последовали амфибийский ARK (AARK), специализированная группа амфибий (ASG) и, наконец, организация зонтика, известная как Альянс выживания амфибий (ASA ) [ 84 ] Исследователи также ссылаются на выручку лесов , что приводит к фрагментации подходящих мест обитания и изменению климата в качестве возможных факторов. Такие виды, как Psedoeurycea Brunnata и Pseudoeurycea Goebeli , которые были в изобилии в облачных лесах Гватемалы и Мексики в 1970 -х годах, были обнаружены к 2009 году, чтобы быть редкими. [ 85 ] Тем не менее, за эти годы было собрано лишь немногие данные, и благодаря интенсивным обследованию исторических и подходящих новых мест было возможно найти особей других видов, таких как Parvimolge Townsendi , которые считались вымершими . [ 83 ] В настоящее время основные линии защиты для сохранения саламандров включают как на in situ , так и ex situ методы сохранения. Предпринимаются усилия для некоторых членов семейства Саламандре, которые должны быть сохранены в рамках программы разведения по сохранению (CBP), но должны быть проведены исследования заранее, чтобы определить, собираются ли виды саламандры на самом деле ценность из CBP, поскольку исследователи имеют отметил, что некоторые виды амфибий полностью терпят неудачу в этой среде. [ 84 ]
Различные инициативы по сохранению предпринимаются во всем мире. Китайский гигант Саламандре , на уровне 1,8 м (6 футов), самый большой амфибия в мире, подвергается критической подростке , поскольку она собирается для еды и для использования в традиционной китайской медицине . Программа экологического образования проводится для поощрения устойчивого управления дикими популяциями в горах Цинлинг , и были созданы программы размножения в неволе. [ 86 ] Адский бендер -еще один большой, долгоживущий вид с сокращающимися числами и меньшим количеством несовершеннолетних, достигающих зрелости, чем ранее. [ 87 ] Другим тревожным выводом является увеличение аномалий в до 90% популяции адского в весеннем водоразделе в Арканзасе. [ 88 ] Потеря среды обитания, зажигание ручьев, загрязнение и заболевание были вовлечены в снижение, и программа размножения в неволе в зоопарке Сент -Луиса . была успешно создана [ 89 ] Из 20 видов мельчайших саламандров ( Thorius spp.) В Мексике, как полагают, вымерли, а большинство других находятся под угрозой исчезновения. Конкретные причины снижения могут включать в себя изменение климата, хитридиомикоз или вулканическую деятельность, но основной угрозой является разрушение среды обитания , поскольку лесозаготовки, сельскохозяйственная деятельность и поселение человека уменьшают их часто крошечные, фрагментированные диапазоны. Работа обследования проводится для оценки статуса этих саламандров и лучше понять факторы, связанные с сокращением их населения, с целью принятия мер. [ 90 ]
Ambystoma Mexicanum , водный саламандр, является видом, защищенным под мексиканским UMA (подразделение по управлению и сохранению дикой природы) по состоянию на апрель 1994 года. Другой вредный фактор заключается такие как Нил Тилапия и Карп. Тилапия и карп непосредственно конкурируют с аксолотлами, потребляя свои яйца, личинки и несовершеннолетних. Изменение климата также сильно повлияло на аксолотли и их популяции по всей области южной части Мексики. Из -за своей близости к Мехико чиновники в настоящее время работают над программами на озере Xochimilco, чтобы привлечь туризм и обучение местного населения о восстановлении естественной среды обитания этих существ. [ 91 ] Эта близость является большим фактором, который повлиял на выживание аксолотла, поскольку город расширился, чтобы захватить регион Xochimilco, чтобы использовать свои ресурсы для воды, обеспечения и сточных вод. [ 92 ] Тем не менее, аксолотл имеет преимущество в том, что он поднимается на фермах с целью исследовательских учреждений. Таким образом, все еще есть шанс, что они смогут вернуться к своей естественной среде обитания. Недавнее снижение численности населения существенно повлияло на генетическое разнообразие среди популяций аксолотля, что затрудняет дальнейшее развитие научно. Важно отметить, что, хотя существует уровень ограниченного генетического разнообразия из -за популяций амбистомы , таких как аксолотль, являясь падеоморфными видами, это не учитывает общее отсутствие разнообразия. Существуют доказательства, указывающие на историческую узкую жизнь Амбистомы , которая способствует вариационным вопросам. К сожалению, нет большого генетического бассейна для вида, чтобы вытащить, в отличие от исторических времен. Таким образом, существует серьезная обеспокоенность за инбридинги из -за отсутствия потока генов. [ 93 ] Одним из способов изучения исследователей поддержание генетического разнообразия в популяции является криоконсервация сперматофоров от мужского аксолотля. Это безопасный и неинвазивный метод, который требует сбора сперматофоров и ставит их в глубокую замораживание для сохранения. Самое главное, они обнаружили, что при оттаивании существует ограниченный ущерб сперматофорам, и, таким образом, это жизнеспособный вариант. По состоянию на 2013 год это метод, который используется для спасения не только аксолотла, но и многих других членов семьи Саламандре. [ 92 ] [ 94 ] [ 95 ]
Исследования проводится по сигналам окружающей среды, которые должны быть воспроизведены, прежде чем пленные животных можно убедить размножаться. Общие виды, такие как Tiger Salamander и Mudpuppy, получают гормоны для стимулирования производства сперматозоидов и яиц, и роль аргининового вазотоцина в поведении в ухаживание исследуется. Другой линейкой исследований является искусственное оплодотворение , либо in vitro , либо путем вставки сперматофоров в клоаки женщин. Результаты этого исследования могут быть использованы в программах размножения в неволе для исчезающих видов. [ 96 ]
Таксономия
[ редактировать ]Название заказа Urodela происходит от имени Urodèles, данного Андре Мари Константа Duméril в 1805 году, [ 2 ] Это происходит от греческих слов из « хвоста » и Δῆλος dēlos «видимых, заметных» из -за их «постоянных» хвостов. [ 97 ]
Разногласия существуют среди разных властей в отношении определения терминов Caudata и Urodela. Некоторые утверждают, что уродела должна быть ограничена группой Crown , при этом Caudata используется для общей группы. [ 2 ] [ Цитация необходима ] Другие ограничивают название Caudata в группе Crown и используют Urodela для общей группы. [ 98 ] [ 99 ] Первый подход кажется наиболее широко принятым и используется в этой статье. [ 68 ]
Десять семей, принадлежащих к Уроделе, разделены на три подчинения. [ 98 ] Clade Neocauda имеет использование Cryptlenchoidea и Salamangonroidea от Syrenoidea.
