ДНК (цитозин-5)-метилтрансфераза 3А
| ДНМТ3А | |||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | DNMT3A , DNMT3A2, M.HsaIIIA, TBRS, ДНК (цитозин-5-)-метилтрансфераза 3 альфа, ДНК-метилтрансфераза 3 альфа, HESJAS | ||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | ОМИМ : 602769 ; МГИ : 1261827 ; Гомогены : 7294 ; Генные карты : DNMT3A ; ОМА : DNMT3A — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||





ДНК (цитозин-5)-метилтрансфераза 3А ( DNMT3A ) представляет собой фермент , который катализирует перенос метильных групп к определенным структурам CpG в ДНК, процесс, называемый метилированием ДНК . У человека фермент кодируется DNMT3A геном . [5] [6]
Этот фермент отвечает за метилирование ДНК de novo . Такую функцию следует отличать от поддерживающего метилирования ДНК, которое обеспечивает точность репликации наследственных эпигенетических паттернов. DNMT3A является частью семейства ферментов ДНК-метилтрансфераз , которое состоит из главных героев DNMT1 , DNMT3A и DNMT3B . [5] [6]
Хотя метилирование ДНК de novo изменяет информацию, передаваемую родителем потомству, оно обеспечивает ключевые эпигенетические модификации, необходимые для таких процессов, как клеточная дифференциация и эмбриональное развитие , регуляция транскрипции , образование гетерохроматина , Х-инактивация , импринтинг и стабильность генома. [7]
DNMT3a — это ген, который чаще всего обнаруживается мутировавшим при клональном гемопоэзе , распространенном феномене, связанном со старением, при котором гемопоэтические стволовые клетки (HSC) или другие ранние предшественники клеток крови способствуют формированию генетически отличной субпопуляции клеток крови . [8] [9] [10]
Ген
[ редактировать ]DNMT3A представляет собой белок массой 130 кДа, кодируемый 23 экзонами , обнаруженными на хромосоме 2p23 у человека. [11] Существует 98% гомология между человеческими и мышиными гомологами. [6] DNMT3A широко экспрессируется среди млекопитающих. [12]
Существует две основные изоформы белка: DNMT3A1 и DNMT3A2 с молекулярной массой около 130 кДа и 100 кДа соответственно. Белок DNMT3A2, у которого отсутствует N-концевая область DNMT3A1, кодируется транскриптом, инициируемым нижестоящим промотором. [13] Эти изоформы существуют в разных типах клеток. [14] При первоначальном создании [13] Было обнаружено, что DNMT3A2 высоко экспрессируется в семенниках, яичниках, селезенке и тимусе. Совсем недавно было показано, что он индуцируемо экспрессируется в гиппокампе головного мозга. [15] и необходим в гиппокампе при формировании памяти. [16] DNMT3A2 также активируется в оболочке прилежащего ядра в ответ на кокаин . [17]
Структура белка
[ редактировать ]DNMT3A состоит из трех основных белковых доменов: домена Pro-Trp-Trp-Pro (PWWP), домена ATRX-DNMT3-DNMT3L (ADD) и домена каталитической метилтрансферазы.

Структуры DNMT3A1 и DNMT3A2 имеют аналогию со структурой DNMT3B1, а также с двумя вспомогательными белками DNMT3B3 и DNMT3L (см. рисунок упрощенных доменов изоформ DNMT3A). Два вспомогательных белка стимулируют метилирование de novo посредством каждого из своих взаимодействий с тремя изоформами, имеющими функциональный каталитический домен. В общем, всем DNMT для выполнения своей биологической функции требуются вспомогательные белки. [18]
Мотив PWWP находится в домене, состоящем примерно из 100 аминокислот, который имеет одну область со значительным количеством основных остатков (лизинов и аргининов), образующих положительно заряженную поверхность, которая может связываться с ДНК. Отдельная область домена PWWP может связываться с метил-лизинами гистонов через гидрофобный карман, который включает сам мотив PWWP. [19] [20]
Домен ADD DNMT3A состоит из N-концевого GATA-подобного цинкового пальца , пальца PHD и C-концевой альфа-спирали , которые вместе образуют единую глобулярную складку. Этот домен может действовать как считыватель, который специфически связывается с гистоном H3, неметилированным по лизину 4 (H3K4me0). [21] Домен ADD служит ингибитором домена метилтрансферазы до тех пор, пока DNMT3A не свяжется с немодифицированным лизином 4 гистона 3 (H3K4me0) с его метилирующей активностью de novo . [14] Таким образом, DNMT3A, по-видимому, имеет встроенный механизм контроля, направленный на метилирование ДНК только в гистонах, которые неметилированы по гистону 3, при этом лизин в 4-м положении от аминоконца неметилирован.
