Rhynchocephalia
| Rhynchocephalians | |
|---|---|

| |
| Tuatara ( ) Sphenodon Punctatus , единственный живой реинчоцефаец | |

| |
| Ископаемое Вадасавр , ринчоцефалиан из позднего юрского периода Германии | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Сорт: | Рептилия |
| Суперкаус: | Лепидозаурия |
| Заказ: | Rhynchocephalia Гюнтер 1867 |
| Тип видов | |
| Sphenodon punctatus Грей , 1842
| |
| Подгруппы | |
|
Смотрите текст | |
Rhynchocephalia ( / ˌ r ɪ ŋ k oʊ s ˈ ˈ f eɪ l i ə / ; lit. ' -головы ' ) порядок похожие ящери , это - на ’ Зеландия . Несмотря на нынешнее отсутствие разнообразия, во время мезозойских ренчоцефалов была специфическая группа с высоким морфологическим и экологическим разнообразием. Самая старая запись группы датируется средним триасом примерно 238-240 миллионов лет назад, и они достигли глобального распределения ранней юрской игрой . [ 1 ] Большинство ренчоцефалов принадлежат к группе Sphenodontia («Wedge-Teeth»). Их ближайшие живые родственники - это ящерицы и змеи в скамате , с двумя порядками, сгруппированными вместе в Lepidosauria Super Dorder .
После того, как представляя доминирующую в мире группу мелких рептилий, многие из ниш, занятых сегодня ящерицами, были проведены ринчоцефальцами во время триасы и юрского периода . Ринчоцефалии подвергались большому спаду во время мелового периода , и они почти полностью исчезли в начале кайнозоя . В то время как современная туатара в первую очередь является насекомым и плотоядным , разнообразие группы также включало травоядные эйленодонтины , и были другие ренчоцефалы со специализированными экологиями, такими как дурофагские сапеозары . Были даже успешные группы водных спенодонтов, таких как плеврозавры . [ 2 ]
История
[ редактировать ]Туатара первоначально были классифицированы как агамидские ящерицы , когда их впервые описали Джон Эдвард Грей в 1831 году. Они оставались неправильно классифицированными до 1867 года, когда Альберт Гюнтер из Британского музея отметил, что особенности, похожие на птиц, черепах и крокодилов. Он предложил приказ Rhynchocephalia (то есть «голова клюва») для туатары и его ископаемых родственников. [ 3 ] В 1925 году Сэмюэль Венделл Уиллистон предложил Sphenodontia включить только Туатару и их ближайшие родственники окаменелостей. [ 4 ] Sphenodon получен из греческого σφήν sphen 'wedge' и ὀΔούς Odous 'зуб'. [ 5 ] [ 6 ] [ 7 ] Впоследствии многие неприятные виды были добавлены в Rhynchocephalia, что привело к тому, что таксономисты называют « таксоном мусора ». К ним относятся поверхностно схожие (как по форме, так и в названии), но не связанные с ринчозаврами , которые жили в триасе . [ 4 ] Исследования в 1970-х и 1980-х годах показали, что ренчозавры не были связаны, при этом компьютерный кладистический анализ, проведенный в 1980-х годах, обеспечивал надежный диагноз для определения группы. [ 8 ]
Анатомия
[ редактировать ]
Rhynchocephalia и их дочерняя группа Squamata (в том числе ящерицы, змеи и амфисбаэниан ) принадлежат к суперпорядочке Lepidosauria , единственному выжившему таксону в Lepidosauromorpha .
Скваматы и ренчоцефалии имеют ряд общих признаков ( синапоморфий ), включая плоскости разрушения в хвостовых позвонках, позволяющих каудальной аутотомии (потеря хвоста при угрозе), поперечные щели , прорезь, отверстие в таблице, известно Дополнительные центры усиления костей конечностей в эпифизе , коленного сустава, где боковое углубление на бедре позволяет артикулировать малоберцовую кость, развитие сексуального сегмента почки и ряд черт костей, включая слитый астралаго - Calcaneun и увеличенный четвертый дистальный лап , который создает новый сустав, а также подключенная пятая плюсневка . [ 9 ]
Как и некоторые ящерицы, туатара обладает теменным глазом (также называемым шишковидным глазом или третьим глазом) в верхней части головы, образованной парапинеальным органом, с сопровождающим отверстием в крыше черепа, окруженной теменными костями , названная « Pineeal Foramen », которая также присутствует в ископаемых ренчоцефалиях. Париетальный глаз обнаруживает световой контроль дневных ночных и сезонных циклов, помогая регулировать циркадный ритм , а также другие функции. [ 10 ] [ 11 ] [ 12 ] [ 13 ] В то время как глаз шишковидных изделий были широко распространены среди ранних позвоночных, в том числе ранние рептилии, они были потеряны среди большинства живых групп. [ 12 ]
Ринчоцефальцы отличаются от скваматов по ряду признаков, включая удержание гастралии (похожие на ребристые кости, присутствующие в животе тела, присутствующие в тетраподах , а также присутствуют у живых крокодильцев ). [ 14 ] В отличие от сквамов, но похожих на большинство птиц, у туатары не хватает пениса. Это вторичная потеря, так как пенис или гемипен , похожие на квамут , вероятно, присутствовали в последнем общем предке ренчоцефалов и сквамов. [ 15 ]

