Сидерокалин
| Липокалин/эпидидимал-специфический липокалин-12, ассоциированная с нейтрофильной гелатиназой, липокалин | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | Lcn2/lcn12ipr003087siderocalin | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | GeneCards : [1] ; OMA : - ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викидид | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
Сидерокалин (SCN) , липокалин-2 , NGAL , 24P3 млекопитающего -белок типа липокалина , который может предотвратить получение железа патогенными бактериями путем связывания сидерофоров , которые представляют собой железо, которые являются хелаторами , изготовленными микроорганизмами. [ 1 ] [ 2 ] Железо служит ключевым питательным веществом во взаимодействиях хозяина - патогена , а патогены могут получать железо от организма хозяина посредством синтеза и высвобождения сидерофоров, таких как энтеробактин . [ 3 ] Сидерокалин является частью механизма защиты млекопитающих и действует как антибактериальный агент . [ 1 ] [ 4 ] [ 5 ] [ 6 ] [ 7 ] Кристаллографические исследования SCN продемонстрировали, что он включает в себя чашечку , лиганд -связывающий домен , который облицован полярными катионными группами. [ 8 ] Центральным в механизме распознавания сидерофора/ сидерокалина являются гибридные электростатические катионные / взаимодействия . [ 5 ] [ 9 ] Чтобы уклониться от защиты хозяина, патогены эволюционировали для получения структурно разнообразных сидерофоров, которые не будут распознаваться сидерокалином, что позволяет бактериям приобретать железо. [ 1 ]
Требования к железу организмов хозяина
[ редактировать ]Организмы требуют железа для различных химических реакций . [ 10 ] Хотя железо может быть обнаружено по всей биосферу , свободный железо железа образует нерастворимые гидроксиды при физиологическом рН , ограничивая его доступность в аэробных условиях живыми организмами. [ 10 ] [ 11 ] Чтобы сохранить гомеостаз , организмы развили специфические белковые сети, с белками и рецепторами, переведенными в соответствии с уровнями внутриклеточного железа. [ 10 ] [ 12 ] Экспорт и импорт дополняются процессом циклирования между железом Fe (II), в уменьшенной среде клетки доступным , и железа Fe (III), обнаруженного в основном в аэробных условиях. [ 13 ] [ 14 ] Механизмы получения железа патогенных бактерий демонстрируют роль железа как ключевого компонента на границе между патогенами и хозяевами. [ 13 ] [ 14 ]
Семейство липокалинов белков связывания железа
[ редактировать ]Семейство липокалинов связывающих белков продуцируется иммунной системой и секвестрами оборотных сидерофорных комплексов из сидерофорских рецепторов бактерий. [ 15 ] [ 16 ] Семейство липокалинов связывающих белков обычно имеет консервативную восьмицепочечную складку β-бокса чашечки с сайтом связывания , [ 16 ] [ 17 ] которые облицованы положительно заряженными аминокислотными остатками, что позволяет связывать взаимодействие с сидерофорами. [ Цитация необходима ]
Клиническое значение
[ редактировать ]Микобактериальные инфекции
[ редактировать ]Липокалин сидерокалин обнаруживается в нейтрофильных гранул матки , выделениях и на особенно высоких уровнях в сыворотке во время бактериальной инфекции . [ 4 ] После инфекции патогены используют сидерофоры для захвата железа из организма хозяина. [ 18 ] Эта стратегия, однако, осложняется сидерокалином белка человека, который может секвестр сидерофоров и предотвращать их использование патогенными бактериями в качестве агентов доставки железа. [ 19 ] Этот эффект был продемонстрирован в исследованиях с мышами, нокаутированными на сидерокалине , которые более чувствительны к инфекциям в условиях ограничения железа. [ 4 ] [ 5 ]