Каталитический домен (домен метилтрансферазы) высоко консервативен даже среди прокариот . [22]

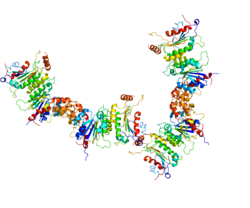
Три ДНК-метилтрансферазы (DNMT3A1, DNMT3A2 и DNMT3B) катализируют реакции размещения метильной группы на цитозине, обычно в CpG-сайте ДНК. [23] На сопроводительном рисунке показан комплекс метилтрансферазы, содержащий DNMT3A2. Чтобы быть эффективными, эти ферменты должны действовать совместно с дополнительным белком (например, DNMT3B3, DNMT3L или другими). [24] [25] [26] Два вспомогательных белка (не обладающих каталитической активностью), образующие комплекс с двумя DNMT с каталитическим доменом, встречаются в виде гетеротетрамера (см. Рисунок). Эти гетеротетрамеры встречаются в следующем порядке: дополнительный белок-каталитический белок-каталитический белок-вспомогательный белок. Конкретный комплекс, показанный на рисунке, иллюстрирует гетеротетрамер, образованный каталитическим белком DNMT3A2 и дополнительным белком DNMT3B3. Один вспомогательный белок комплекса связывается с кислым участком ядра нуклеосомы (см. верхнюю часть 3B3 на рисунке). Соединение одного дополнительного белка с нуклеосомой ориентирует гетеротетрамер. Такая ориентация помещает первый каталитический DNMT (ближайший к вспомогательному белку, связанному с нуклеосомой) в промежуточное положение (не близко к линкерной ДНК). Второй каталитический DNMT (нижний 3A2 на рисунке) расположен у линкерной ДНК. Метилирование может происходить внутри этой линкерной ДНК (как показано на рисунке), но не в какой-либо ДНК, обернутой вокруг ядра нуклеосомы.
Как показали Манзо и др., [27] существуют как специфические индивидуальные сайты связывания трех каталитических DNMT (3A1, 3A2 и 3B3), так и перекрывающиеся сайты связывания этих ферментов. насчитывается 28 миллионов сайтов CpG . В геноме человека [28] Многие из этих CpG расположены внутри CpG-островков (областей ДНК) с относительно высокой плотностью CpG-сайтов. [28] Из этих областей 3970 областей обогащены исключительно DNMT3A1, 3838 областей DNMT3A2 и 3432 области DNMT3B, а также есть сайты, которые являются общими для белков DNMT de novo. [27] Кроме того, то, действует ли ДНК-метилтрансфераза (DNMT3A1, DNMT3A2 или DNMT3B) на доступный сайт CpG, зависит от последовательности, фланкирующей сайт CpG в линкерной ДНК. [26]
Функция
[ редактировать ]DNMT1 отвечает за поддержание метилирования ДНК, тогда как DNMT3A и DNMT3B выполняют как поддержание (исправление ошибок DNMT1), так и метилирование ДНК de novo. Было обнаружено, что после нокаута DNMT1 в раковых клетках человека эти клетки сохраняют унаследованный характер метилирования. [29] что предполагает поддерживающую активность экспрессируемых DNMT3. DNMT3 проявляют одинаковое сродство к неметилированным и гемиметилированным субстратам ДНК. [29] в то время как DNMT1 в 10-40 раз предпочитает гемиметилированную ДНК. [30] [31] DNMT3 могут связываться с обеими формами и, следовательно, потенциально осуществлять как поддержание, так и модификации de novo.