Полный нижний височный стержень (вызванный слиянием мугальных и квадроциклевых / квадратоджугальных костей черепа) туатары, часто исторически утверждается, что является примитивной особенностью, удерживаемой от более ранних рептилий, на самом деле является производной среди спгенодонтов, с примитивными лепидозаврафов, на самом деле является производной среди спенодонтов, с примитивными лепидозаврифами, на самом деле является полученной особенностью среди спенодонтов, с примитивными лепидозаврифами. и многие ринчоцефалии, в том числе самые примитивные, имеющие открытый нижний височный фенестра без височной бары. [ 16 ] [ 17 ] В то время как часто не хватает полной височной полосы, подавляющее большинство ренчоцефальцев имеют сзади -направленный процесс (разгибание) югольной кости. У всех известных ринчоцефалов не хватает селеневой кости, присутствующей в нижней челюсти более примитивных рептилий, [ 18 ] С черепами всех членов Sphenodontia не хватает ласковых костей . [ 19 ] У большинства ренчоцефалов также есть лобные кости черепа. [ 20 ] [ 18 ] В то время как ранние ринчоцефалии обладали барабанной перепонкой в ухе и соответствующей квадратной раковиной, аналогичной тем, которые обнаружены у ящериц, они были потеряны в туатаре и, вероятно, других полученных ринчоцефалиях. Эта потеря может быть связана с развитием движения взад -вперед нижней челюсти. [ 21 ]

Зубной зуб большинства реинчоцефалов, включая туатару, описывается как акродонт , который связан с состоянием зубов, прикрепленных к гребню челюстной кости, отсутствует замену зубов и обширный рост костей, слияние зубов к челюстям, в результате Граница между зубами и костью трудно различить. Это отличается от состояния, обнаруженного у большинства ящериц (кроме акродонтанов ), которые имеют зубы плевродонта , которые прикреплены к полке на стороне, обращенной внутрь челюсти, и заменяются на протяжении всей жизни. У зубов туатары нет корней, хотя зубы некоторых других ренчоцефалов обладают корнями. [ 22 ] Акродонт, по -видимому, является полученным характером ренчоцефалов, не обнаруженных у более примитивных лепидозаврафоров. [ 20 ] Наиболее примитивные ринчоцефальцы имеют либо зубы плевродонта, либо комбинацию как спереди, так и задних зубов Pleurodont. [ 22 ] [ 18 ] Некоторые ринчоцефальцы отличаются от этих условий, при этом ankylosphenodon имеет поверхностно акродонтные зубы, которые проходят глубоко в челюстной кости и сливаются с костью у основания гнезда (Ankylothecodont). [ 22 ] Во многих производных спенодонте, предчелюстные зубы в передней части верхней челюсти объединены в большую зубчатую структуру. [ 23 ]
Ринчоцефальцы обладают небным зубным зубом (зубы, присутствующие на костях крыши рта). Пятнистые зубы присутствуют в тетраподах, но были потеряны во многих группах. У самых ранних ринчоцефальцев были зубы на костях Палатина , Вомера и Птеригоида , хотя в некоторых группах витает во рва и/или птеригоидных зубах, включая живую туатару, у которого есть только палатинские зубы. [ 24 ] Отличительным персонажем, обнаруженным во всех ренчоцефалиях, является увеличение зубного ряда, присутствующего на нехватимых костях. В то время как у других ренчоцефальцев палатинский зубной ряд наклонен к зубам верхней челюсти , у членов Sphenodontinae (включая Tuatara) и Eilenodontinae, он ориентирован параллельно верхней челюсти. В этих группах, во время укуса зубы зубных сил в нижней части челюсти между верхнечелюстными и лаконичными зубными рядами. Эта договоренность, которая является уникальной среди амниотов, разрешает изгиб продуктов питания с тремя точками, [ 25 ] и в сочетании с пропалентным движением (заднее и вперед движение нижней челюсти) позволяет укусить сдвиг. [ 24 ] [ 26 ]

Размер тела ренчоцефалов сильно варьируется. Туатара имеет среднюю общую длину 34,8 и 42,7 сантиметра (13,7 и 16,8 дюйма) для женщин и мужчин соответственно. [ 27 ] Clevosaurus SectumSemper имеет предполагаемую общую длину 12 сантиметров (4,7 дюйма), [ 28 ] В то время как самый большой известный наземный спенодонтиан, Priosphenodon Avelasi имеет предполагаемую общую длину чуть более 100 сантиметров (39 дюймов). [ 29 ] Водные плеврозавры достигли длины до 150 сантиметров (59 дюймов). [ 30 ]
Туатара имеет среди самых высоких возрастов сексуальной зрелости среди рептилий, [ 31 ] в возрасте от 9 до 13 лет, [ 32 ] и имеет высокую долговечность по сравнению с ящерицами аналогичного размера, [ 31 ] С дикими людьми, вероятно, достигнут 70 лет, и, возможно, старше 100 лет в возрасте. [ 33 ] Такое позднее начало сексуальной зрелости и долговечности, возможно, не было типичным для вымерших ренчоцефалов. [ 30 ] [ 34 ]
Классификация
[ редактировать ]
В то время как группировка Rhynchocephalia хорошо поддерживается, отношения многих таксонов друг с другом неясны, существенно различающиеся между исследованиями. [ 35 ] В современной кладистике кладийская спенодонтия включает в себя всех ренчоцефалов, кроме Виртбергии , а также Gephyrosaurus и других гефирозавридов . Гефиросавриды были обнаружены как более тесно связанные с скваматами в некоторых анализах. [ 36 ] [ 18 ] В 2018 году были определены две основные клады в спенодонтиях, InfraRoder эсфенодонтия , который определяется наименее инклюзивной кладкой, содержащей полисфенодон , Clevosaurus Hudsoni и Sphenodon , который поддерживается наличием синапоморфи трех Зубы зубов или верхней челюсти, зубцы перед добычами объединяются в структуру, похожие на зубило, а не палатиновые зубы сводятся к одному зубному ряду, с наличием дополнительного изолированного