| Cryptobranchoidea (гигантские саламандры) | |||
| Семья | Общие имена | Пример видов |
Пример изображения |
|---|---|---|---|
| Cryptobranchidae | Гигантские саламандры | Hellbender ( Cryptobranchus Alleganiensis ) | |
| Hynobiidae | Азиатские саламандры | Хида Саламандр ( Hynobius kimurae ) | 
|
| Salamandroidea (продвинутые саламандры) | |||
| Ambystomatidae | Крот саламандерс | Мраморная саламандре ( Амбистома непрозрачный ) | 
|
| Amphiumidae | Амфия или Конго угры | AMPHIUMA с двумя точками ( AmphiumA означает ) | 
|
| PLETHODONTIDAE | Саламандры без легких | Красная саламандра ( Plethodon Cinereus ) | 
|
| Proteidae | Грязные тупики и олмс | OLM ( Proteus a Ejon ) | |
| Rhyacotritonidae | Торрент саламандерс | Southern Torrent Salamander ( Rhyacotriton variegatus ) | 
|
| Salamandridae | Newts и True Salamanders | Alpine Newt ( Ichthyosaura alpestris ) | |
| Sirenoida (сирены) | |||
| Sirenidae | Сирены | Большая сирена ( Сирена Лакертина ) |
Филогения и эволюция
[ редактировать ]Происхождение и эволюционные отношения между тремя основными группами амфибий (гимнофонаны, уродели и анураны) являются вопросом дебатов. Молекулярная филогения 2005 года, основанная на анализе РДНК , предположила, что первая дивергенция между этими тремя группами произошла вскоре после того, как они разветвлялись от рыб с долемы на Девониане (около 360 миллионов лет назад), и до разрыва суперконтинента Пангея . Краткость этого периода и скорость, с которой имело место излучение, могут помочь объяснить относительный дефицит окаменелостей амфибий, которые, по -видимому, тесно связаны с Lissamphians . [ 100 ] Тем не менее, более поздние исследования обычно обнаруживались более поздние (позднее каменноугольное. [ 101 ] Пермски [ 102 ] ) Возраст для самой базальной дивергенции среди лиссамбийцев.

Самый ранний известный лиссамппольский саламандр-это триасасур из среднего триаса Кыргызстана. [ 103 ] Другие ископаемые саламандры известны из юрского периода Евразии, в том числе гонорар Кокартуса из средней юры Кыргызстана, два вида, по-видимому, неотенического, водного мармореретона из среднего юрского периода Англии и Шотландии, [ 104 ] и Карауру из среднего юрского периода Казахстана напоминал современные саламандры в морфологии и, вероятно, имел аналогичный стиль жизни. [ 68 ] Они выглядели как надежные современные саламандры, но не имели ряда анатомических особенностей, которые характеризуют все современные саламандры. [ 105 ]
Две группы существующих саламандров - это Cryptobranchoidea (в том числе азиатские и гигантские саламандры) и саламандроидэ (которая включает в себя все остальные живые саламандры), также известные как Diadectosalamandroidei. Обе группы известны из юрского периода в Китае. Первый был примером Чунерпетона Tianyiensis , Pangerpeton Sinensis , Jeholotriton Paradoxus , Regalerpeton weichangensis , Liaoxitriton daohugouensis и Iridotriton Hechti , а также с первым от Beianerpeton jianpingensis . На верхнемееловый , большинство или все живые семьи Саламандре, вероятно, появились. [ 68 ]
Следующая кладограмма показывает взаимосвязь между семьями саламандры на основе молекулярного анализа Pyron and Wiens (2011). [ 106 ] Положение Sirenidae оспаривается, но позиция в качестве сестры в саламандроиде лучше всего подходит для молекулярных и ископаемых доказательств. [ 68 ]
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Геном и генетика
[ редактировать ]Саламандры обладают гигантскими геномами, охватывающими от 14 до 120 ГБ. [ 107 ] ( Человеческий геном длиной 3,2 ГБ). Геномы плевроделей Waltl (20 ГБ) и Ambystoma Mexicanum (32 ГБ) были секвенированы. [ 108 ] [ 109 ]
Их гигантские геномы сильно повлияли на их физиологию. Это включает в себя их скелетные и кровообратные системы и привело к упрощенному мозгу, слабому сердцу и медленному метаболизму. [ 110 ] Клеточные механизмы, которые предотвращают накапливаться транспозоны, представляются частично дефектом в саламандре. Некоторые виды с самыми большими геномами потеряли способность проходить метаморфозу. Развитие тела медленнее, чем его рост по сравнению с их предками, и останавливается в определенном возрасте, оставляя им эмбриональные черты. Ткани саламандры содержат клетки, которые дифференцируются медленно, слабо или совсем не вообще из -за задержки интрона, которая дает им регенеративные свойства, которые включают регенерирующие части лица и глаза, легкие, печень, сердце и даже спинной мозг и мозг , и они были описаны как «ходячие мешки с стволовыми клетками». [ 111 ] [ 112 ] [ 113 ] Исследования также показали, что у них не разрабатываются типичные признаки старения и не накапливают возрастные заболевания, такие как рак. [ 114 ]
В человеческом обществе
[ редактировать ]Миф и легенда
[ редактировать ]
Легенды развивались вокруг Саламандры на протяжении веков, многие из которых были связаны с огнем. Это соединение, вероятно, происходит от тенденции многих саламандров останавливаться внутри гниющих бревна. Когда бревна была помещена в огонь, саламандре попыталась бы сбежать, придавая доверие к убеждению, что саламандры были созданы из пламени. [ 115 ]
Ассоциация саламандры с огнем впервые появилась в древности с Аристотелем ( История животных 5, 17) и с Плинием, старшим писанием в своей естественной истории (10, 86), что «саламандре настолько холод . [ 116 ] Способность выпускать огонь повторяется святым Августином в пятом веке и Исидоре Севильи в седьмом веке. [ 117 ] [ 118 ]

