Серый мышиный лемур
| Серый мышиный лемур | |
|---|---|
 | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Приматы |
| Подотряд: | Стрепсиррини |
| Семья: | хейрогалеиды |
| Род: | Микроцебус |
| Разновидность: | М. муринус |
| Биномиальное имя | |
| Микроцебус муринус | |
 | |
| Распространение Microcebus murinus [1] | |
| Синонимы [3] [4] | |
| |

Серый мышиный лемур ( Microcebus murinus ), серый мышиный лемур или малый мышиный лемур — небольшой лемур , разновидность стрепсирриновых приматов , встречающийся только на острове Мадагаскар . При весе от 58 до 67 граммов (от 2,0 до 2,4 унции) это самый крупный из мышиных лемуров ( род Microcebus ), группы, в которую входят самые маленькие приматы в мире. Вид назван в честь мышиного размера и окраски и известен на местном уровне (на малагасийском языке ) как циди , койцики , титиливаха , пондики и вакиандри . Серый мышиный лемур и все другие мышиные лемуры считаются загадочными видами , поскольку по внешнему виду они практически неотличимы друг от друга. По этой причине серый мышиный лемур на протяжении десятилетий считался единственным видом мышиного лемура, пока недавние исследования не начали различать эти виды.
Как и все мышиные лемуры, этот вид ведет ночной и древесный образ жизни . Он очень активен, и хотя добывает пищу в одиночку, группы самцов и самок образуют спящие группы и в течение дня делят норы на деревьях. он находится в состоянии покоя, называемом оцепенением В прохладные и сухие зимние месяцы , а в некоторых случаях впадает в сезонное оцепенение (или спячку ), что необычно для приматов. Серого мышиного лемура можно встретить в нескольких типах лесов на западе и юге Мадагаскара. Его рацион состоит в основном из фруктов, насекомых, цветов и нектара. В дикой природе его естественными хищниками являются совы, змеи и эндемичные хищники-млекопитающие. Давление хищников у этого вида выше, чем у любого другого вида приматов: каждый год хищник забирает одну из четырех особей. Это уравновешивается его высокой репродуктивной способностью. используются отдельные вокализации Размножение носит сезонный характер, и для предотвращения гибридизации с видами, перекрывающими его ареал, . Беременность длится около 60 дней, и обычно рождается двое детенышей. Потомство обычно становится самостоятельным через два месяца и может воспроизводиться через год. Продолжительность репродуктивной жизни серого мышиного лемура составляет пять лет, хотя, как сообщается, особи в неволе живут до 15 лет.
Несмотря на то, что ему угрожают вырубка лесов , деградация среды обитания и отлов живых животных для торговли домашними животными , он считается одним из самых многочисленных мелких местных млекопитающих Мадагаскара. Он может переносить умеренную нехватку продовольствия, ежедневно находясь в оцепенении для сохранения энергии, но длительная нехватка продовольствия из-за изменения климата может представлять значительный риск для этого вида.
Этимология
[ редактировать ]Серый или малый мышиный лемур назван в честь своего коричневато-серого меха, мышиного размера и внешнего вида. Название рода Microcebus происходит от греческих слов mikros , что означает «маленький», и kebos , что означает «обезьяна». Латинская не версия кебоса , cebus , — распространенный суффикс, используемый для названий приматов, несмотря на то, что серый мышиный лемур — это лемур, а обезьяна . Название вида , murinus , означает «мышиный» и происходит от латинского слова mus , или «мышь», и латинского суффикса -inus , что означает «подобный». [5]
Лемур известен на малагасийском языке под несколькими именами, в зависимости от региона. Вокруг Толанаро (форт Дофин) его называют пондики [punˈdikʲ] . В северной части ареала он известен как циди [ˈtʃidʲ] . В окрестностях Морондавы его называют койцики [куиˈтʃикʲ] , титиливаха [ти̥тилива] и вакиандри [вакиˈaɳɖʐʲ] . [6] [7] Во многих случаях эти малагасийские названия также используются для других визуально неотличимых видов мышиных лемуров, обитающих в пределах его ареала. [8]
Таксономия