зуба. нефронтальной клады Несфенодонтия определяется как наиболее инклюзивная клада, содержащая спенодон , но не Clevosaurus Hudsoni, который подтверждается наличием шести синапоморфий, включая повышенную относительную длину антиорбитальной области (часть черепа вперед кожу гнезда), достигая от 1/4 до 1/3 от общей длины черепа, задний (задний) край париетальной кости лишь слегка изогнут внутрь, теменное отверстие обнаруживается на том же уровне или вперед от передняя граница супратропорального фенестра (отверстие черепа), лаконичные зубы дополнительно уменьшаются от состояния у эйсфенодонтианцев до одного бокового зубного ряда, количество птеригоидных зубных рядов уменьшается до одного или нет, а задние границы Иший . характеризуется отличительным процессом [ 23 ] В 2021 году была определена клада -акросфенодонтия , которая менее инклюзивна, чем спенодонтия и более инклюзивная, чем эусфенодонтия, и включает в себя все спенодонтианцы с полностью акродонтом, исключая базальные частично акродонтные спенодонтианцы. [ 37 ] В 2022 году была определена вымершая кладская лепторгинхия , в том числе различные неосфенодонтианцы, по крайней мере, некоторые из которых были адаптированы водно, характеризованные удлинением четвертого метакарпала, наличием заднего процесса на исиме и антарбитальной областью черепа. находится от трети до четверти от общей длины черепа. [ 19 ] Клада Opisthodontia использовалась для группировки всех спенодонтианцев, более тесно связанных с приосфенодоном (член Eilenodontinae ), чем с Sphenodon. [ 38 ] Не все исследования используют эту кладу, так как некоторые исследования обнаружили, что объем клады идентична Eilenodontinae. [ 19 ]
Семейство Sphenodontidae использовалась для включения туатары и ее ближайших родственников в Rhynchocephalia. Однако в группе не хватало официального определения, причем включенные таксоны существенно различаются между анализами. [ 36 ] Ближайшие родственники туатары помещаются в клад -спенодонтину , которые характеризуются полностью закрытым височным баром. [ 17 ]
Ниже приведена кладограмма Rhynchocephaly после Demar et al. 2022 (на основе максимальной экономии , обратите внимание, что кладограмма падает в политомию при байесовском анализе ): [ 19 ]
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Кладограмма после Simoes et al. 2022 (на основе байесовского анализа вывода ): [ 17 ]
| Sphenodontia |
| |||||||||||||||||||||||||||||||||||||||||||||||||||
Клады и роды
[ редактировать ]- † Вирмибергия
- † Gephyrosauridae ?
- † Бхаратагама ?
- Sphenodontia Williston 1925
- † Diphydontosaurus
- † Микроменодон
- † Paleollanosaurus ?
- † Пелецимала
- † Whitakersaurus
- † Parvosaurus
- Acrosphenodontia Chambi-Trowell et al., 2021
- † Godavarisaurus
- † Planocephalosaurus
- † Террет
- † Sphenocondor
- † Rebbanasaurus
- Eusphenodontia Herrera-Flores et al. 2018
- † Opisthiamus
- † Брэйхинодон
- † Колобопы ?
- † Lanceirosphenodon
- † Полисфенодон
- Clevosauridae
- † Clevosaurus
- † Брахирхинодон ?
- † Polysphenodon ?
- † Сигмала
- † Микросфенодон
- † Trullidens
- Neosphenodontia errera-flores et al. 2018
- † Lamarquesaurus
- † Pamizinsaurus
- † Тингитана
- † ankylosphenodon
- † Derasmosaurus
- † Opisthodontia
- Sphenodontidae
- † Leporhyncia demar, Jones & Carrano, 2022
- † Лептозавр
- † Каллимодон
- † Гомеозавр
- † Vadasaurus
- † Sapheosauridae
- † Pleurosauridae
-
 Реконструкция черепа Gephyrosaurus вероятный базальный ринчоцефан
Реконструкция черепа Gephyrosaurus вероятный базальный ринчоцефан -
 Реконструкция черепа дипидонтозавра. Базальный член Sphenodontia
Реконструкция черепа дипидонтозавра. Базальный член Sphenodontia -
 Реконструкция черепов Clevosaurus Hudsoni (A) и Clevosaurus Cambrica (B)
Реконструкция черепов Clevosaurus Hudsoni (A) и Clevosaurus Cambrica (B) -
 Череп базального эусфенодонтса Описттиамимус
Череп базального эусфенодонтса Описттиамимус -
 Череп Сфинотитан , ранний член Eilenodontinae
Череп Сфинотитан , ранний член Eilenodontinae -
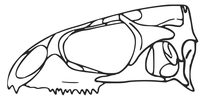 Реконструкция черепа Наваджосенодона , раннего члена Sphenodontinae
Реконструкция черепа Наваджосенодона , раннего члена Sphenodontinae -
 Реконструкция черепа эйленодонтинского приосфенодона
Реконструкция черепа эйленодонтинского приосфенодона -
 Череп неосфенодонтского вадасавра
Череп неосфенодонтского вадасавра -
 Череп Pleurosaurus
Череп Pleurosaurus -
 Схема черепа туатары ( Sphenodon Punctatus )
Схема черепа туатары ( Sphenodon Punctatus )










Экология
[ редактировать ]