У мифического правителя, предварительного Джона, предположительно, была одежда, сделанная из предполагаемых волос саламандры, на самом деле асбестоссового волокна, уже известного древней Грецией и Римом (Линам Вивум Плиния Старшая Историлия натуралиса , 19, 4). [ 119 ] «Император Индии» обладал иском из тысячи шкур; У папы Александра III была туника, которую он высоко ценил, и Уильям Кэкстон (1481) писал: «Этот Салеманр Берите Вул, из которого сделана ткань и гирдлы, которые не могут Бренна в Файре». [ 120 ] Говорят, что саламандр был настолько токсичен, что, повернув вокруг дерева, он мог отравить фрукты и так убить любого, кто их съел, и, упав в колодец, мог убить всех, кто выпил из него. [ 120 ]
В автобиографии , приветствует Cellini, рассказывает:
Когда мне было около пяти лет, мой отец сидел один в одной из наших маленьких комнат, пел и играл в его алис. Там только что было сделано там, и хороший бревенчатый огонь все еще горел. Было очень холодно, и он притянул возле огня. Затем, когда он смотрел на пламя, его глаза упали на маленькое животное, как ящерицу, которое весело бегало в самой жаркой части огня. Внезапно осознав, что это было, он позвонил моей сестре и мне и показал это нам. А потом он дал мне такую сильную коробку на ушах, что я закричал, и разразился слезами. При этом он успокоил меня как можно более любезно, и сказал: «Мой дорогой маленький мальчик, я не ударил тебя, потому что ты сделал неправильно. Я только сделал это, чтобы вы никогда не забудете, что ящерица, которую вы видели в огне, является саламандре, и, насколько мы знаем, наверняка никто никогда не видел его раньше ». [ 121 ]
Японский гигант Саламандр был предметом легенды и произведений искусства в Японии (например, Утагава работа Куниёси ). Известное японское мифологическое существо, известное как Каппа, может быть вдохновлено этой саламандре. [ 122 ]
Медицинские исследования
[ редактировать ]Саламандерса Регенерация конечностей уже давно представляла интерес среди ученых. Первое обширное исследование на уровне клеток было проведено Vincenzo Colucci в 1886 году. [ 123 ] Исследователи пытались выяснить условия, необходимые для роста новых конечностей, и надеются, что такая регенерация может быть воспроизведена у людей с использованием стволовых клеток . Аксолотлы использовались в исследованиях и были генетически спроектированы, чтобы флуоресцентный белок присутствовал в клетках в ноге, что позволяет отслеживать процесс деления клеток под микроскопом. Похоже, что после потери конечности клетки объединяются, чтобы сформировать кухню, известный как бланка . Это поверхностно кажется недифференцированным, но клетки, которые возникли в коже, впоследствии превращаются в новую кожу, мышечные клетки в новые мышечные и хрящевые клетки в новый хрящ. Только клетки сразу под поверхностью кожи, которые являются плюрипотентными и способными развиваться в любом типе клетки. [ 124 ] Исследователи из Австралийского института регенеративной медицины обнаружили, что когда макрофаги были удалены, саламандры утратили способность регенерации и вместо этого образовали рубцовую ткань. Если процессы, участвующие в формировании новой ткани, могут быть обработаны в людях, возможно, может быть возможным лечить травмы спинного мозга или мозга, восстановить поврежденные органы и уменьшить рубцы и фиброз после операции. [ 125 ]
Пятнистая саламандре (Amblystoma maculatum) живет в симбиотических отношениях с зелеными водорослями, известными как oophila amblystomatis. Клетки водорослей попадают в тканевые клетки по всему телу эмбриона и, по -видимому, избегают отторжения, активируя гены, которые подавляют иммунный ответ эмбриона. Механизм, который можно использовать при лечении от аутоиммунных заболеваний у людей. [ 126 ]
бренди
[ редактировать ]В статье 1995 года в журнале Sloven Weekly Mradina сообщила Salamander Brandy, ликер, предположительно коренной для Словении . Говорят, что он объединяет галлюциногенный с афродизиакскими эффектами и производится путем помещения нескольких живых саламандров в бочку ферментирующих фруктов. Стимулируемые алкоголем, они выделяют токсичную слизь в защите и в конечном итоге умирают. Говорят , что, помимо вызывания галлюцинаций, нейротоксины, присутствующие в варелке, вызывают крайнее сексуальное возбуждение . [ 127 ]
Позднее исследование словенского антрополога Миха Козорог ( Университет Любляны ) рисует совершенно другую картину - саламандр в Бренди, по -видимому, традиционно рассматривался как прелюбодеящий , который вызвал плохое здоровье. Он также использовался в качестве термина клеветы. [ 128 ]
Ссылки
[ редактировать ]Цитаты
[ редактировать ]- ^ Цзя, Цзя; Андерсон, Джейсон С.; Гао, Ке-Кин (23 июля 2021 года). «Средние юры гинобииды из Китая проливают свет на эволюцию базальных саламандров» . ISCAING . 24 (7): 102744. Bibcode : 2021isci ... 24J2744J . doi : 10.1016/j.isci.2021.102744 . ISSN 2589-0042 . PMC 8264161 . PMID 34278256 .
- ^ Jump up to: а беременный в Фрост, Даррел Р. (2023) [1998]. "Caudata" . Амфибийские виды мира . Американский музей естественной истории . Архивировано из оригинала 22 марта 2023 года . Получено 22 марта 2023 года .
- ^ Следы жизни побережья Джорджии: раскрытие невидимой жизни растений и животных
- ^ «Пропавшие части? Секрет регенерации Саламандры раскрыл» . Живая наука . 20 мая 2013 года. Архивировано с оригинала 16 июля 2018 года . Получено 15 июля 2018 года .
- ^ Чжун, Джиксинг; Айрес, Рита; Тишиос, Джорджиос; Скуфа, Евангелия; Брандт, Керстин; Сандовал-Гузман, Татьяна; Азтекин, Кан (10 октября 2023 г.). «Атлас мультиспецифический атлас решает и парадокс развития и регенерации аксолотла » Природная связь 14 (1): Bibda : 2023natco..14.6346Z 6346. Doi : 10.1038/s41467-023-41944-w . PMC 10564727 PMID 3781673
- ^ Харпер, Дуглас. "Саламандре" . Онлайн этимологический словарь . Получено 22 марта 2023 года .
- ^ Лидделл, Генри Джордж; Скотт, Роберт (1940). "σαλαμάνΔρα" . Грек-английский лексикон . Цифровая библиотека Персея. Архивировано из оригинала 12 марта 2023 года . Получено 22 марта 2023 года .