[ редактировать ]Как следует из названия, серый мышиный лемур — это лемур, тип приматов, принадлежащий к подотряду Strepsirrini и инфраотряду Lemuriformes. [3] В пределах Lemuriformes он принадлежит к семейству Cheirogaleidae , которое включает мышиных лемуров, карликовых лемуров , гигантских мышиных лемуров , лемуров с вилкой и волосатых ушей карликового лемура . Род мышиных лемуров Microcebus включает самых маленьких приматов в мире. [3] [6] [9] [10] Филогенетический анализ последовательностей D-петли ДНК различных видов лемуров позволяет предположить, что серый мышиный лемур может быть наиболее тесно связан с красновато-серым мышиным лемуром ( M. griseorufus ). [11]
В 1777 году английский иллюстратор Джон Фредерик Миллер включил раскрашенную вручную пластину с изображением серого мышиного лемура в свою книгу «Icones Animalium et Plantarum» и придумал биномиальное название Lemur murinus . [12] Вид был переведен в род Microcebus французским натуралистом Этьеном Жоффруа Сен-Илером в 1834 году. [13] M. murinus оставался единственным видом своего рода, а также названием, используемым для всех мышиных лемуров на Мадагаскаре, между первым крупным таксономическим пересмотром в 1931 году и обширным исследованием, в 1972 году. проведенным полевым M. rufus — тогда считавшийся подвидом — как отдельный симпатрический вид, обитавший в юго-восточной части острова. [14] После этой редакции считалось, что серый мышиный лемур представляет всех мышиных лемуров, обитавших в более засушливых частях севера, запада и юга, в то время как коричневый мышиный лемур представляет восточных мышиных лемуров тропических лесов. В последнее время научное понимание распространения и разнообразия мышиных лемуров стало намного сложнее. [9] Дополнительные полевые исследования, генетическое тестирование и последующий таксономический пересмотр на протяжении 1990-х и 2000-х годов выявили многочисленные новые виды мышиных лемуров, продемонстрировав, что этот род включает по меньшей мере 17 загадочных видов . [15] [16] [17] [18]
Анатомия и физиология
[ редактировать ]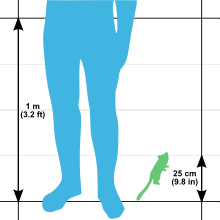
Серый мышиный лемур имеет много общих черт с другими мышиными лемурами, включая мягкий мех, длинный хвост, длинные задние конечности, спинную полосу на спине (не всегда отчетливую), короткую морду, округлый череп, выдающиеся глаза. [10] и большие, перепончатые, оттопыренные уши. [6] [7] [9] У него большие глаза и блестящий тапетум, позволяющий лучше видеть в ночное время. [9] Шерсть на спине коричневато-серая с различными красноватыми оттенками, бока от светло-серого до бежевого, а на брюшной части шерсти имеются отдельные тускло-бежевые или беловато-бежевые пятна вдоль частей живота. На округлом лице над носом и между глазами имеется бледно-белое пятно; у некоторых особей есть темные отметины на орбитах. Покрытые мехом части рук и ног грязно-белые. [6]
Серый мышиный лемур — один из самых маленьких приматов в мире, но также и самый крупный мышиный лемур. [9] Его общая длина составляет от 25 до 28 см (от 9,8 до 11,0 дюйма), длина головы и тела от 12 до 14 см (от 4,7 до 5,5 дюйма) и длина хвоста от 13 до 14,5 см (от 5,1 до 5,7 дюйма). [6] Средний вес этого вида составляет 60 г (2,1 унции). [9] с диапазонами 58 и 67 г (2,0 и 2,4 унции) [6] и 40 и 70 г (1,4 и 2,5 унции) [9] сообщил. Это меньше, чем самая маленькая обезьяна в мире, карликовая игрунка , вес которой колеблется от 85 до 140 г (от 3,0 до 4,9 унции). [10] Вес варьируется в зависимости от сезона: перед засушливым сезоном и периодами покоя представители обоих полов накапливают жировые запасы, составляющие до 35% массы тела, в хвосте и задних лапах. [9] Хвост серого мышиного лемура может увеличиваться в объеме в четыре раза во время сезона дождей, когда он накапливает жир. Даже в неволе, когда экологические ограничения не являются проблемой, мышиные лемуры демонстрируют сезонные диетические предпочтения с большим потреблением белка в период их более активной активности. [19]
Исследователи выявили различия в морфологии зубов первых и вторых коренных зубов между серым мышиным лемуром и красновато-серым мышиным лемуром. У красновато-серого мышиного лемура первый нижний коренной зуб (m1) имеет квадратную форму, а первые и вторые верхние коренные зубы (M1 и M2) имеют небольшие углубления вокруг середины заднего края. У серого мышиного лемура m1 более удлиненный и отсутствует вышеупомянутое углубление на M1 и M2. [20] Что касается общего зубного ряда , серый мышиный лемур имеет ту же зубную формулу, что и все другие члены его семейства Cheirogaleidae: 2.1.3.3 2.1.3.3 × 2 = 36. [21]