Микобактериальная вирулентность
[ редактировать ]
Сидерофоры являются железными хелаторами, что позволяет организмам приобретать железо из окружающей среды. В случае патогенов железо может быть получено у организма хозяина. [ 20 ] Сидерофоры и железо железа могут ассоциироваться с образованием стабильных комплексов. [ 10 ] [ 21 ] [ 22 ] Сидерофоры связывают железо, используя различные лиганды , чаще всего в виде α-гидроксикарбоксилатов (например, цитрат), катехолатов и гидроксаматов . [ 5 ] [ 10 ] [ 23 ] [ 24 ] Как защитный механизм, сидерокалин может заменить комплексы бис-катехолов железа (образуется в физиологических условиях) третьим катехолом, чтобы достичь гексакоординатного комплекса железа, что приводит к более высокому аффинному связыванию. [ 5 ] [ 18 ] [ 25 ]
Как посредник транспорта железа млекопитающих
[ редактировать ]Сидерофоры млекопитающих, в частности, катехолы , можно найти в кишечнике человека и в сидерофорах, таких как энтеробактин , и служат железными связывающими фрагментами . [ 5 ] [ 26 ] Катехол, напоминающие молекулы, могут действовать как железные лиганды в клетке и в систематической циркуляции , позволяя сидерокалину связываться с железо-катехолом. [ 27 ] Катехолы могут быть связаны сидерокалином, в виде свободных лигандов или в железном комплексе. [ 28 ] 24p3 является рецептором липокалина-2 позвоночных , который позволяет импортировать комплекс сидерофора железа в клетки млекопитающих. [ 27 ] Во время почки эмбриогенеза происходит опосредованный сидерокалином транспорт железа, поскольку концентрация железа должна быть высоко контролируется, чтобы ограничить воспаление . [ 4 ] [ 11 ] После секреции нейтрофилами сидерокалин может связываться с патогенными сидерофорами, такими как бациллибактин , и предотвращать транспортировку сидерофора. [ 29 ] Сидерокалин был связан с различными клеточными процессами, кроме транспорта железа, включая апоптоз , клеточную дифференцировку , онкогенез и метастазирование . [ 10 ] [ 30 ]
Структура
[ редактировать ]Птицы ортологов боковыми сидерокалина (Q83 и EX-FABP) и NGAL ( ассоциированная с нейтрофильной желатиназой липокалином-2) содержат CALYCE с положительно заряженными лизином и аргининовыми цепями. [ 8 ] [ 30 ] [ 31 ] [ 32 ] [ 33 ] Эти боковые цепи взаимодействуют через катион-PI и кулоновские взаимодействия с отрицательно заряженными сидерофорами, которые содержат ароматические катехолярные группы. [ 10 ] [ 30 ] Кристаллографические исследования сидерокалина показали, что лиганд -связывающий домен SCN, известный как чашечка, является мелким и широким и облицован полярными катионными группами из трех положительно заряженных остатков ARG81, Lys125 и Lys134. [ 5 ] [ 8 ] [ 34 ] SCN также может связывать неферерические комплексы и был идентифицирован как потенциальный транспортер для тяжелых актинид-ионов. Были получены кристаллические структуры SCN, содержащие тяжелые металлы (торий, плутоний, американец, Curium и Californium). [ 35 ] [ 36 ] SCN был найден в виде мономера , гомомерного или тримера в плазме человека. [ 5 ] Складка сидерокалина исключительно стабильна. [ 4 ] [ 5 ] Чашечка является структурно стабильной и жесткой, а конформационные изменения обычно не происходят при изменении pH , ионной силы или связывания лиганда. [ 5 ]
Перепляющий карман