Окаменевает репутацию ринчоцефалов, демонстрируя, что они были разнообразной группой, которая использовала широкий спектр экологических ниш. [ 3 ] [ 25 ] Ранние ренчоцефалы обладают небольшими яйцевидными зубами, предназначенными для пирсинга, и, вероятно, были насекомыми . [ 39 ] Как и современная туатара, вымершие члены Sphenodontinae, вероятно, были универсалами с хищной/насекомоядтной диетой. [ 40 ] Среди наиболее разных ринчоцефалов-плеврозавры , была адаптирована для морской жизни, с удлиненными змеями, похожими Европы, которая известные из юрского периода Другие ренчоцефалии. Считается, что плеорозавры были писци -распадами (потребляя рыбу). [ 30 ] Предполагается, что несколько других линий ренчоцефалов обладали полуаколичными привычками. [ 41 ] Считается, что eilenodontines были травоядными, с батареями с широкими зубами с толстой эмалью, используемой для обработки растительного материала. [ 42 ] Сафеозавриды из позднего юрского периода Европы , , такие как Oenosaurus и Sapheosaurus обладают широкими зубными пластинами, уникальными среди тетрапод, и, как полагают, были дурофаговыми , причем зубные пластины используются для раздавливания организмов с твердыми оболочкой. [ 43 ] [ 36 ] Предполагается, что Sphenovipera из юрского периода Мексики была ядовитой , основываясь на наличии канавок на двух увеличенных зубах в передней части нижней челюсти [ 44 ] хотя это интерпретация была поставлена под сомнение другими авторами. [ 44 ] Тело Pamizinsaurus из раннего мела Мексики было покрыто остеоскатами , аналогично таковым у Helodermatid ящериц , таких как Gila Monster , который является уникальным среди известных спхенодонтов, которые, вероятно, служили для защиты его от хищников. [ 45 ]
Эволюционная история
[ редактировать ]
По оценкам, сроки, когда Rhynchocephalia расходится от Squamata, оспаривается. Старые оценки помещают дивергенцию между средним пермским и самым ранним триасом, около 270-252 миллионов лет назад, [ 36 ] в то время как другие авторы устанавливают более молодую дату около 242 миллионов лет назад. [ 1 ] Самыми старыми известными останками ренчоцефалов являются остатки Виртбергии, известные из формации Эрфурта возле Велберга на юге Германии, начиная с ладинианской стадии среднего триаса , около 238-240 миллионов лет, которая также является самой примитивной ринчоцефальной. [ 18 ] Rhynchocephalians подвергся значительной диверсификации во время позднего триаса, [ 3 ] и достиг всемирного распределения по всему Пангее к концу триаса, с поздней триасой - раннего юрского периода родной родом Clevosaurus с 10 видами по всей Азии, Африке, Европе, Северной и Южной Америке. [ 46 ] Самыми ранними ринчоцефальцами были мелкие животные, но к позднему триасу группа развила широкий спектр размеров тела. [ 47 ] Во время юрского периода ринчоцефалии были доминирующей группой небольших рептилий во всем мире, [ 48 ] достигнув вершины морфологического разнообразия в этот период, включая специализированные травоядные и водные формы. [ 3 ] Единственной записью ренчоцефалов из Азии (исключая индийский субконтинент , который не был частью Азии во время мезозойского), являются неопределенные остатки Clevosaurus из ранней юрской ( синемурийской ) старой формирования Луфенга в Юньнане , Китай. Ринчоцефалии заметно отсутствуют в молодых населенных пунктах региона, несмотря на наличие благоприятных условий сохранения. [ 49 ] Rhynchocephalians оставались разнообразными в позднем юре, [ 50 ] и были более распространены, чем ящерицы во время поздней юры в Северной Америке. [ 48 ]
Разнообразие Rhynchocephalian сократилось во время раннего мела , исчезая из Северной Америки и Европы после окончания эпохи, [ 51 ] и отсутствовали в Северной Африке [ 52 ] и северная Южная Америка [ 53 ] к раннему поздному мелу . Причина снижения ринчоцефалии остается неясной, но часто предполагается, что она связана с конкуренцией с продвинутыми ящерицами и млекопитающими. [ 54 ] Похоже, что они оставались распространенными в юге Южной Америки во время позднего мела, где ящерицы оставались редкими, а их по -прежнему превосходили наземные ящерицы в этом регионе в 200 лет. [ 52 ] Поздние меловые спенодонтианцы южноамериканцев представлены как Eilenodontinae, так и Sphenodontidae (включая Sphenodontinae). [ 55 ] Неопределенный ринчоцефайец известен из частичной нижней челюсти вылупления из последних меловых или, возможно, самых ранних палеоценовых межтрапейских слоев , в том, что тогда было изолированной сухопутной массией островной Индии , которая, по -видимому, является акросфенодонтом, возможно, связанным с Годаваризавром из юридического Индия. [ 50 ] Самыми молодыми несомненными останками ренчоцефалов за пределами Новой Зеландии являются остатки спенодонтид Кавасфенодон Пелигенсис из раннего палеоцена ( Даниан ) из Патагонии , вскоре после события критического ипалеогенового вымирания . [ 56 ] известны из раннего миоцена (19–16 миллионов лет назад) Неопределенные фрагменты с сфенодонтиной челюсти с зубами . Маловероятно, что предки туатары прибыли в Новую Зеландию через океаническую рассеивание , и считается, что они уже присутствовали в Новой Зеландии, когда она отделялась от Антарктиды между 80 и 66 миллионами лет назад. [ 54 ]
Ссылки
[ редактировать ]- ^ Jump up to: а беременный Джонс я, Андерсон К.Л., Хипсли К.А., Мюллер Дж., Эванс С.Е., Шох Р.Р. (сентябрь 2013 г.). «Интеграция молекул и новых окаменелостей подтверждает триасовое происхождение для лепидозаурии (ящерицы, змеи и туатара)» . BMC Эволюционная биология . 13 (1): 208. Bibcode : 2013bmcee..13..208j . doi : 10.1186/1471-2148-13-208 . PMC 4016551 . PMID 24063680 .
- ^ Рейносо В.Х. (2000). «Необычный водный спенодонтиан (рептилия: Диапсида) из формации Тлаюа (Альбиан), Центральная Мексика». Журнал палеонтологии . 74 (1): 133–148. Bibcode : 2000jpal ... 74..133r . doi : 10.1017/s0022336000031310 . S2CID 232346834 .