- ^ Jump up to: а беременный в дюймовый и фон Lanza, B.; Ванни, с.; Nistri, A. (1998). Cogger, Hg; Zweifel, RG (Eds.). Энциклопедия рептилий и амфибий . Академическая пресса. С. 60–68. ISBN 978-0-12-178560-4 .
- ^ Повторные экологические и жизненные переходы делают саламандры идеальной моделью для эволюции и развития
- ^ Метаболизм, газообмен и кислотный баланс гигантских саламандров
- ^ Полевое руководство по амфибиям и рептилиям Калифорнии: пересмотренное издание
- ^ Stebbins & Cohen (1995) с. 3
- ^ Лопес, Карл Х.; Броди, Эдмунд Д. младший. (1972). «Функция Costal Grooves (амфибия)» Журнал герпеологии 11 (3): 372–374. doi : 10.2307/1563252 . JSTOR 1563252 .
- ^ Jump up to: а беременный в Stebbins & Cohen (1995) с. 26–30
- ^ Jump up to: а беременный Stebbins & Cohen (1995) с. 10–16
- ^ Броди, Эдмунд Д. младший (1968). «Исследования на коже токсине взрослого грубого кожа Newt, Taricha Granulosa ». Копея . 1968 (2): 307–313. doi : 10.2307/1441757 . JSTOR 1441757 . S2CID 52235877 .
- ^ Jump up to: а беременный Уэллс, Кентвуд, Д. (2010). Экология и поведение амфибий . Университет Чикагской Прессы. С. 411–417. ISBN 978-0-226-89333-4 .
{{cite book}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Леон, Иезкиль Гонсалес; Ramírez-Pinilla, Марта Патриция (2009). «Ментальная железа Bolitoglossa Nicefori (Flow: Plethodontidae)» . Амфибийская рефилия . 30 (4): 561–569. Doi : 10.1163/156853809789647013 .
- ^ Stebbins & Cohen (1995) с. 37–40
- ^ Stebbins & Cohen (1995) с. 42–44
- ^ Przyrembel, C.; Келлер, Б.; Neumeyer, C. (1995). «Трихроматическое цветовое зрение в саламандре ( Саламандра Саламандра )». Журнал сравнительной физиологии . 176 (4): 575–586. doi : 10.1007/bf00196422 . S2CID 749622 .
- ^ Schoch, Rainer R. (19 марта 2014 г.). Эволюция амфибий: жизнь ранних земельных позвоночных . Джон Уайли и сыновья. ISBN 9781118759134 Полем Получено 15 июля 2018 года - через Google Books.
- ^ «Исследователи показывают, как развивалось слух» . sciencedaily.com . Архивировано из оригинала 16 июля 2018 года . Получено 15 июля 2018 года .
- ^ Беккер, RP; Lombard, Re (1977). «Структурные корреляты функции в мышцах« opercularis »амфибий». Клеточные и тканевые исследования . 175 (4): 499–522. doi : 10.1007/bf00222415 . PMID 830429 . S2CID 25650919 .
- ^ Stebbins & Cohen (1995) с. 69–72
- ^ Jump up to: а беременный Hubáček, J.; Шугеркова, м.; Gvoždík, L. (2019). «Подводное производство звука варьируется не между видами в симпатрических Ньютонах» . ПЕРЕЙ . 7 : E6649. doi : 10.7717/peerj.6649 . PMC 6441559 . PMID 30944780 .
- ^ Биология позвоночных: систематика, таксономия, естественная история и сохранение
- ^ Stebbins & Cohen (1995) с. 76–77
- ^ Jump up to: а беременный Stebbins & Cohen (1995) с. 17–25
- ^ Cogger & Doubt (1998), с. 74–75.
- ^ Kakehashi, Ryosuke; Курабаяши, Ацуши (2021). «Паттерны естественного отбора на генах, кодирующих митохондриальный белок в саламандерах без легких: смягченный очищающий отбор и наличие положительно выбранных кодовых сайтов в семействе Plethodontidae» . Международный журнал геномики . 2021 : 1–12. doi : 10.1155/2021/6671300 . PMC 8053045 . PMID 33928143 .
- ^ Тойс, Даниэль П. (1974). «Респираторные механизмы в водном саламандре, Amphiuma tridactylum». Копея . 1974 (4): 917–920. doi : 10.2307/1442591 . JSTOR 1442591 .
- ^ Rogge, Jessica R.; ВАКЕНТИН, Карен М. (15 ноября 2008 г.). «Внешние жабры и адаптивное поведение эмбрионов облегчают синхронное развитие и вылупление пластичности при дыхательных ограничениях» . Журнал экспериментальной биологии . 211 (22): 3627–3635. doi : 10.1242/jeb.020958 . ISSN 0022-0949 . PMID 18978228 .
- ^ Duellman, William Edward (1994). Биология амфибий . Издательство Университета Джона Хопкинса.
- ^ V., Kardong, Kenneth (2012). Позвоночные: сравнительная анатомия, функция, эволюция . МакГроу-Хилл. ISBN 9780073524238 Полем OCLC 939087630 .
{{cite book}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Залиско, Эдвард Дж. (2015). Сравнительная анатомия позвоночных. Руководство по рассечению лабораторного рассечения . McGraw-Hill Education. ISBN 9780077657055 Полем OCLC 935173274 .
- ^ Jump up to: а беременный в Хайрстон, Нельсон Г. (1949). «Местное распространение и экология саламандров плетодонтид южных аппалачи». Экологические монографии . 19 (1): 47–73. Bibcode : 1949ecom ... 19 ... 47H . doi : 10.2307/1943584 . JSTOR 1943584 .
- ^ «Plethodontidae» . Amphibiaweb. Архивировано с оригинала 22 февраля 2014 года . Получено 9 февраля 2014 года .
- ^ Коппес, Стив (24 сентября 2009 г.). «Он не вкусно, он мой брат» . Спросите биолога . Университет штата Аризона. Архивировано с оригинала 22 февраля 2014 года . Получено 8 февраля 2014 года .
- ^ " Desmognathus Quadramaculatus " . Amphibiaweb. Архивировано с оригинала 22 февраля 2014 года . Получено 9 февраля 2014 года .
- ^ Кардонг (2009), стр. 505–506.