[ редактировать ]Структурная стабильность чашечки была отнесена к трем карманам связывания в чашечке, которые стерически ограничивают, что лиганды совместимы с сидерокалином. [ 5 ] [ 8 ] Calyx SCN может вместить три ароматических кольца катехолярных фрагментов, в трех доступных карманах для связывания. [ 5 ] [ 28 ] Структурные результаты с твердым состоянием и раствором продемонстрировали, что энтеробактин, полученный из бактерий, связан с связывающим карманом SCN, что позволяет участвовать в SCN в остром иммунном ответе на бактериальную инфекцию. [ 5 ] [ 21 ] Одним из методов, с помощью которого патогенные микроорганизмы могут обходить механизмы иммунитета , является модификация химической структуры сидерофора для предотвращения взаимодействия с SCN. [ 24 ] Одним из примеров является добавление молекул глюкозы в энтеробактин сальмохелина (C- глюкозилированный энтеробактин), чтобы повысить гидрофильность и объемность сидерофора и ингибировать связывание с SCN. [ 24 ] [ 37 ]
Связывающие взаимодействия
[ редактировать ]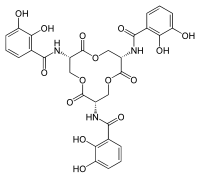
Сидерофоры, как правило, связаны с сидерокалином с субнамолярным сродством и конкретно взаимодействуют с сидерокалином. [ 10 ] [ 25 ] Значение KD взаимодействия сидерокалина/сидерофора, измеренное с помощью гашения флуоресценции (KD = 0,4 нм), указывает на то, что сидерокалин может захватывать сидерофоры с высокой аффинностью. [ 31 ] [ 38 ] Это значение KD аналогично значению бактериального рецептора FEPA (KD = 0,3 нм). [ 5 ] Связывание сидерофора/сидерокалина направлено электростатическими взаимодействиями. [ 5 ] [ 38 ] В частности, механизм включает в себя гибридные электростатические и катион-PI взаимодействия в положительно заряженном белке. [ 25 ] Сидерофор расположен в центре сидерокалиновой чашечки и связан с множественными прямыми полярными взаимодействиями. [ 25 ] Структурный анализ взаимодействия сидерокалина/сидерофора показал, что сидерофор сопровождается плохим и диффузным качеством электронной плотности , при этом большая часть лиганда подвергается воздействию растворителя , когда сидерофор находится в чашечке. [ 5 ] [ 6 ] Сидерокалин обычно не связывает сидерофоры на основе гидроксамата , потому что эти субстраты не имеют необходимой ароматической электронной структуры для взаимодействия катион -PI. [ 5 ] [ 25 ] Для получения железа в присутствии сидерокалина патогенные бактерии используют несколько сидерофоров, которые не связываются с сидерокалином или структурно модифицируют сидерофоры, чтобы ингибировать связывание сидерокалина. [ 5 ] [ 39 ] Сидерокалин может связывать растворимые сидерофоры микобактерий , включая карбоксимикобактины. [ 5 ] [ 6 ] Исследования in vivo показали, что взаимодействие связывания между карбоксимикобактином и сидерокалином служит для защиты организма хозяина от микобактериальных инфекций, при этом сидерокалин ингибирует приобретение микобактериального железа. [ 5 ] [ 28 ] [ 40 ] Сидерокалин может секвестр карбоксимикобактинов железа, используя механизм полиспецифического распознавания. [ 5 ] Механизм распознавания сидерофора/сидерокалина в первую очередь включает в себя гибридные электростатические/катиони-PI взаимодействия. [ 5 ] [ 9 ] [ 11 ] Хвосты жирных кислот карбоксимикобактина находятся в конформации «хвоста или« хвоста »в кармане 2. [ 5 ] Конформация «хвоста в» длины цепи жирной кислоты вводит значительное взаимодействие между чашечкой и лигандом, увеличивая сродство сидерокалинового чалиса и карбоксимикобактина. [ 5 ] Хвосты жирной кислоты коротких длины имеют соответственно менее благоприятную связующую среду с сидерокалином и не могут поддерживать необходимое взаимодействие с связывающим карманом. [ 5 ] Поскольку липокалин-2 не может связываться с карбоксимикобактинами цепи длинных жирных кислот микобактерий, очевидно, что ряд патогенов развивались, чтобы избежать активности липокалина-2. [ 41 ]