Метилирование de novo является основной признанной активностью DNMT3A, которая необходима для процессов, подобных тем, которые упомянуты во вводных параграфах. Генетический импринтинг предотвращает партеногенез у млекопитающих. [32] и, следовательно, усиливает половое размножение и его многочисленные последствия для генетики и филогенеза. DNMT3A необходим для генетического импринтинга. [33]
Исследования долговременной памяти у людей показывают, что память поддерживается за счет метилирования ДНК . [34] Крысы, у которых новая, сильная долговременная память индуцируется из-за контекстуального обусловливания страха, снизили экспрессию около 1000 генов и увеличили экспрессию около 500 генов в области гиппокампа мозга. Эти изменения происходят через 24 часа после тренировки. На данный момент наблюдается модифицированная экспрессия 9,17% генома гиппокампа крысы. Снижение экспрессии генов связано с de novo . метилированием генов [35]
Исследования на животных
[ редактировать ]У мышей было показано, что снижение экспрессии этого гена у стареющих животных приводит к ухудшению когнитивной долговременной памяти. [15]
У мышей Dnmt3a-/- экспрессия многих генов, связанных с самообновлением HSC, увеличивается, а некоторые не могут быть должным образом репрессированы во время дифференцировки. [36] Это предполагает отмену дифференцировки гемопоэтических стволовых клеток (ГСК) и вместо этого увеличение самообновляющегося клеточного деления. Действительно, было обнаружено, что дифференцировка частично восстанавливалась, если HSC Dnmt3a-/- подвергались дополнительному нокдауну Ctnb1 – Ctnb1 кодирует β-катенин, который участвует в клеточном делении самообновления. [14]
Клиническая значимость
[ редактировать ]Этот ген часто мутирует при раке и является одним из 127 часто мутирующих генов, выявленных в «Атлас генома рака» . проекте [37] Мутации DNMT3A чаще всего наблюдались при остром миелоидном лейкозе (ОМЛ), где они встречались чуть более чем в 25% секвенированных случаев. Эти мутации чаще всего возникают в положении R882 белка, и эта мутация может привести к потере функции. [38] Мутации DNMT3A связаны с плохой общей выживаемостью, что позволяет предположить, что они оказывают важное общее влияние на способность клеток ОМЛ вызывать летальное заболевание. [39] Также было обнаружено, что DNMT3A клеточные линии с мутацией демонстрируют нестабильность транскриптома , поскольку они имеют гораздо более ошибочный сплайсинг РНК по сравнению с их изогенными аналогами дикого типа. [40] Мутации в этом гене также связаны с синдромом Таттона-Брауна-Рахмана , расстройством избыточного роста.
Взаимодействия
[ редактировать ]Было показано, что DNMT3A взаимодействует с:
Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Версия Ensembl 89: ENSG00000119772 – Ensembl , май 2017 г.
- ^ Jump up to: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000020661 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Jump up to: а б Окано М., Се С., Ли Э (июль 1998 г.). «Клонирование и характеристика семейства новых метилтрансфераз ДНК (цитозин-5) млекопитающих». Природная генетика . 19 (3): 219–220. дои : 10.1038/890 . ПМИД 9662389 . S2CID 256263 .
- ^ Jump up to: а б с Се С., Ван З., Окано М., Ногами М., Ли Ю., Хэ WW и др. (август 1999 г.). «Клонирование, экспрессия и расположение хромосом семейства генов DNMT3 человека». Джин . 236 (1): 87–95. дои : 10.1016/S0378-1119(99)00252-8 . ПМИД 10433969 .
- ^ Цзя Ю, Ли П, Фанг Л, Чжу Х, Сюй Л, Ченг Х и др. (12 апреля 2016 г.). «Негативная регуляция метилирования ДНК de novo DNMT3A с помощью часто сверхэкспрессируемых белков семейства UHRF как механизм широко распространенного гипометилирования ДНК при раке» . Открытие клеток . 2 : 16007. дои : 10.1038/celldisc.2016.7 . ПМЦ 4849474 . ПМИД 27462454 .
- ^ Ян М., Эберт Б.Л., Джайсвал С. (январь 2017 г.). «Клональное кроветворение» . Семинары по гематологии . 54 (1): 43–50. doi : 10.1053/j.seminhematol.2016.10.002 . ПМЦ 8045769 . ПМИД 28088988 .
- ^ Сперлинг А.С., Гибсон С.Дж., Эберт Б.Л. (январь 2017 г.). «Генетика миелодиспластического синдрома: от клонального кроветворения к вторичному лейкозу» . Обзоры природы. Рак . 17 (1): 5–19. дои : 10.1038/nrc.2016.112 . ПМК 5470392 . ПМИД 27834397 .
- ^ Стинсма Д.П., Бежар Р., Джайсвал С., Линдсли Р.К., Секерес М.А., Хассерджян Р.П., Эберт Б.Л. (июль 2015 г.). «Клональный кроветворение неопределенного потенциала и его отличие от миелодиспластических синдромов» . Кровь . 126 (1): 9–16. дои : 10.1182/blood-2015-03-631747 . ПМЦ 4624443 . ПМИД 25931582 .