- ^ Stebbins & Cohen (1995) с. 57–58
- ^ Stebbins & Cohen (1995) с. 58–60
- ^ Ларсен, Джон Х. младший; Гатри, Дэн Дж. (1975). «Система кормления наземного тигра саламандров ( Ambystoma tigrinum melanostictum baird)». Журнал морфологии . 147 (2): 137–153. doi : 10.1002/jmor.1051470203 . PMID 30309060 . S2CID 52959566 .
- ^ Дебан, С.М.; Wake, DB; Рот, Г. (1997). «Саламандре с балистическим языком». Природа . 389 (6646): 27–28. Bibcode : 1997natur.389 ... 27d . doi : 10.1038/37898 . S2CID 205026166 .
- ^ Дебан, С.М.; O'Reilly, U. Dicke (2007). «Чрезвычайно мощный проекция языка в саламандерах Plethodontid» . Журнал экспериментальной биологии . 210 (4): 655–667. doi : 10.1242/jeb.02664 . PMID 17267651 .
- ^ Jump up to: а беременный Регал, Филипп Дж. (1966). «Специализации кормления и классификация наземных саламандров». Эволюция 20 (3): 392–407. doi : 10.2307/2406638 . JSTOR 2406638 . PMID 28562974 .
- ^ Хилл, RL; Мендольсон, младший; Sabile, JL (2015). Полем Юго -восточный натуралист 14 : N5 - N9. doi : 10.1656/058.014.0104 . S2CID 86233204 .
- ^ Stebbins & Cohen (1995) с. 110–120
- ^ Мейсон, Дж. Рассел; Рабин, Майкл Д.; Стивенс, Дэвид А. (1982). «Основные отвращения вкуса: выделители кожи, используемые для защиты Тигра Саламандерс, Ambystoma tigrinum ». Копея . 1982 (3): 667–671. doi : 10.2307/1444668 . JSTOR 1444668 .
- ^ Brodie, Edmund D. Jr.; Smatresk, Neal J. (1990). «Антипредатор Арсенал огненного саламандров: распыление секретов из дорсальных кожных желез с высоким давлением». Herpetologica . 46 (1): 1–7. JSTOR 3892595 .
- ^ Heiss, E.; Natchev, N.; Salaberger, D.; Gumpenberger, M.; Рабансер, А.; Weisgram, J. (2010). «Повредят себе, причиняя боль своему врагу: новое понимание функции странного механизма антипредредоватора в саламандридских плевроделях Waltl » . Журнал зоологии . 280 (2): 156–162. doi : 10.1111/j.1469-7998.2009.00631.x .
- ^ Jump up to: а беременный Говард, Ронни Р.; Броди, Эдмунд Д. младший (1973). «Бейтсианский миметический комплекс в саламандре: ответы птичьих хищников». Herpetologica . 29 (1): 33–41. JSTOR 3891196 .
- ^ Кучта, ср; Кракауэр, ах; Sinervo, B (2008). «Почему у Энсатины желтоглазый есть желтые глаза? Бейтсианская мимикрия Pacific Newts (род Trachia ) от саламандры Энсатина Eschscholtzii xanthoptica » . Эволюция 62 (4): 984–990. doi : 10.1111/j.1558-5646.2008.00338.x . PMID 18248632 . S2CID 998486 .
- ^ Котт, 1940. Стр. 204.
- ^ Монаган, Джеймс Р.; Эпп, Леонард Г.; Путта, Шрикришна; Пейдж, Роберт Б.; Уокер, Джон А.; Бични, Крис К.; Чжу, Вэй; Пао, Джеральд М.; Верма, Индер М.; Охотник, Тони; Брайант, Сьюзен v.; Гардинер, Дэвид М.; Харкинс, Тим Т.; Восс, С. Рэндал (2009). «Анализ последовательности микрочипов и кДНК транскрипции во время нервной регенерации конечностей» . BMC Biology . 7 (1): 1. DOI : 10.1186/1741-7007-7-1 . PMC 2630914 . PMID 19144100 .
- ^ Schoch, Rainer R.; Вернебург, Ральф; Voigt, Себастьян (2020). «Триасовый ствол-саламандр из Кыргызстана и происхождение саламандров» . Труды Национальной академии наук . 117 (21): 11584–11588. Bibcode : 2020pnas..11711584S . doi : 10.1073/pnas.2001424117 . PMC 7261083 . PMID 32393623 .
- ^ Эванс, SE; Милнер, Ар; Mussett, F. (1988). «Самые ранние известные саламандры (амфибия, каудата): запись из средней юры Англии». Geobios . 21 (5): 539–552. Bibcode : 1988geobi..21..539e . doi : 10.1016/s0016-6995 (88) 80069-x .
- ^ Jump up to: а беременный Джонс, Марк Эх; Бенсон, Роджер Б.Дж.; Skutschas, Павел; Хилл, Люси; Панчироли, Эльза; Шмитт, Армин Д.; Уолш, Стиг А.; Эванс, Сьюзен Э. (11 июля 2022 года). «Окаменелости среднего юрского периода документируют раннюю стадию в эволюции Саламандры» . Труды Национальной академии наук . 119 (30): E2114100119. Bibcode : 2022pnas..11914100J . doi : 10.1073/pnas.2114100119 . ISSN 0027-8424 . PMC 9335269 . PMID 35858401 .
- ^ Gao, Ke-Qin; Шубин, Нил Х. (2012). «Поздний юрский саламандроид из Western Liaoning, Китай» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (15): 5767–5772. BIBCODE : 2012PNAS..109.5767G . doi : 10.1073/pnas.1009828109 . PMC 3326464 . PMID 22411790 .
- ^ Marjanovic, D.; Лаурин М. (2014). «Обновленная палеонтологическая срока Lissamphipians, с комментариями по анатомии юрских саламандерс (Urodela)». Историческая биология . 26 (4): 535–550. Bibcode : 2014hbio ... 26..535m . doi : 10.1080/08912963.2013.797972 . S2CID 84581331 .
- ^ Shekhovtsov, Sergei V.; Bulakhova, Nina A.; Tsentalovich, Yuri P.; Zelentsova, Ekaterina A.; Meshcheryakova, Ekaterina N.; Poluboyarova, Tatiana V.; Berman, Daniil I. (12 November 2021). "Biochemical Response to Freezing in the Siberian Salamander Salamandrella keyserlingii" . Biology . 10 (11): 1172. doi : 10.3390/biology10111172 . PMC 8614755 . PMID 34827165 .