- ^ Jump up to: а беременный в дюймовый Herrera-Flores JA, Stubbs TL, Benton MJ (2017). «Макроэволюционные узоры в Rhynchocephalia: Tuatara (Sphenodon Punctatus) живым ископаемым?» Полем Палеонтология . 60 (3): 319–328. Bibcode : 2017Palgy..60..319H . doi : 10.1111/pala.12284 . ISSN 1475-4983 .
- ^ Jump up to: а беременный Fraser N, Sues HD, eds. (1994). «Филогения» в тени динозавров: ранние мезозойские тетраподы . Издательство Кембриджского университета . ISBN 0-521-45242-2 .
- ^ Эванс С.Е. , Прасад Г.В., Манхас Б.К. (ноябрь 2001 г.). «Rhynchocephalians (Diapsida: Lepidosauria) из формирования юрского кота в Индии» . Зоологический журнал Линневого общества . 133 (3): 309–34. doi : 10.1111/j.1096-3642.2001.tb00629.x .
- ^ "Спенодон" . Dictionary.com Unabridged (V 1.1) . Random House , Inc. Получено 8 января 2007 года .
- ^ Эванс С.Е., Борсук-Биалайника М. (2009). «Небольшая лепидозавраморфная рептилия из раннего триаса Польши» (PDF) . Paleontologica Polonica . 65 : 179–202.
- ^ Gauthier, J., Estes, R. & De Queiroz, K. (1988). Филогенетический анализ лепидозавраморфы. В филогенетических отношениях семейств ящериц (ред. Р. Эстес и Г. Прегилл), с. 15–98. Издательство Стэнфордского университета, Стэнфорд.
- ^ «Ринчоцефалии» . Университетский колледж Лондон . Получено 2022-08-09 .
- ^ Денди А (1911). «VI. О структуре, развитии и морфологической интерпретации органов шишки и прилегающих частей мозга в туатаре (Sphenodon Punctatus)» . Философские транзакции Королевского общества б . 201 (274–281): 227–331. doi : 10.1098/rstb.1911.0006 . ISSN 0264-3960 .
- ^ Смит К.Т., Бхуллар Б.А., Келер Г., Хаберсетцер Дж (2 апреля 2018 года). «Единственный известный челюстный позвоночный с четырьмя глазами и бауплан комплекса шишковидной т. Д.» . Текущая биология . 28 (7): 1101–1107.e2. Bibcode : 2018cbio ... 28E1101S . doi : 10.1016/j.cub.2018.02.021 . ISSN 0960-9822 . PMID 29614279 .
- ^ Jump up to: а беременный Benoit J, Abdala F, Manger P, Rubidge B (2016). «Шестой смысл у предшественников млекопитающих: изменчивость теменного отверстия и эволюция пинецкового глаза у южноафриканской пермо-триасик-эвриодонтской терапиды» . Acta Palaeontologica Polonica . doi : 10.4202/app.00219.2015 .
- ^ Полина-Карабаджал А., Хименес-Хуидбро П., Тривиньо Л.Н., Стэнли Э.Л., Захер Х.Д., Даза Д.Д. (2023), Дозо М.Т., Паулина-Карабиджал А., Макрини Т.Е., Уолш С. (ред.), «Взгляд к невронауниуму Живой и вымершей Лепидозаурией » , Палинерология Амниотов , Cham: Springer International Publishing, с. 123–177, doi : 10.1007/978-3-031-13983-3_5 , ISBN 978-3-031-13982-6 Получено 2023-12-08
- ^ Vitt LJ, Caldwell JP (2014). «Глава 20: Rhynchocephalians (Sphenodontids)». Герпетология . Elsevier. С. 553–554. doi : 10.1016/b978-0-12-386919-7.00020-4 . ISBN 978-0-12-386919-7 .
- ^ Sanger TJ, Gredler ML, Cohn MJ (октябрь 2015 г.). «Воскресение эмбрионов туатары, Спенодонского пропачка, для разрешения эволюции фаллоса позвоночных» . Биологические письма . 11 (10): 20150694. DOI : 10.1098/rsbl.2015.0694 . PMC 4650183 . PMID 26510679 .
- ^ Эванс С.Е., Джонс я (2010). «Происхождение, ранняя история и диверсификация лепидозавраморфных рептилий». Новые аспекты мезозойского биоразнообразия . Заметки лекций в науках о Земле. Тол. 132. Берлин, Гейдельберг: Спрингер Берлин Хейдельберг. С. 27–44. Bibcode : 2010lnes..132 ... 27e . doi : 10.1007/978-3-642-10311-7_2 . ISBN 978-3-642-10310-0 .
- ^ Jump up to: а беременный в Simões TR, Kinney-Broderick G, Pierce Se (март 2022 г.). «Исключительно сохранившаяся спенодоноподобная спенодонтиан выявляет глубокое сохранение скелета и онтогенезы Туатары» . Биология связи . 5 (1): 195. doi : 10.1038/s42003-022-03144-y . PMC 8894340 . PMID 35241764 .
- ^ Jump up to: а беременный в дюймовый и Суды HD, Schoch RR (2023-11-07). «Самая старая известная рептилия с Ринчоцефалией из среднего триаса (ладиниан) Германии и ее филогенетическое положение среди лепидозавраморфы» . Анатомическая запись . 307 (4): 776–790. doi : 10.1002/ar.25339 . ISSN 1932-8486 . PMID 37937325 .
- ^ Jump up to: а беременный в дюймовый DeMar DG, Jones ME, Carrano MT (2022-12-31). «Почти полный скелет нового Eusphenodontian из формации Верхней Юры Моррисон, штат Вайоминг, США, дает представление о эволюции и разнообразии Rhynchocephalia (Reptilia: Lepidosauria)» . Журнал систематической палеонтологии . 20 (1): 1–64. doi : 10.1080/14772019.2022.2093139 . HDL : 2440/136608 . ISSN 1477-2019 . S2CID 252325953 .