Механизм распознавания
[ редактировать ]Электростатическое взаимодействие играет ключевую роль в механизме распознавания сидерофоров сидерокалином. [ 1 ] Связывание сидерофора и сидерокалина связывающего кармана в первую очередь направлена катион-PI-взаимодействиями, с положительно заряженным связывающим карманом сидерокалина, притягивающим отрицательно заряженный комплекс. [ 1 ] Структурный фактор, связанный с механизмом распознавания сидерокалина, сидерофоров фенолята / катехолата, включает в себя линкер основной цепь, который позволяет сидерокалину взаимодействовать с различными сидерофорами фенолята / катехолата. [ 4 ] [ 42 ] В то время как распознавание сидерокалина минимально влияет замена различных металлов, метилирование трех катехолатных колец энтеробактина может препятствовать распознаванию сидерокалина. [ 5 ] [ 34 ] [ 38 ] [ 43 ] Стратегия, используемая патогенами для преодоления иммунного ответа , - это производство сидерофоров, которые не будут распознаваться сидерокалином. [ 19 ] [ 44 ] Например, сидерокалин не может распознавать сидерофоры C-глюкозилированного аналога энтеробактина, поскольку донорские группы являются гликозилированными , вводя стерические взаимодействия в позиции 5- углерод катехольных групп. [ 1 ] [ 24 ]
История
[ редактировать ]Требование железа от людей и патогенов было известно много лет. [ 10 ] Связь между железом и микобактамином, железо-хелатирующими факторами роста из микобактерий была впервые сделана в 1960-х годах. [ 5 ] В то время интерес растут к разрешению применения микобактинов в качестве целевых молекул для рационального антиберкулеза . [ 5 ] [ 45 ] Эксперименты в 1960 -х и 1970 -х годах показали, что дефицит железа в микобактериях был причиной « анемичных » клеток . [ 46 ] Большинство генов и систем, необходимых для приобретения железа с высокой аффинностью, были идентифицированы в патогенных и сапрофитных микобактериях. [ 5 ] Эти гены кодируют белки для хранения железа, поглощения железами-сидерофоров и гема . [ 5 ] [ 47 ] Люди эволюционировали защиту от сидерофор-опосредованного приобретения железа путем разработки сидерокалина. Чтобы бороться с этим, различные патогены развили сидерофоры, которые могут избежать распознавания сидерокалина. [ 5 ] Было показано, что сидерокалин связывается с сидерофорами и ингибирует приобретение железа и предотвращает рост туберкулеза Mycobacterium во внеклеточных культурах; Однако влияние сидерокалина на этот патоген в макрофагах остается неясным. [ 24 ] [ 31 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а беременный в дюймовый и фон Ван Эльдик Р., Хаббард CD (2009). Достижения в неорганической химии Том 61 (1 -е изд.). Лондон, Великобритания: Elsevier. С. 237–239. ISBN 9780123750334 Полем Получено 16 февраля 2015 года .
- ^ Correnti C, Richardson V, Sia, Bondayeke AD, Ruiz M, Suryo Rahmanto Y, Kovačević ž, Clifton MC, Holmes MA, Kaisi BK, Barasch J, Raymond KN, Richardd DR, Strong Rk (2012). «Сидерокалин/LCN2/NGAL/24P3 не управляют кислотой через снятие железа, опосредованное гентисновой кислотой, в гемопоэтических клеточных линиях » Plos один 7 (8): E4 Bibcode : 2012plose ... 743696c Doi : 10.1371/ journal.pone.0 PMC 3424236 PMID 22928018
- ^ Cherayil BJ (май 2011 г.). «Роль железа в иммунном ответе на бактериальную инфекцию» . Иммунологические исследования . 50 (1): 1–9. doi : 10.1007/s12026-010-8199-1 . PMC 3085559 . PMID 21161695 .