- ^ Уэллс, Кентвуд Д. (2007). Экология и поведение амфибий . Университет Чикагской Прессы. п. 130. ISBN 978-0-226-89334-1 .
- ^ Элмер, Кр; Бонетт, RM; Wake, DB; Lougheed, SC (4 марта 2013 г.). «Раннее миоценовое происхождение и загадочная диверсификация южноамериканских саламандров» . BMC Эволюционная биология . 13 (1): 59. Bibcode : 2013bmcee..13 ... 59e . doi : 10.1186/1471-2148-13-59 . PMC 3602097 . PMID 23497060 .
- ^ «Первое открытие саламандры в янтаре проливает свет на эволюцию Карибских островов | Новости и исследовательские коммуникации | Орегонский государственный университет» . Oregonstate.edu . 17 августа 2015 года. Архивировано с оригинала 19 февраля 2017 года . Получено 25 марта 2016 года .
- ^ Болес, мы (1997). «Отопаемые певчие птицы (пассирифа) из раннего эоцена Австралии». Эму . 97 (1): 43–50. Bibcode : 1997emuao..97 ... 43b . doi : 10.1071/mu97004 . ISSN 0158-4197 .
- ^ «Вид по номеру» . Amphibiaweb. Архивировано из оригинала 12 января 2021 года . Получено 11 января 2021 года .
- ^ Jump up to: а беременный в дюймовый и Найш, Даррен (1 октября 2013 г.). «Удивительный мир саламандров» . Scientific American . Архивировано из оригинала 16 января 2014 года . Получено 14 января 2014 года .
- ^ Брэд Шаффер; Оскар Флорес-Виллела; Габриэла Парра-Олея; Дэвид Уэйк (2004). «Амбистома Андерсон». МСОП красный список угрожаемых видов. Версия 2013.2. Международный союз сохранения природы
- ^ Брэдли, Дж. Гэвин; Eason, Perri K. (2018). « Eurycea lucifuga (Cave Salamander) вокализация» . Герпетологический обзор . 49 (4): 725.
{{cite journal}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а беременный в Stebbins & Cohen (1995) с. 143–154
- ^ Буэшинг, Кристина Д. (19 июля 2019 г.). Химические сигналы у позвоночных 14 . Спрингер. ISBN 9783030176167 .
- ^ «Изучение влияния систем спаривания на размер яичек в саламандре» . Архивировано из оригинала 16 февраля 2023 года . Получено 16 февраля 2023 года .
- ^ Цзя, Цзя; Андерсон, Джейсон С.; Gao, Ke-Qin (2021). «Средние юры гинобииды из Китая проливают свет на эволюцию базальных саламандров» . ISCAING . 24 (7). Bibcode : 2021isci ... 24J2744J . doi : 10.1016/j.isci.2021.102744 . PMC 8264161 . PMID 34278256 .
- ^ «Эволюционная репродуктивная морфология амфибий» (PDF) . Архивировано (PDF) из оригинала 7 марта 2023 года . Получено 15 февраля 2023 года .
- ^ Buverro, S.; Sotgiu, G.; Кастелано, с.; Джакома, С. (2003). Саламандриды из Сардинии " Копея . 2003 : 149–1 doi : (2003) 003 [0149: Zepia] 10.1643/ 00045-8511 S2CID 8590980 .
- ^ Уэллс, Кентвуд Д. (15 февраля 2010 г.). Экология и поведение амфибий . Университет Чикагской Прессы. ISBN 9780226893334 .
- ^ Ваги, Балазс; Марш, Даниэль; Солдат, Гергели; Végvári, Zsolt; Freckleton, Robert P.; Лайкер, Андрас; Székely, Tamás (5 октября 2022 г.). «Эволюция родительской помощи в саламандре - природа» . Научные отчеты . 12 (1): 16655 doi : 10.1038/s41598-022-20903-3 . PMC 9535019 . PMID 36198742 .
- ^ Репродуктивная биология и филогения уроделы
- ^ Jump up to: а беременный Stebbins & Cohen (1995) с. 175–179
- ^ «Plethodontidae» . Amphibiaweb. Архивировано из оригинала 9 июня 2015 года . Получено 9 января 2014 года .
- ^ Jump up to: а беременный Kiyonaga, Robin R. «Метаморфоза против Неотен (педорфоза) в саламандре (Caudata)» . Архивировано из оригинала 1 декабря 2008 года . Получено 4 мая 2008 года .
- ^ Jump up to: а беременный Сандовал-Комт, Адриана; Пинеда, Эдуардо; Агилар-Лопес, Хосе Л. (2012). «В поисках критических исчезающих видов: нынешняя ситуация двух крошечных видов саламандры в неотропических горах Мексики» . Plos один . 7 (4): E34023. BIBCODE : 2012PLOSO ... 734023S . doi : 10.1371/journal.pone.0034023 . PMC 3317776 . PMID 22485155 .
- ^ Jump up to: а беременный Тапли, Бенджамин; Брэдфилд, Кей С.; Майклс, Кристофер; Бунгард, Майк (24 июля 2015 г.). «Амфибии и программы разведения консервации: все ли угрожающие амфибии принадлежат ковчегу?». Биоразнообразие и сохранение . 24 (11): 2625–2646. Bibcode : 2015bicon..24.2625t . doi : 10.1007/s10531-015-0966-9 . ISSN 0960-3115 . S2CID 11824410 .
- ^ Фонтан, Генри (16 февраля 2009 г.). «Еще одна амфибия в опасности: саламандры» . New York Times . Архивировано с оригинала 26 ноября 2013 года . Получено 28 июня 2013 года .
- ^ "Китайский гигант саламандре" . ZSL сохранение . Зоологическое общество Лондона. Архивировано из оригинала 28 июня 2013 года . Получено 21 июля 2013 года .
- ^ Уилер, Бенджамин А.; Просен, Итан; Матис, Алисия; Уилкинсон, Роберт Ф. (2003). «Снижение населения долгоживущего саламандры: 20+-лежащее исследование« Адскихбендеров », Cryptobranchus Alleganiensis ». Биологическое сохранение . 109 (1): 151–156. Bibcode : 2003bcons.109..151W . doi : 10.1016/s0006-3207 (02) 00136-2 .
- ^ Уилер, Бенджамин А.; McCallum, Malcolm L.; Trauth, Stanley E. (2002). «Аномалии в Озарке Алдбендер ( Ctyptobranchm Alleganiensis Bishopi ) в Арканзасе: сравнение между двумя реками с исторической перспективой» . Журнал Арканзасской академии науки . 58 : 250–252.