- ^ Jump up to: а беременный Ford DP, Evans SE, Choiniere JN, Fernandez V, Benson RB (2021-08-25). «Переоценка загадочной диапсидной палигуаны и ранней истории lepidosauromorpha» . Труды Королевского общества B: Биологические науки . 288 (1957): 20211084. DOI : 10.1098/rspb.2021.1084 . ISSN 0962-8452 . PMC 8385343 . PMID 34428965 .
- ^ Эванс С.Е. (2016), Clack JA, Fay RR, Popper An (Eds.), «Лепидозаврское ухо: вариации на тему» , Эволюция уха позвоночных , Справочник Springer of Survey Research, vol. 59, CHAM: Springer International Publishing, стр. 245–284, doi : 10.1007/978-3-319-46661-3_9 , ISBN 978-3-319-46659-0 Получено 2024-01-08
- ^ Jump up to: а беременный в Дженкинс К.М., Джонс М.Е., Зикмунд Т., Бойде А., Даза Д.Д. (сентябрь 2017 г.). «Обзор зубной имплантации среди реинчоцефалов (чешуекрылая)». Журнал герпетологии . 51 (3): 300–306. doi : 10.1670/16-146 . ISSN 0022-1511 . S2CID 90519352 .
- ^ Jump up to: а беременный Herrera-Flores JA, Stubbs TL, Elsler A, Benton MJ (2018-04-06). «Таксономическая переоценка Clevosaurus Latidens Fraser, 1993 (Lepidosauria, Rhynchocephalia) и филогения Rhynchocephalian, основанная на экологической и байесовской выводе» . Журнал палеонтологии . 92 (4): 734–742. Bibcode : 2018Jpal ... 92..734H . doi : 10.1017/jpa.2017.136 . HDL : 1983/59126B60-16D8-46D2-B657-954693A39D4E .
- ^ Jump up to: а беременный Matsumoto R, Evans SE (январь 2017 г.). «Пятниковая зубная зуба тетрапод и его функциональная значимость» . Журнал анатомии . 230 (1): 47–65. doi : 10.1111/joa.12534 . PMC 5192890 . PMID 27542892 .
- ^ Jump up to: а беременный Джонс меня (август 2008 г.). «Форма черепа и стратегия кормления в Спенодоне и других реинчоцефалии (Diapsida: Lepidosauria)» . Журнал морфологии . 269 (8): 945–966. doi : 10.1002/jmor.10634 . PMID 18512698 .
- ^ Джонс я, О'Гиггин П., Фаган М.Дж., Эванс С.Е., Кертис Н. (июль 2012 г.). «Механика сдвига и влияние гибкого симфиза при оральной обработке пищи в Sphenodon (Lepidosauria: Rhynchocephalia)» . Анатомическая запись . 295 (7): 1075–1091. doi : 10.1002/ar.22487 . PMID 22644955 .
- ^ Эррел А., Мур Дж. А., Бредевег Э.М., Нельсон Нью-Джерси (2010-05-19). «Сексуальный диморфизм, размер тела, сила укуса и успех спаривания мужского пола в Туатаре: половой диморфизм в Туатаре» . Биологический журнал Линневого общества . 100 (2): 287–292. doi : 10.1111/j.1095-8312.2010.01433.x .
- ^ Keeble E, Whiteside DI, Benton MJ (апрель 2018 г.). «Земная фауна поздней триасовой брюки-я-Финнон Карьер, Южный Уэльс, Великобритания и новый вид Clevosaurus (Lepidosauria: Rhynchocephalia)» . Материалы Ассоциации геологов . 129 (2): 99–119. Bibcode : 2018prga..129 ... 99K . doi : 10.1016/j.pgeola.2017.11.001 . HDL : 1983/5AFDC677-3EA0-4519-813D-6052EF8370EC .
- ^ Apesteguía S, Novas Fe (9 октября 2003 г.). «Большой меловой спенодонтиан из Патагонии дает представление о эволюции чешуекрылых в Гондване». Природа . 425 (6958): 609–612. Bibcode : 2003natur.425..609a . doi : 10.1038/nature01995 . PMID 14534584 . S2CID 4425130 .
- ^ Jump up to: а беременный в Klein N, Scheyer TM (февраль 2017 г.). «Микроанатомия и история жизни у палеоплеурозавра (Rhynchocephalia: Pleurosauridae) из раннего юрского периода Германии». Умирайте натуральный . 104 (1–2): 4. Bibcode : 2017scina.104 .... 4K . doi : 10.1007/s00114-016-1427-3 . PMID 28005148 . S2CID 27133670 .
- ^ Jump up to: а беременный Hallmann K, Griebeler EM (июнь 2018 г.). «Исследование различий в масштабировании черт истории жизни с массой тела в рептилиях и между амниотами» . Экология и эволюция . 8 (11): 5480–5494. Bibcode : 2018ecoev ... 8.5480h . doi : 10.1002/ECE3.4069 . ISSN 2045-7758 . PMC 6010814 . PMID 29938067 .
- ^ Newman DG (июль 1988 г.). «Свидетельство о хищничестве на молодой туатаре, Спенодон Пурнат , Киоре, Раттус Эсуланс , на острове леди Алиса». Новая Зеландия Журнал зоологии . 15 (3): 443–446. doi : 10.1080/03014223.1988.10422973 . ISSN 0301-4223 .
- ^ Нельсон Н.Дж., Келл С.Н., Пледер С., Догерти Ч. (май 2002). «Соотношение пола, смещенное мужчина, в небольшой популяции туатары» . Журнал биогеографии . 29 (5–6): 633–640. Bibcode : 2002jbiog..29..633n . doi : 10.1046/j.1365-2699.2002.00712.x . ISSN 0305-0270 .