- ^ Робертсон К.Д., Узволги Э., Лян Г., Талмадж С., Сумеги Дж., Гонсалес Ф.А., Джонс П.А. (июнь 1999 г.). «ДНК-метилтрансферазы человека (DNMT) 1, 3a и 3b: координируют экспрессию мРНК в нормальных тканях и сверхэкспрессию в опухолях» . Исследования нуклеиновых кислот . 27 (11): 2291–2298. дои : 10.1093/нар/27.11.2291 . ПМЦ 148793 . ПМИД 10325416 .
- ^ Дель Кастильо Фалькони В.М., Торрес-Арсига К., Матус-Ортега Г., Диас-Чавес Х., Эррера Л.А. (август 2022 г.). «ДНК-метилтрансферазы: от эволюции к клиническому применению» . Международный журнал молекулярных наук . 23 (16): 8994. doi : 10.3390/ijms23168994 . ПМЦ 9409253 . ПМИД 36012258 .
- ^ Jump up to: а б Чен Т, Уэда Ю, Се С, Ли Э (октябрь 2002 г.). «Новая изоформа Dnmt3a, полученная с помощью альтернативного промотора, локализуется в эухроматине, и ее экспрессия коррелирует с активным метилированием de novo» . Журнал биологической химии . 277 (41): 38746–38754. дои : 10.1074/jbc.M205312200 . ПМИД 12138111 .
- ^ Jump up to: а б с Ян Л., Рау Р., Гуделл М.А. (март 2015 г.). «DNMT3A при гематологических злокачественных новообразованиях» . Обзоры природы. Рак . 15 (3): 152–165. дои : 10.1038/nrc3895 . ПМЦ 5814392 . ПМИД 25693834 .
- ^ Jump up to: а б Оливейра А.М., Хемстедт Т.Дж., Бадинг Х. (июль 2012 г.). «Спасение связанного со старением снижения экспрессии Dnmt3a2 восстанавливает когнитивные способности». Природная неврология . 15 (8): 1111–1113. дои : 10.1038/nn.3151 . ПМИД 22751036 . S2CID 10590208 .
- ^ Оливейра А.М., Хемштедт Т.Дж., Фрайтаг Х.Э., Бадинг Х. (август 2016 г.). «Dnmt3a2: хаб для улучшения когнитивных функций» . Молекулярная психиатрия . 21 (8): 1130–1136. дои : 10.1038/mp.2015.175 . ПМИД 26598069 . S2CID 25308306 .
- ^ Каннелла Н., Оливейра А.М., Хемстедт Т., Лиссек Т., Бюхлер Э., Бадинг Х., Спанагель Р. (август 2018 г.). «Dnmt3a2 в оболочке прилежащего ядра необходим для восстановления поиска кокаина» . Журнал неврологии . 38 (34): 7516–7528. doi : 10.1523/JNEUROSCI.0600-18.2018 . ПМК 6596133 . ПМИД 30030395 .
- ^ Гуджар Х, диджей Вайзенбергер, Лян Джи (февраль 2019 г.). «Роль ДНК-метилтрансфераз человека и их изоформ в формировании эпигенома» . Гены . 10 (2): 172. doi : 10.3390/genes10020172 . ПМК 6409524 . ПМИД 30813436 .
- ^ ван Нуланд Р., ван Шайк Ф.М., Симонис М., ван Хиш С., Куппен Э., Боленс Р. и др. (май 2013 г.). «Связывание нуклеосомной ДНК управляет распознаванием H3K36-метилированных нуклеосом доменом PSIP1-PWWP» . Эпигенетика и хроматин . 6 (1): 12. дои : 10.1186/1756-8935-6-12 . ПМЦ 3663649 . ПМИД 23656834 .
- ^ Рона ГБ, Элеутерио ЕС, Пиньейру А.С. (март 2016 г.). «Домены PWWP и их способы восприятия ДНК и гистон-метилированных лизинов» . Биофизические обзоры . 8 (1): 63–74. дои : 10.1007/s12551-015-0190-6 . ПМЦ 5425739 . ПМИД 28510146 .
- ^ Рен В., Гао Л., Сон Дж. (декабрь 2018 г.). «Структурная основа метилирования ДНК, опосредованного DNMT1 и DNMT3A» . Гены . 9 (12): 620. doi : 10.3390/genes9120620 . ПМК 6316889 . ПМИД 30544982 .
- ^ Сюй Ф, Мао С, Дин Ю, Жуй С, Ву Л, Ши А и др. (01.01.2010). «Молекулярные и ферментативные профили ДНК-метилтрансфераз млекопитающих: структуры и мишени для лекарственных средств» . Современная медицинская химия . 17 (33): 4052–4071. дои : 10.2174/092986710793205372 . ПМЦ 3003592 . ПМИД 20939822 .