- ^ Jump up to: а беременный в дюймовый и фон Paragas N, Qiu A, Hollmen M, Nickolas TL, Devarajan P, Barasch J (сентябрь 2012 г.). «Нгалл-сидрокал при заболевании почек » Biochimica et Biophysica Acta (BB) - исследования молекулярных клеток 1823 (9): 1451–8 Doi : 10.1016/ j.bmcr.2012.06.0 3664277PMC 22728330PMID
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. ведущий с Т в v В х и С аа Аб и объявление Но Byers BR (2013). Приобретение железа с помощью рода Mycobacterium . Springerbriefs в молекулярной науке. Спрингер. С. 1–88. doi : 10.1007/978-3-319-00303-0 . ISBN 978-3-319-00303-0 Полем S2CID 12666634 .
- ^ Jump up to: а беременный в Holmes MA, Paulsene W, Jide X, Ratledge C, Strong RK (январь 2005 г.). «Сидерокалин (LCN 2) также связывает карбоксимикобактины, потенциально защищая от микобактериальных инфекций посредством секвестрации железа» . Структура 13 (1): 29–41. doi : 10.1016/j.str.2004.10.009 . PMID 15642259 .
- ^ Sige L A, Sigel H, Sige L RK (2013). Взаимосвязи между важными ионами металлов и заболеваниями человека . Гейдельберг, Германия: Спрингер. С. 282–283. ISBN 9789400774995 Полем Получено 14 февраля 2015 года .
- ^ Jump up to: а беременный в дюймовый Sia AK, Allred BE, Raymond KN (апрель 2013 г.). «Сидерокалины: сидерофоры, связывающие белки, развивались для обороны первичной патогена -хозяина» . Современное мнение о химической биологии . 17 (2): 150–7. doi : 10.1016/j.cbpa.2012.11.014 . PMC 3634885 . PMID 23265976 .
- ^ Jump up to: а беременный Абергель Р.Дж., Уилсон М.К., Арсено Дж., Хотт Т.М., Стронг Р.К., Байерс Б.Р., Рэймонд К.Н. (декабрь 2006 г.). «Антракс -патоген уклоняется от иммунной системы млекопитающих через производство стелс -сидерофора» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (49): 18499–503. Bibcode : 2006pnas..10318499a . doi : 10.1073/pnas.0607055103 . PMC 1693691 . PMID 17132740 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Correnti C, Strong RK (апрель 2012 г.). «Сидерофоры млекопитающих, сидерофоры, связывающие липокалины и лабильный железный бассейн» . Журнал биологической химии . 287 (17): 13524–31. doi : 10.1074/jbc.r111.311829 . PMC 3340207 . PMID 22389496 .
- ^ Jump up to: а беременный в Чакраборти Р., Браун В., Хантке К., Корнелис П. (2013). Поглощение железа у бактерий с акцентом на E. coli и Pseudomonas . Springerbriefs в биометалах. С. 31–66. ISBN 978-94-007-6087-5 .
- ^ Ganz T (октябрь 2013). «Системный железный гомеостаз». Физиологические обзоры . 93 (4): 1721–41. doi : 10.1152/physrev.00008.2013 . PMID 24137020 .
- ^ Jump up to: а беременный Doherty CP (май 2007 г.). «Взаимодействие хозяина-патогена: роль железа» . Журнал питания . 137 (5): 1341–4. doi : 10.1093/jn/137.5.1341 . PMID 17449603 .
- ^ Jump up to: а беременный Skaar EP (2010). «Битва за железо между бактериальными патогенами и их хозяевами позвоночных» . PLO -патогены . 6 (8): E1000949. doi : 10.1371/journal.ppat.1000949 . PMC 2920840 . PMID 20711357 .
- ^ Сэнди М, Батлер А (октябрь 2009 г.). «Микробное приобретение железа: морские и наземные сидерофоры» . Химические обзоры . 109 (10): 4580–95. doi : 10.1021/cr9002787 . PMC 2761978 . PMID 19772347 .
- ^ Jump up to: а беременный Цветок DR (август 1996 г.). «Семейство липокалиновых белков: структура и функция» . Биохимический журнал . 318 (1): 1–14. doi : 10.1042/bj3180001 . PMC 1217580 . PMID 8761444 .