- ^ «Первое в мире пленник размножения Озарка Адпендеров» . Наука ежедневно . 1 декабря 2011 года. Архивировано из оригинала 11 мая 2013 года . Получено 21 июля 2013 года .
- ^ «Саламандры без легких Мексики» . Край: эволюционный отличный и глобальный под угрозой исчезновения . Зоологическое общество Лондона. Архивировано из оригинала 29 мая 2013 года . Получено 21 июля 2013 года .
- ^ «Край существования» . Край существования . Архивировано с оригинала 17 ноября 2015 года . Получено 29 октября 2015 года .
- ^ Jump up to: а беременный Контрерас, Виктория; Мартинес-Мейер, Энрике; Ванент, Эльза; Замбрано, Луис (1 декабря 2009 г.). «Недавнее снижение и потенциальное распределение в последней остаточной области микроэндемической мексиканской аксолотля ( Ambystoma Mexicanum )». Биологическое сохранение . 142 (12): 2881–2885. Bibcode : 2009bcons.142.2881c . doi : 10.1016/j.biocon.2009.07.008 .
- ^ Parra-Olea, G.; Zamudio, KR; Recuero, E.; Aguilar-Miguel, x.; Huaacuz, D.; Замбрано Л. (1 февраля 2012 г.). «Сохранение генетики угроз мексиканских аксиканских аксолотлов ( Ambystoma )» животных Сохранение 15 (1): 61–7 Bibcode : 2012ancon..15 ... 61p Doi : 10.1111/j.1469-1795.2011.00488.x . ISSN 1469-1 S2CID 46992721
- ^ Алькарас, Гильмина; Лопес-портала, Ксари; Роблс-Мендоза, Сесилия (1 февраля 2015 г.). «Ответ родного исчезающего аксолотля, Ambysto Mexicanum (амфибия) на экзотический рыбный хищник» Гидробиология 753 (1): 73–8 Doi : 10.1007/ s10750-015-2194-4 ISSN 0018-8 S2CID 17468971
- ^ Фигил, Честер (2013). «Криоконсервация сперматозоидов из аксолотл -амбистомы Мексиканум: значение для сохранения» (PDF) . Герпетологическое сохранение и биология . Архивировал (PDF) из оригинала 22 марта 2016 года . Получено 26 октября 2015 года .
- ^ Марсек, Рут; Бемент, Ханна. «Сохранение саламандры» . Сохранение амфибий . Мемфис зоопарк. Архивировано из оригинала 31 июля 2013 года . Получено 21 июля 2013 года .
- ^ Райс, Эдвард Лоранус (1935). Введение в биологию . Бостон, Массачусетс, США: Джинн и Компания. п. 253.
- ^ Jump up to: а беременный Ларсон, А.; Dimmick, W. (1993). «Филогенетические отношения семейств саламандре: анализ конгруэнтности среди морфологических и молекулярных признаков». Герпетологические монографии . 7 (7): 77–93. doi : 10.2307/1466953 . JSTOR 1466953 .
- ^ Блэкберн, Дэвид С.; Wake, Дэвид Б. (23 декабря 2011 г.). «Класс Амфибия Грей, 1825. В: Zhang, Z.-Q. (ed.) Биоразнообразие животных: план классификации более высокого уровня и обследование таксономического богатства». Zootaxa . 3148 (1). doi : 10.11646/Zootaxa.3148.1.8 .
- ^ Сан -Мауро, Диего; Венс, Майкл; Alcobendas, Марина; Зардоя, Рафаэль; Мейер, Аксель (2005). «Первоначальная диверсификация живых амфибий предшествовала расставанию пангеи» (PDF ) Американский натуралист 165 (5): 590–5 Doi : 10.1086/ 4 PMID 15795855 S2CID 17021360
- ^ Сан -Мауро, Д. (2010). «Многолокусный шкала времени для происхождения существующих амфибий». Молекулярная филогенетика и эволюция . 56 (3): 554–561. Bibcode : 2010molpe..56..554S . doi : 10.1016/j.ympev.2010.04.019 . PMID 20399871 .
- ^ Marjanović D, Laurin M (2007). «Окаменелости, молекулы, время дивергенции и происхождение лиссамбийцев» . Систематическая биология . 56 (3): 369–388. doi : 10.1080/10635150701397635 . PMID 17520502 .
- ^ Schoch, Rainer R.; Вернебург, Ральф; Voigt, Себастьян (26 мая 2020 г.). «Триасовый ствол-саламандр из Кыргызстана и происхождение саламандров» . Труды Национальной академии наук . 117 (21): 11584–11588. Bibcode : 2020pnas..11711584S . doi : 10.1073/pnas.2001424117 . ISSN 0027-8424 . PMC 7261083 . PMID 32393623 .
- ^ Лаурин М (2014). Старый известный известен. Старинная болология 27 (1): 109–1 doi : 10.1080/ 0 S2CID 83557507 .
- ^ Marjanovic D, Laurin M (2014). «Обновленная палеонтологическая срока Lissamphipians, с комментариями по анатомии юрских саламандерс (Urodela)». Историческая биология . 26 (4): 535–550. Bibcode : 2014hbio ... 26..535m . doi : 10.1080/08912963.2013.797972 . S2CID 84581331 .
- ^ Р. Александр Пирон; Джон Дж. Винс (2011). «Большая филогения амфибий, в том числе более 2800 видов, и пересмотренная классификация существующих лягушек, саламандров и кесэцилия» . Молекулярная филогенетика и эволюция . 61 (2): 543–583. Bibcode : 2011molpe..61..543a . doi : 10.1016/j.ympev.2011.06.012 . PMID 21723399 .
- ^ Солнце, Ченг; Шепард, Дональд Б.; Чонг, Ребекка А.; Лопес Арриаса, Хосе; Холл, Кэтрин; Кастоэ, Тодд А.; Feschotte, Cédric; Поллок, Дэвид Д.; Мюллер, Рэйчел Локридж (1 января 2012 г.). «Ретротранспозоны LTR способствуют геномному гигантизму у саламандров Plethodontid» . Биология и эволюция генома . 4 (2): 168–183. doi : 10.1093/gbe/evr139 . PMC 3318908 . PMID 22200636 .