- ^ Цзэн Ю, Рен Р, Каур Г, Хардикар С, Инь З, Бэбкок Л, Гупта Э, Чжан Икс, Чен Т, Ченг Икс (ноябрь 2020 г.). «Неактивная изоформа Dnmt3b3 преимущественно усиливает Dnmt3b-опосредованное метилирование ДНК» . Генс Дев . 34 (21–22): 1546–1558. дои : 10.1101/gad.341925.120 . ПМЦ 7608744 . ПМИД 33004415 .
- ^ Цзя Д., Юрковска Р.З., Чжан X, Елч А., Ченг X (сентябрь 2007 г.). «Структура Dnmt3a, связанного с Dnmt3L, предполагает модель метилирования ДНК de novo» . Природа . 449 (7159): 248–51. Бибкод : 2007Natur.449..248J . дои : 10.1038/nature06146 . ПМК 2712830 . ПМИД 17713477 .
- ^ Сюй Т.Х., Лю М., Чжоу С.Е., Лян Г., Чжао Г., Сюй Хе, Мельчер К., Джонс П.А. (октябрь 2020 г.). «Структура нуклеосомосвязанных ДНК-метилтрансфераз DNMT3A и DNMT3B» . Природа . 586 (7827): 151–155. Бибкод : 2020Natur.586..151X . дои : 10.1038/s41586-020-2747-1 . ПМЦ 7540737 . ПМИД 32968275 .
- ^ Jump up to: а б Гао Л, Эмперле М, Го Й, Гримм С.А., Рен В, Адам С, Урю Х, Чжан ЗМ, Чен Д, Инь Дж, Дукац М, Антене Х, Юрковска РЗ, Лу Дж, Ван Й, Баштрыков П, Уэйд П.А. , Ван Г.Г., Йельч А., Сон Дж (июль 2020 г.). «Комплексная структурно-функциональная характеристика DNMT3B и DNMT3A выявляет отличительные механизмы метилирования ДНК de novo» . Нат Коммун 11 (1): 3355. Бибкод : 2020NatCo..11.3355G . дои : 10.1038/ s41467-020-17109-4 ПМЦ 7335073 . ПМИД 32620778 .
- ^ Jump up to: а б Манзо М., Вирц Дж., Амбрози С., Вилласеньор Р., Рошицкий Б., Баубек Т. (декабрь 2017 г.). «Изоформ-специфическая локализация DNMT3A регулирует точность метилирования ДНК на двухвалентных CpG-островках» . ЭМБО Дж . 36 (23): 3421–3434. дои : 10.15252/embj.201797038 . ПМЦ 5709737 . ПМИД 29074627 .
- ^ Jump up to: а б Левквист С., Додд И.Б., Снеппен К., Хаертер Дж.О. (июнь 2016 г.). «Метилирование ДНК в эпигеномах человека зависит от локальной топологии сайтов CpG» . Нуклеиновые кислоты Рез . 44 (11): 5123–32. дои : 10.1093/нар/gkw124 . ПМЦ 4914085 . ПМИД 26932361 .
- ^ Jump up to: а б Ри И., Джаир К.В., Йен Р.В., Ленгауэр С., Герман Дж.Г., Кинцлер К.В. и др. (апрель 2000 г.). «Метилирование CpG сохраняется в раковых клетках человека, лишенных DNMT1». Природа . 404 (6781): 1003–1007. Бибкод : 2000Natur.404.1003R . дои : 10.1038/35010000 . ПМИД 10801130 . S2CID 4425037 .
- ^ Прадхан С., Баколла А., Уэллс Р.Д., Робертс Р.Дж. (ноябрь 1999 г.). «Рекомбинантная человеческая ДНК (цитозин-5) метилтрансфераза. I. Экспрессия, очистка и сравнение de novo и поддерживающего метилирования» . Журнал биологической химии . 274 (46): 33002–33010. дои : 10.1074/jbc.274.46.33002 . ПМИД 10551868 .
- ^ Прадхан С., Талбот Д., Ша М., Беннер Дж., Хорнстра Л., Ли Э. и др. (ноябрь 1997 г.). «Бакуловирус-опосредованная экспрессия и характеристика полноразмерной мышиной ДНК-метилтрансферазы» . Исследования нуклеиновых кислот . 25 (22): 4666–4673. дои : 10.1093/нар/25.22.4666 . ПМК 147102 . ПМИД 9358180 .