- ^ Fuentes-Prior P, Noeske-Jungblut C, Donner P, Schleuning WD, Huber R, Bode W (октябрь 1997 г.). «Структура тромбинного комплекса с триабином, липокалиноподобный экзозит-связывающий ингибитор, полученный в результате пуатомановой ошибки» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (22): 11845–50. Bibcode : 1997pnas ... 9411845f . doi : 10.1073/pnas.94.22.11845 . PMC 23629 . PMID 9342325 .
- ^ Jump up to: а беременный Miethke M, Marahiel Ma (сентябрь 2007 г.). «Приобретение железа на основе сидерофора и контроль патогенов» . Микробиология и молекулярная биология обзоры . 71 (3): 413–51. doi : 10.1128/mmbr.00012-07 . PMC 2168645 . PMID 17804665 .
- ^ Jump up to: а беременный Allred BE, Correnti C, Clifton MC, Strong RK, Raymond KN (сентябрь 2013). «Сидерокалин переоценивает координационную химию вицибактина, сидерофора вибрио -холеры» . ACS Химическая биология . 8 (9): 1882–7. doi : 10.1021/cb4002555 . PMC 3783644 . PMID 23755875 .
- ^ Miethke M (январь 2013). «Молекулярные стратегии микробной ассимиляции железа: от высокоаффинных комплексов до систем сборки кофакторов» . Металломика . 5 (1): 15–28. doi : 10.1039/c2mt20193c . PMID 23192658 .
- ^ Jump up to: а беременный Абергель Р.Дж., Клифтон М.К., Пизарро Дж.С., Уорнер Дж. А., Шух Д.К., Стронг Р.К., Рэймонд К.Н. (август 2008 г.). «Взаимодействие сидерокалина/энтеробактина: связь между иммунитетом млекопитающих и бактериальным транспортом железа» . Журнал Американского химического общества . 130 (34): 11524–34. doi : 10.1021/ja803524w . PMC 3188318 . PMID 18680288 .
- ^ Fukushima T, Allred BE, Sia AK, Nichiporuk R, Andersen UN, Raymond KN (август 2013 г.). «Грамположительная сидерофора с обменом железом от Fe-siderophore до апо-сидедерофора Bacillus cereus yxeb» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (34): 13821–6. BIBCODE : 2013PNAS..11013821F . doi : 10.1073/pnas.1304235110 . PMC 3752266 . PMID 23924612 .
- ^ Dhungana S, Harrington JM, Gebhardt P, Möllmann U, Crumbliss AL (октябрь 2007 г.). «Уравновешивание хелатирования железа, окислительно -восстановительное и сидерофоры аналогов Ferrichrome Suderophore платформы» (PDF) . Неорганическая химия . 46 (20): 8362–71. doi : 10.1021/ic070158l . PMID 17824601 .
- ^ Jump up to: а беременный в дюймовый и Yehuda S, Monsofsky DI (2010). Дефицит железа и перегрузка: от основной биологии до клинической медицины . Нью -Йорк, Нью -Йорк: Humana Press. С. 66–69. ISBN 9781934115220 Полем Получено 14 февраля 2015 года .
- ^ Jump up to: а беременный в дюймовый и Hoette TM, Abergel RJ, Xu J, Strong Rk, Raymond KN (Dec 2008). «Роль электростатики в распознавании сидерофора иммунопротеином сидерокалином» . Журнал Американского химического общества . 130 (51): 17584–92. doi : 10.1021/ja8074665 . PMC 2778733 . PMID 19053425 .
- ^ Роджерс HJ (март 1973 г.). «Железные связывающие катехолы и вирулентность в Escherichia coli» . Инфекция и иммунитет . 7 (3): 445–56. doi : 10.1128/iai.7.3.445-456.1973 . PMC 422698 . PMID 16558077 .