- ^ Cavasín S, Cerda I, Apesteguia S (2024). «Костяная микроструктура Priosphenodon avesi (Rhynchocephalia: spherontia): палеобиологические последствия» . Acta palaeontologica policica . 69 Doi : 10.4202/app.01071.2023 .
- ^ Ромо де Вивар П.Р., Мартинелли А.Г., Шмальц Хсиу А, Соареш М.Б. (2020-07-02). «Новый Rhynchocephalian из позднего триаса Южной Бразилии усиливает эсфенодонтское разнообразие». Журнал систематической палеонтологии . 18 (13): 1103–1126. Bibcode : 2020jspal..18.1103r . doi : 10.1080/14772019.2020.1732488 . ISSN 1477-2019 . S2CID 216226211 .
- ^ Jump up to: а беременный в дюймовый Simões TR, Caldwell MW, Pierce SE (декабрь 2020 г.). «Спенодонтская филогения и влияние выбора модели в байесовских морфологических часах оценок времени дивергенции и эволюционных показателей» . BMC Biology . 18 (1): 191. DOI : 10.1186/S12915-020-00901-5 . PMC 7720557 . PMID 33287835 .
- ^ Chambi-Trowell SA, Martinelli AG, Whiteside DI, Vivar PR, Soares MB, Schultz CL, et al. (2021-06-03). «Разнообразие триасовых южноамериканских спенодонтианцев: новая базальная форма, клевозавры и пересмотр ринчоцефальной филогения» . Журнал систематической палеонтологии . 19 (11): 787–820. Bibcode : 2021jspal..19..787c . doi : 10.1080/14772019.2021.1976292 . HDL : 1983/AF14AFFC-A26E-426B-83CA-E1833E355882 . ISSN 1477-2019 . S2CID 240487298 .
- ^ Apesteguía S, Novas Fe (2003-10-09). «Большой меловой спенодонтиан из Патагонии дает представление о эволюции чешуекрылых в Гондване» . Природа . 425 (6958): 609–612. Bibcode : 2003natur.425..609a . doi : 10.1038/nature01995 . ISSN 0028-0836 . PMID 14534584 .
- ^ Джонс я (2009). Koppe T, Meyer G, Alt KW, Brook A (ред.). «Форма зубного зуба у Спенодона и его окаменелостей (Diapsida: Lepidosauria: Rhynchocephalia)». Границы пероральной биологии . 13 Базель: Каргер: 9–15. doi : 10.1159/000242382 . ISBN 978-3-8055-9229-1 Полем PMID 19828962 .
- ^ Villa A, Montie R, Röper M, Rothgaenger M, Rauhut OW (2021-05-03). «Sphenofontis Velserae Gen. Et sp. Nov., Новый Rhynchocephalian от покойного юрского периода Брунна (Solnhofen Archipelago, Южная Германия)» . ПЕРЕЙ . 9 : E11363. doi : 10.7717/peerj.11363 . ISSN 2167-8359 . PMC 8101455 . PMID 33987027 .
- ^ Bever GS, Norell MA (ноябрь 2017 г.). «Новый Rhynchocephalian (Reptilia: Lepidosauria) из покойной юры Солнхофена (Германия) и происхождения морских Pleurosauridae» . Королевское общество открыто наука . 4 (11): 170570. DOI : 10.1098/rsos.170570 . PMC 5717629 . PMID 29291055 .
- ^ Джонс я, Лукас П.В., Такер А.С., Уотсон А.П., Сертич Дж.Дж., Фостер Дж.Р. и др. (Июнь 2018 г.). «Сканирование нейтронов выявляет неожиданную сложность в толщине эмали травяной рептилии юрского периода» . Журнал Королевского общества, интерфейс . 15 (143): 20180039. DOI : 10.1098/RSIF.2018.0039 . PMC 6030635 . PMID 29899156 .
- ^ Раухут Оу, Хейнг А.М., Лопес-Арбарелло А., Хеккер А. (2012). Фарке А.А. (ред.). «Новый Rhynchocephalian из позднего юрского периода Германии с уникальным зубным зубным промышленностью» . Plos один . 7 (10): E46839. BIBCODE : 2012PLOSO ... 746839R . doi : 10.1371/journal.pone.0046839 . PMC 3485277 . PMID 23118861 .
- ^ Jump up to: а беременный Folinsbee KE, Müller J, Reisz RR (2007-06-12). «Канавки для собак: морфология, функция и отношение к яду » Журнал палеонтологии позвоночных 27 (2): 547–551. Doi : 10.1671/0272-4634 (2007) 27 [547: cgmfar] 2.0.co ; ISSN 0272-4
- ^ Рейносо В.Х. (1997-04-16). «А" бисера "спенодонтиан (Diapsida: Lepidosauria) из раннего мела Центральной Мексики» . Журнал палеонтологии позвоночных . 17 (1): 52–59. Bibcode : 1997jvpal..17 ... 52r . doi : 10.1080/02724634.1997.10010953 . ISSN 0272-4634 .
- ^ Chambi-Trowell SA, Whiteside DI, Benton MJ, Rayfield EJ (ноябрь 2020 г.). Lautenschlager S (ред.). «Биомеханические свойства челюстей двух видов Clevosaurus и повторный анализ Rhynchocephalian Dentary Morphospace». Палеонтология . 63 (6): 919–939. Bibcode : 2020palgy..63..919c . doi : 10.1111/pala.12493 . S2CID 220902843 .
- ^ Herrera-Flores JA, Elsler A, Stubbs TL, Benton MJ (2021). «Медленные и быстрые эволюционные показатели в истории чешуекрылых». Палеонтология . 65 doi : 10.1111/pala.12579 . ISSN 1475-4983 . S2CID 244019684 .