- ^ Рейк В., Уолтер Дж. (январь 2001 г.). «Геномный импринтинг: родительское влияние на геном». Обзоры природы. Генетика . 2 (1): 21–32. дои : 10.1038/35047554 . ПМИД 11253064 . S2CID 12050251 .
- ^ Канеда М., Окано М., Хата К., Садо Т., Цудзимото Н., Ли Э., Сасаки Х. (июнь 2004 г.). «Основная роль ДНК-метилтрансферазы de novo Dnmt3a в отцовском и материнском импринтинге». Природа . 429 (6994): 900–903. Бибкод : 2004Natur.429..900K . дои : 10.1038/nature02633 . ПМИД 15215868 . S2CID 4344982 .
- ^ Миллер Калифорния, Суэтт Дж. Д. (март 2007 г.). «Ковалентная модификация ДНК регулирует формирование памяти» . Нейрон . 53 (6): 857–869. дои : 10.1016/j.neuron.2007.02.022 . ПМИД 17359920 .
- ^ Дюк К.Г., Кеннеди А.Дж., Гэвин К.Ф., Дэй Дж.Дж., Суэтт Дж.Д. (июль 2017 г.). «Эпигеномная реорганизация в гиппокампе, зависящая от опыта» . Изучите Мем . 24 (7): 278–288. дои : 10.1101/lm.045112.117 . ПМК 5473107 . ПМИД 28620075 .
- ^ Челлен Г.А., Сунь Д., Чон М., Луо М., Джелинек Дж., Берг Дж.С. и др. (декабрь 2011 г.). «Dnmt3a необходим для дифференцировки гемопоэтических стволовых клеток» . Природная генетика . 44 (1): 23–31. дои : 10.1038/ng.1009 . ПМЦ 3637952 . ПМИД 22138693 .
- ^ Кандот С., Маклеллан, доктор медицинских наук, Вандин Ф., Йе К., Ниу Б., Лу С. и др. (октябрь 2013 г.). «Мутационный ландшафт и значение 12 основных типов рака» . Природа . 502 (7471): 333–339. Бибкод : 2013Natur.502..333K . дои : 10.1038/nature12634 . ПМЦ 3927368 . ПМИД 24132290 .
- ^ Ши А.Х., Абдель-Вахаб О., Патель Дж.П., Левин Р.Л. (сентябрь 2012 г.). «Роль мутаций в эпигенетических регуляторах при миелоидных новообразованиях». Обзоры природы. Рак . 12 (9): 599–612. дои : 10.1038/nrc3343 . ПМИД 22898539 . S2CID 20214444 .
- ^ Лей Т.Дж., Дин Л., Уолтер М.Дж., Маклеллан М.Д., Лампрехт Т., Ларсон Д.Э. и др. (декабрь 2010 г.). «Мутации DNMT3A при остром миелолейкозе» . Медицинский журнал Новой Англии . 363 (25): 2424–2433. дои : 10.1056/NEJMoa1005143 . ПМК 3201818 . ПМИД 21067377 .
- ^ Банасзак Л.Г., Джудис В., Чжао X, Ву З, Гао С., Хосокава К. и др. (март 2018 г.). «Аномальный сплайсинг РНК и нестабильность генома после индукции мутаций DNMT3A путем редактирования гена CRISPR/Cas9» . Клетки крови, молекулы и болезни . 69 : 10–22. дои : 10.1016/j.bcmd.2017.12.002 . ПМК 6728079 . ПМИД 29324392 .
- ^ Jump up to: а б Ким Г.Д., Ни Дж., Келесоглу Н., Робертс Р.Дж., Прадхан С. (август 2002 г.). «Сотрудничество и связь между содержанием человека и метилтрансферазами ДНК (цитозин-5) de novo» . Журнал ЭМБО . 21 (15): 4183–4195. дои : 10.1093/emboj/cdf401 . ПМК 126147 . ПМИД 12145218 .
- ^ Jump up to: а б с д Линг Ю., Санкпал Ю.Т., Робертсон А.К., МакНелли Дж.Г., Карпова Т., Робертсон К.Д. (2004). «Модификация ДНК-метилтрансферазы 3a (Dnmt3a) de novo с помощью SUMO-1 модулирует ее взаимодействие с деацетилазами гистонов (HDAC) и ее способность подавлять транскрипцию» . Исследования нуклеиновых кислот . 32 (2): 598–610. дои : 10.1093/nar/gkh195 . ПМЦ 373322 . ПМИД 14752048 .