- ^ Jump up to: а беременный Андерсон Г.Дж., Макларен Г.Д. (2012). Железная физиология и патофизиология у людей . Нью -Йорк, Нью -Йорк: Спрингер. с. 237–239, 658. ISBN 9781603274845 Полем Получено 14 февраля 2015 года .
- ^ Jump up to: а беременный в Bao G, Clifton M, Hoette TM, Mori K, Deng SX, Qiu A, Voltardid M, Williams D, Paragas N, Leete T, Culmanni R, Li X, Lee B, Kalandadze A, Ratner AJ, Pizarro JC, Schmidt- , Schmidt-Ott KM, Landry DW, Raymond KN, Strong RK, Barasch J (август 2010 г.). «Транспортировка железа в обращении, связанном с комплексом сидерокалина (NGAL) -катехол » Природная химическая биология 6 (8): 602–9 Doi : 10.1038/nchembio . PMC 2907470 PMID 20581821
- ^ Бергман Н.Х. (2011). Bacillus anthracis и сибирская язва . Хобокен, Нью -Джерси: Уайли. с. Глава 7. ISBN 9781118148082 Полем Получено 14 февраля 2015 года .
- ^ Jump up to: а беременный в Correnti C, Clifton MC, Abergel RJ, Allred B, Hoette TM, Ruiz M, Cancedda R, Raymond KN, Descalzi F, Strong RK (Dec 2011). «Galline Ex-FABP-это антибактериальный сидерокалин и датчик лизофосфатидной кислоты, функционирующий с помощью специфичности двойного лиганда» . Структура 19 (12): 1796–806. doi : 10.1016/j.str.2011.09.019 . PMC 3240821 . PMID 22153502 .
- ^ Jump up to: а беременный в Эштон Актон Q (2012). Достижения в области исследований и применения Serine (2012: Scholarylybrief Ed.). Атланта, Джорджия: научная динамика. С. 42–43. ISBN 9781481614276 Полем Получено 14 февраля 2015 года .
- ^ Thongboonkerd V (2007). Протеомика жидкостей человеческого тела: принципы, методы и применение . Тотова, Нью -Джерси: Humana Press. С. 338–339. ISBN 9781597454322 Полем Получено 16 февраля 2015 года .
- ^ Clifton MC, Corrent C, Strong RK (август 2009 г.). «Сидерокалины: связывающие сидерофоры белки врожденной иммунной системы» (PDF) . Биометалы . 22 (4): 557–64. doi : 10.1007/s10534-009-9207-6 . PMID 19184458 . S2CID 8776376 .
- ^ Jump up to: а беременный Hoette TM, Clifton MC, Zawadzka AM, Holmes MA, Strong RK, Raymond KN (Dec 2011). «Иммунное вмешательство в микобактерию туберкулеза внутриклеточного приобретения железа посредством распознавания сидерокалина карбоксимикобактинов» . ACS Химическая биология . 6 (12): 1327–31. doi : 10.1021/cb200331g . PMC 3241878 . PMID 21978368 .
- ^ Deblonde, Gauthier J.-P.; Sturzbecher-Hoehne, Manuel; Руперт, Питер Б.; Далия, и д.; Илли Мари-Клэр; Ralston, Corie Y.; Brabec, Jiri; де Йонг, Wide A.; Стронг, Роланд (сентябрь 2017 г.). «Хелатирование и стабилизация беркелия в состоянии окисления +IV» (PDF) . Природная химия . 9 (9): 843–849. Bibcode : 2017natch ... 9..843d . doi : 10.1038/nchem.2759 . ISSN 1755-4349 . Ости 1436161 . PMID 28837177 .
- ^ Капитан Илья; Deblonde, Gauthier J.-P.; Руперт, Питер Б.; An, Dahlia D.; Илли, Мари-Клэр; Ростан, Эмелин; Ralston, Corie Y.; Сильный, Роланд К.; Абергель, Ребекка Дж. (2016-11-21). «Инженерное распознавание тетравалентного циркония и тория хелаторно -белковыми системами: к гибкой лучевой терапии и платформам визуализации» . Неорганическая химия . 55 (22): 11930–11936. doi : 10.1021/acs.inorgchem.6b02041 . ISSN 0020-1669 . Ости 1458481 . PMID 27802058 .