- ^ Jump up to: а беременный Brownstein CD, Meyer DL, Fabbri M, Bhular BA, Gauthhier JA (2022-11-29). «Эволюционное происхождение длительного существующего скваматного излучения » Природная связь 13 (1): Bibcode : 2022natco..13.7087b 7087. Doi : 10.1038/ s41467-022-34217-5 ISSN 2041-1 PMC 9708687 36446761PMID
- ^ Джонс я (2006). «Ранняя юрская эвозавры из Китая (Diapsida: Lepidosauria)» . Музей естественной истории и науки Нью -Мексико . 37 : 548–562.
- ^ Jump up to: а беременный Anantharaman S, Demar DG, Sivakumar R, Dassarma DC, Wilson Mantilla GP, Wilson Mantilla JA (2022-06-30). «Первый Rhynchocephalian (Reptilia, Lepidosauria) из мела -палеогена Индии» . Журнал палеонтологии позвоночных . 42 (1): E2118059. Bibcode : 2022jvpal..42e8059a . doi : 10.1080/02724634.2022.2118059 . ISSN 0272-4634 . S2CID 252558728 .
- ^ Cleary TJ, Benson RB, Evans SE, Barrett PM (март 2018 г.). «Разнообразие лепидозавра в мезозозой-палеогене: потенциальная роль отбора проб и экологических драйверов» . Королевское общество открыто наука . 5 (3): 171830. Bibcode : 2018rsos .... 571830c . doi : 10.1098/rsos.171830 . PMC 5882712 . PMID 29657788 .
- ^ Jump up to: а беременный Apesteguía S, Daza JD, Simões TR, Rage JC (сентябрь 2016 г.). «Первая игуанская ящерица из мезозойского Африки» . Королевское общество открыто наука . 3 (9): 160462. Bibcode : 2016rsos .... 360462a . doi : 10.1098/rsos.160462 . PMC 5043327 . PMID 27703708 .
- ^ Simões TR, Wilner E, Caldwell MW, Weinschütz LC, Kellner AW (август 2015 г.). «Ящерица Acrodontan в меловом фоне Бразилии пересматривает раннюю эволюцию ящерицы в Гондване» . Природная связь . 6 (1): 8149. Bibcode : 2015natco ... 6.8149s . doi : 10.1038/ncomms9149 . PMC 4560825 . PMID 26306778 .
- ^ Jump up to: а беременный Jones Me, Tennyson AJ, Worthy JP, Evans SE, Worthy Th (апрель 2009 г.). «Спенодонтин (Rhynchocephalia) из миоцена Новой Зеландии и палеобиогеографии туатары (Спенодон)» . Труды Королевского общества B: Биологические науки . 276 (1660): 1385–90. doi : 10.1098/rspb.2008.1785 . PMC 2660973 . PMID 19203920 .
- ^ Agnolín FL, Aranciaga Rolando AM, Chimento NR, Novas Fe (октябрь 2023 г.). «Новая небольшая рептилий остается от позднего мела из Патагонии, увеличивая морфологическое разнообразие спенодонтидов (лепидозавриат)» . Материалы Ассоциации геологов . 135 : 36–44. doi : 10.1016/j.pgeola.2023.09.007 .
- ^ Apesteguía S, Gómez Ro, Rougier GW (октябрь 2014). «Самый молодой южноамериканский ринчоцефалан, выживший вымирание K/PG» . Труды Королевского общества B: Биологические науки . 281 (1792): 20140811. DOI : 10.1098/rspb.2014.0811 . PMC 4150314 . PMID 25143041 .
Дальнейшее чтение
[ редактировать ]- Apesteguia S, Rougier GW (2007). «Поздний кампанский мельчайшая верхняя челюсть из Северной Патагонии» (PDF) . Американский музей новичок (3581): 1–12. doi : 10.1206/0003-0082 (2007) 3581 [1: ALCSMF] 2.0.co; 2 . Получено 2019-03-30 .
- Daugherty CH, Cree A, Hay JM, Thompson MB (1990). «Заброшенная таксономия и продолжающиеся исчезновения туатары ( Спенодон )». Природа . 347 (6289): 177–179. Bibcode : 1990natur.347..177d . doi : 10.1038/347177A0 . S2CID 4342765 .
- Эванс SE (ноябрь 2003 г.). «У ног динозавров: ранняя история и радиация ящериц» (PDF) . Биологические обзоры Кембриджского философского общества . 78 (4): 513–51. doi : 10.1017/s1464793103006134 . PMID 14700390 . S2CID 4845536 .
- Gemmell NJ, Rutherford K, Prost S, Tollis M, Winter D, Macey JR, et al. (Август 2020). «Геном Туатары раскрывает древние черты эволюции амниота» . Природа . 584 (7821): 403–409. doi : 10.1038/s41586-020-2561-9 . PMC 7116210 . PMID 32760000 .
Внешние ссылки
[ редактировать ] СМИ, связанные с Sphenodontia в Wikimedia Commons
СМИ, связанные с Sphenodontia в Wikimedia Commons  Данные, связанные с Rhynchocephalia в Wikispecies
Данные, связанные с Rhynchocephalia в Wikispecies
Основные существующие рептилий клады |
|---|
- Rhynchocephalia
- Таксоны названы Альбертом Гюнтером
- Тетрапод заказы
- Существующие первые выступления среднего триаса
- Ladinian первые выступления
- Средние триасовые таксономические заказы
- Поздние триасовые таксономические заказы
- Ранние юрские таксономические заказы
- Таксономические заказы среднего юрского периода
- Таксономические заказы на поздние юры
- Ранние меловые таксономические заказы
- Поздние меловые таксономические заказы
- Палеокеновые таксономические заказы
- Эоценовые таксономические заказы
- Олигоценовые таксономические заказы
- Миоценовые таксономические заказы
- Таксономические заказы плиоцена
- Плейстоценовые таксономические заказы
- Холоценовые таксономические заказы