- ^ Ленертц Б., Уеда Ю., Дерийк А.А., Брауншвейг Ю., Перес-Бургос Л., Кубичек С. и др. (июль 2003 г.). «Метилирование лизина 9 гистона H3, опосредованное Suv39h, направляет метилирование ДНК к основным сателлитным повторам в перицентрическом гетерохроматине» . Современная биология . 13 (14): 1192–1200. Бибкод : 2003CBio...13.1192L . дои : 10.1016/s0960-9822(03)00432-9 . ПМИД 12867029 . S2CID 2320997 .
- ^ Jump up to: а б Фукс Ф., Бургерс В.А., Годин Н., Касаи М., Кузаридес Т. (май 2001 г.). «Dnmt3a связывает деацетилазы и рекрутируется с помощью специфичного для последовательности репрессора для подавления транскрипции» . Журнал ЭМБО . 20 (10): 2536–2544. дои : 10.1093/emboj/20.10.2536 . ПМК 125250 . ПМИД 11350943 .
- ^ Бреннер С., Деплюс Р., Дидело С., Лорио А., Вире Е., Де Смет С. и др. (январь 2005 г.). «Myc подавляет транскрипцию посредством рекрутирования корепрессора ДНК-метилтрансферазы» . Журнал ЭМБО . 24 (2): 336–346. дои : 10.1038/sj.emboj.7600509 . ПМЦ 545804 . ПМИД 15616584 .
- ^ Фукс Ф., Херд П.Дж., Деплюс Р., Кузаридес Т. (май 2003 г.). «ДНК-метилтрансферазы связываются с HP1 и гистон-метилтрансферазой SUV39H1» . Исследования нуклеиновых кислот . 31 (9): 2305–2312. дои : 10.1093/нар/gkg332 . ПМК 154218 . ПМИД 12711675 .
Дальнейшее чтение
[ редактировать ]- Адамс М.Д., Керлаваж А.Р., Флейшманн Р.Д., Фулднер Р.А., Балт С.Дж., Ли Н.Х. и др. (сентябрь 1995 г.). «Первоначальная оценка разнообразия и моделей экспрессии генов человека на основе 83 миллионов нуклеотидов последовательности кДНК». Природа . 377 (6547 Приложение): 3–174. ПМИД 7566098 .
- Бональдо М.Ф., Леннон Дж., Соарес М.Б. (сентябрь 1996 г.). «Нормализация и вычитание: два подхода к открытию генов» . Геномные исследования . 6 (9): 791–806. дои : 10.1101/гр.6.9.791 . ПМИД 8889548 .
- Робертсон К.Д., Узволги Э., Лян Г., Талмадж С., Сумеги Дж., Гонсалес Ф.А., Джонс П.А. (июнь 1999 г.). «ДНК-метилтрансферазы человека (DNMT) 1, 3a и 3b: координируют экспрессию мРНК в нормальных тканях и сверхэкспрессию в опухолях» . Исследования нуклеиновых кислот . 27 (11): 2291–2298. дои : 10.1093/нар/27.11.2291 . ПМЦ 148793 . ПМИД 10325416 .
- Фукс Ф., Бургерс В.А., Годин Н., Касаи М., Кузаридес Т. (май 2001 г.). «Dnmt3a связывает деацетилазы и рекрутируется с помощью специфичного для последовательности репрессора для подавления транскрипции» . Журнал ЭМБО . 20 (10): 2536–2544. дои : 10.1093/emboj/20.10.2536 . ПМК 125250 . ПМИД 11350943 .
- Воте О, Николя Э, Вандель Л, Труш Д (январь 2002 г.). «Функциональное и физическое взаимодействие между метилтрансферазой гистонов Suv39H1 и деацетилазами гистонов» . Исследования нуклеиновых кислот . 30 (2): 475–481. дои : 10.1093/нар/30.2.475 . ПМК 99834 . ПМИД 11788710 .
- Ди Кроче Л., Рейкер В.А., Корсаро М., Фази Ф., Фанелли М., Фаретта М. и др. (февраль 2002 г.). «Привлечение метилтрансферазы и гиперметилирование ДНК целевых промоторов онкогенным фактором транскрипции». Наука . 295 (5557): 1079–1082. Бибкод : 2002Sci...295.1079D . дои : 10.1126/science.1065173 . hdl : 11576/2506625 . ПМИД 11834837 . S2CID 29532358 .