- ^ Alvarez MV (2007). Выделение, структура и обнаружение сальмохелинов: новые сидерофоры у энтеробактерий . Геттинген, Германия: Кювильер Верлаг. С. 29–34. ISBN 9783867271097 Полем Получено 14 февраля 2015 года .
- ^ Jump up to: а беременный в Абергель Р.Дж., Мур Э.Г., Стронг Р.К., Рэймонд К.Н. (август 2006 г.). «Микробное уклонение иммунной системы: структурные модификации энтеробактина нарушают распознавание сидерокалина» . Журнал Американского химического общества . 128 (34): 10998–9. doi : 10.1021/ja062476+ . PMC 3188317 . PMID 16925397 .
- ^ Allela L, Boury O, Pouillot R, Délicat A, Yaba P, Kumulungui B, Rouquet P, Gonzalez JP, Leroy EM (Mar 2005). «Распространенность антител вируса Эбола у собак и риск человека» . Возникающие инфекционные заболевания . 11 (3): 385–90. doi : 10.3201/eid1103.040981 . PMC 3298261 . PMID 15757552 .
- ^ Åkerström B (2006). Липокалины . Остин, Техас: Landes Bioscience. П. 9781587062971 .
- ^ Kidd SP (2011). Стресс -реакция в патогенных бактериях, том 19 достижений в молекулярной и клеточной микробиологии . Уоллингфорд, Великобритания: Каби. С. 287–290. ISBN 9781845937775 Полем Получено 14 февраля 2015 года .
- ^ Сильный, rk; Akerstrom, B.; Borregaard, N.; Цветок, доктор; Salier, J.-P. (Ред.). «Сидерокалины» (PDF) . Фред Хатчинсон РАССКИЙ РАССКОЛЬНЫЙ ЦЕНТР.
- ^ Абергель Р.Дж., Уорнер Дж.А., Шух Д.К., Рэймонд К.Н. (июль 2006 г.). «Протонирование энтеробактина и высвобождение железа: структурная характеристика координационного сдвига салицилата в энтеробактине железа» . Журнал Американского химического общества . 128 (27): 8920–31. doi : 10.1021/ja062046j . PMC 3188320 . PMID 16819888 .
- ^ Stintzi A, Barnes C, Xu J, Raymond KN (сентябрь 2000). «Микробный транспорт железа через шаттл сидерофора: парадигма переноса мембраны» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (20): 10691–6. Bibcode : 2000pnas ... 9710691S . doi : 10.1073/pnas.200318797 . PMC 27084 . PMID 10995480 .
- ^ Barclay R, Ratledge C (март 1983 г.). «Железные соединения Mycobacterium avium, M. intracellulare, M. scrofulaceum и микобактин-зависимый M. paratuberculosis и M. avium» . Журнал бактериологии . 153 (3): 1138–46. doi : 10.1128/jb.153.3.1138-1146.1983 . PMC 221756 . PMID 6826517 .
- ^ Jamison DT, Breman JG, Teamham AR, Alleyne G, Claeson M, Evans DB, Jha P, Mills A, Musgrove P (2006). Приоритеты контроля заболевания в развивающихся странах (2 -е изд.). Вашингтон, округ Колумбия: Всемирный банк. с. Глава 16. ISBN 978-0-8213-6179-5 Полем Получено 16 февраля 2015 года .
- ^ Caza M, Kronstad JW (2013). «Общие и четкие механизмы приобретения железа бактериальными и грибковыми патогенами людей» . Границы в клеточной и инфекционной микробиологии . 3 (80): 80. doi : 10.3389/fcimb.2013.00080 . PMC 3832793 . PMID 24312900 .