- ^ Элева, Ахмед; Ван, Хенг; Талавера-Лопес, Карлос; Джовен, Альберто; Брито, Гонсало; Карар, Аноп; Хамид, Л. Шахул; Penrad-Mobayed, май; Яо, Зейу (22 декабря 2017 г.). «Чтение и редактирование генома Plerodeles Waltl раскрывает новые особенности регенерации Tettrapod » Природная связь 8 (1): 2286. Bibcode : 2017natco ... 8.2286e Doi : 10.1038/ s41467-017-01964-9 ISSN 2041-1 5741667PMC PMID 29273779
- ^ Nowoshilow, Sergej; Schloissnig, Siegfried; Фэй, Джи-Фенг; Дал, Андреас; Пан, Энди В.К.; Пиппел, Мартин; Винклер, Сильке; Hastie, Alex R.; Молодой, Джордж; Roscito, Juliana G.; Сокол, Франциско; Кнапп, Данджа; Пауэлл, Шон; Круз, Альфредо; Цао, Хан; Хаберманн, Бьянка; Хиллер, Майкл; Танака, Элли М.; Майерс, Юджин В. (2018). «Геном аксолотла и эволюция ключевых регуляторов образования тканей» . Природа . 554 (7690): 50–55. Bibcode : 2018natur.554 ... 50n . doi : 10.1038/nature25458 . HDL : 21.11116/0000-0003-F659-4 . PMID 29364872 .
- ^ Солнце, C.; Мюллер, RL (17 июля 2014 г.). «Последовательности генома Hellbender проливают свет на геномную экспансию у основания Crown Salamanders» . Биология и эволюция генома . 6 (7): 1818–1829. doi : 10.1093/gbe/evu143 . PMC 4122941 . PMID 25115007 .
- ^ Сессии, Стэнли К.; Wake, David B. (июнь 2021 г.). «Навсегда молодой: связывание регенерации и размера генома в саламандре». Динамика развития . 250 (6): 768–778. doi : 10.1002/dvdy.279 . PMID 33320991 .
- ^ Фокс, Дуглас (февраль 2022 г.). «Нежелательная ДНК деформирует тела саламандры» . Scientific American . 326 (2): 40.
- ^ Фокс, Дуглас (29 сентября 2022 г.). «Живые загадки: у этого твара в 38 раз больше ДНК, чем вы» . Science News исследует .
- ^ Преимущества «зомби» клеток: стареющие клетки помогают регенерации в саламандерах
- ^ Эшкрофт, Фрэнсис (2002). Жизнь в крайности: наука о выживании . Калифорнийский университет. п. 112 . ISBN 978-0-520-22234-2 .
- ^ Плиний старший (ок. 100). Естественная история . С. Книга 10, 86. Архивировано из оригинала 18 октября 2008 года . Получено 8 января 2014 года .
- ^ Святой Августин (ок. 500). Город Божий . С. Книга 21, 4.
- ^ Исидор из Севилья (ок. 700). Этимологии . С. Книга 12, 4:36. Архивировано из оригинала 18 октября 2008 года . Получено 8 января 2014 года .
- ^ Браун, Клэр (2003). «Шерсть Саламандры: исторические доказательства текстиля, сотканного с асбестом». История текстиля . 34 : 64–73. doi : 10.1179/004049603235001454 . S2CID 191599472 .
- ^ Jump up to: а беременный Уайт, Т. Т. (1992) [1954]. Книга зверей: быть переводом из латинского бестиария двенадцатого века . Алан Саттон . С. 183–184. ISBN 978-0-7509-0206-9 .
- ^ Cellini, Benvenuto (1998). Автобиография Бенвенуто Каллини . Англия: книги пингвинов. п. 34. ISBN 0140447180 .
- ^ Директор: Дункан Чард (14 февраля 2012 г.). «Речные монстры». Программа 6, серия 3 . Itv . Itv1 .
- ^ Голландия, Николас (2021), «Мемуары Висензо Колуччи 1886 года, вокруг регенерации конечностей и хвоста в Тритонах, аннотируют и переведены на английский как: в отношении регенерации конечностей и хвоста в саламандре», европейский журнал , 88 : 837 –890, doi : 10.1080/24750263.2021.1943549
- ^ Кейм, Брэндон (1 июля 2009 г.). «Открытие саламандры может привести к возрождению человеческой конечности» . Проводной . Получено 7 мая 2010 года .
- ^ «Иммунная система саламандров держат ключ к регенерации?» Полем Scienceday . Архивировано из оригинала 6 декабря 2013 года . Получено 21 мая 2013 года .
- ^ Симбиотические отношения саламандры/водоросли могут вдохновить новые лекарства
- ^ Голдсмит, Ричард (11 июня 2010 г.). «Обозначающие галлюциногенные напитки» . Fox News . Архивировано из оригинала 5 декабря 2014 года . Получено 2 января 2014 года .
- ^ Козорог, Миха (2003). «Саламандре Бренди:« психоделический напиток »между медиа -мифом и практикой домашнего алкоголя в Словении» . Антропология Восточной Европы обзор . 21 (1): 63–71. Архивировано из оригинала 31 марта 2014 года . Получено 22 марта 2014 года .
Цитируемые тексты
[ редактировать ]- Cogger, Hg; Zweifel, RG , Eds. (1998). Энциклопедия рептилий и амфибий (2 -е изд.). Академическая пресса. ISBN 978-0-12-178560-4 .
- Котт, Хью Б. (1940). Адаптивная окраска у животных . Издательство Оксфордского университета.
- Дорит, RL; Уокер, WF; Barnes, RD (1991). Зоология . Saunders College Publishing. ISBN 978-0-03-030504-7 .
- Кардонг, Кеннет В. (2009). Позвоночные: сравнительная анатомия, функция, эволюция (5 -е изд.). МакГроу-Хилл. ISBN 978-0-07-304058-5 .
- Стеббинс, Роберт С .; Коэн, Натан В. (1995). Естественная история амфибий . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. ISBN 978-0-691-03281-8 .
Внешние ссылки
[ редактировать ]- Дерево жизни: Каудата
- Галерея саламандры
- Caudata Culture
- Крайтерные перекрестки: саламандры туннели в Министерстве транспорта США
- Archéozooteque: Уроделе скелетоны рисунок : доступно в форматах вектора, изображения и PDF




Хвостовые семьи по подройке |
|---|

| Базы данных управления авторитетом : национальный |
|---|