- Хата К., Окано М., Лей Х., Ли Э (апрель 2002 г.). «Dnmt3L сотрудничает с семейством Dnmt3 ДНК-метилтрансфераз de novo для установления материнских отпечатков у мышей». Разработка . 129 (8): 1983–1993. дои : 10.1242/dev.129.8.1983 . ПМИД 11934864 .
- Чен Т, Уэда Ю, Се С, Ли Э (октябрь 2002 г.). «Новая изоформа Dnmt3a, полученная с помощью альтернативного промотора, локализуется в эухроматине, и ее экспрессия коррелирует с активным метилированием de novo» . Журнал биологической химии . 277 (41): 38746–38754. дои : 10.1074/jbc.M205312200 . ПМИД 12138111 .
- Ким Г.Д., Ни Дж., Келесоглу Н., Робертс Р.Дж., Прадхан С. (август 2002 г.). «Сотрудничество и связь между содержанием человека и метилтрансферазами ДНК (цитозин-5) de novo» . Журнал ЭМБО . 21 (15): 4183–4195. дои : 10.1093/emboj/cdf401 . ПМК 126147 . ПМИД 12145218 .
- Янагисава Ю., Ито Э., Юаса Ю., Маруяма К. (сентябрь 2002 г.). «ДНК-метилтрансферазы человека DNMT3A и DNMT3B имеют два типа промоторов с различным содержанием CpG». Biochimica et Biophysical Acta (BBA) – Структура и экспрессия генов . 1577 (3): 457–465. дои : 10.1016/S0167-4781(02)00482-7 . ПМИД 12359337 .
- Фатеми М., Герман А., Гоуэр Х., Йельч А. (октябрь 2002 г.). «Dnmt3a и Dnmt1 функционально взаимодействуют во время метилирования ДНК de novo» . Европейский журнал биохимии . 269 (20): 4981–4984. дои : 10.1046/j.1432-1033.2002.03198.x . ПМИД 12383256 .
- Вайзенбергер DJ, Велическу М., Пресиадо-Лопес М.А., Гонсалес Ф.А., Цай Ю.К., Лян Г., Джонс П.А. (сентябрь 2002 г.). «Идентификация и характеристика альтернативно сплайсированных вариантов ДНК-метилтрансферазы 3а в клетках млекопитающих». Джин . 298 (1): 91–99. дои : 10.1016/S0378-1119(02)00976-9 . ПМИД 12406579 .
- Чедин Ф., Либер М.Р., Се К.Л. (декабрь 2002 г.). «ДНК-метилтрансферазоподобный белок DNMT3L стимулирует метилирование de novo с помощью Dnmt3a» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (26): 16916–16921. Бибкод : 2002PNAS...9916916C . дои : 10.1073/pnas.262443999 . ПМК 139244 . ПМИД 12481029 .
- Роберт М.Ф., Морен С., Болье Н., Готье Ф., Чуть И.С., Барсалу А., Маклеод А.Р. (январь 2003 г.). «DNMT1 необходим для поддержания метилирования CpG и аберрантного молчания генов в раковых клетках человека». Природная генетика . 33 (1): 61–65. дои : 10.1038/ng1068 . ПМИД 12496760 . S2CID 561490 .
- Фукс Ф., Херд П.Дж., Деплюс Р., Кузаридес Т. (май 2003 г.). «ДНК-метилтрансферазы связываются с HP1 и гистон-метилтрансферазой SUV39H1» . Исследования нуклеиновых кислот . 31 (9): 2305–2312. дои : 10.1093/нар/gkg332 . ПМК 154218 . ПМИД 12711675 .
- Якусидзи Т., Узава К., Шибахара Т., Нома Х., Танзава Х. (июнь 2003 г.). «Сверхэкспрессия ДНК-метилтрансфераз и статус метилирования гена CDKN2A при плоскоклеточном раке полости рта». Международный журнал онкологии . 22 (6): 1201–1207. дои : 10.3892/ijo.22.6.1201 . ПМИД 12738984 .
- Линг Ю., Санкпал Ю.Т., Робертсон А.К., МакНелли Дж.Г., Карпова Т., Робертсон К.Д. (2004). «Модификация ДНК-метилтрансферазы 3a (Dnmt3a) de novo с помощью SUMO-1 модулирует ее взаимодействие с деацетилазами гистонов (HDAC) и ее способность подавлять транскрипцию» . Исследования нуклеиновых кислот . 32 (2): 598–610. дои : 10.1093/nar/gkh195 . ПМЦ 373322 . ПМИД 14752048 .