У этого вида 66 хромосом . [22] очень напоминает кариотип карликовых лемуров (род Cheirogaleus ). За исключением Х-хромосомы , все хромосомы акроцентричны (короткое плечо отсутствует или практически отсутствует). [23] его Размер генома составляет 3,12 пикограмма (пг). [22]
Экология
[ редактировать ]Как и все другие представители семейства Cheirogaleidae , серый мышиный лемур ведет ночной и древесный образ жизни . Он населяет низинные тропические сухие леса, субзасушливые терновые заросли, галерейные леса , колючие леса , восточные прибрежные леса, сухие лиственные леса , полувлажные лиственные, влажные низинные леса , переходные леса, а также вторичные леса или деградированные леса (включая плантации). на высоте до 800 м (2600 футов) над уровнем моря. [6] [7] [9] Этот вид чаще встречается во вторичных лесах, чем в девственных лесах, особенно в среде обитания кустарников и кустарников, где он занимает нишу «тонких ветвей» , ограничивая вертикальный диапазон тонкими ветвями, тонкими верхними опорами, лианами и густой листвой. [7] [9] [24] Этих лемуров обычно можно увидеть на ветвях диаметром менее 5 см (2,0 дюйма). [24] Серый мышиный лемур, как правило, предпочитает более низкие уровни леса и подлесок , где ветви и растительность густые. [9]
Во вторичном лесу это обычно наблюдается на высоте до 10 м (33 футов) над землей и на высоте от 15 до 30 м (от 50 до 100 футов) в пологе девственного леса . [7] [24] Исследования показали, что этот вид может проводить до 40% своего времени на глубине ниже 3 м (10 футов), причем 70% времени он проводит на этом уровне в конце засушливого сезона, когда растительная пища ограничена и насекомые составляют больший процент рациона. [24] Этот вид более многочисленнен в колючем лесу, например в специальном заповеднике Андохахела , чем в галерейном лесу, предпочитая более сухой прибрежный лес, тогда как коричневый мышиный лемур предпочитает внутренние тропические леса. [7]
Пределы распространения плохо изучены. Считается, что его территория простирается от реки Онилахи или озера Циманампецоца на юге до национального парка Анкарафанцика на севере. [6] [9] Есть также изолированное и разрозненное население в юго-восточной части острова, недалеко от Толанаро и национального парка Андоахела, вплоть до заповедной зоны Мандена . [6] [9] Люди, как правило, занимают небольшие участки дома площадью от 1 до 2 га (от 2,5 до 4,9 акров). [9] Серый мышиный лемур также симпатичен красновато-серому мышиному лемуру, золотисто-коричневому мышиному лемуру , мышиному лемуру мадам Берты и некоторым другим хейрогалеидным лемурам. [6] В национальном парке Анкарафантика , где он сосуществует с золотисто-коричневым мышиным лемуром, относительная плотность популяции серого мышиного лемура была самой высокой на больших высотах и в более сухой среде обитания, в то время как золотисто-коричневый мышиный лемур предпочитал обратное. [25] Другое исследование изучало сосуществование серого мышиного лемура и мышиного лемура мадам Берты и обнаружило небольшие, исключительные скопления каждого вида и высокую степень перекрытия кормовых ниш . [26]
Серый мышиный лемур может достигать высокой плотности популяции — до нескольких сотен особей на квадратный километр. [7] [9] (до 167 особей/км 2 в национальном парке Анкарафантика до 712 особей/км. 2 в национальном парке Киринди Митеа ). [1] Эта численность неоднородна и имеет тенденцию концентрироваться в «популяционных ядрах». [9] предполагая, что трудно точно оценить плотность населения при экстраполяции с небольшой территории (по данным различных исследований) на большую территорию. Трудность обнаружения особей в некоторые времена года, особенно в засушливые периоды, может еще больше усложнить задачу оценки плотности населения. [7]
Мышиные лемуры всеядны и большую часть своего рациона отдают фруктам и насекомым. Серый мышиный лемур может даже спуститься на землю, чтобы поймать добычу насекомых. [7] хотя он быстро возвращается под защитный покров подлеска, чтобы съесть добычу. [24] Нектар также входит в рацион серого мышиного лемура, что делает его потенциальным опылителем местных видов растений. [9] Известен единичный случай, когда самец поедал взрослую самку. [27]
Хищничество
[ редактировать ]

Наиболее значительными хищниками серого мышиного лемура являются мадагаскарская сова и сипуха . [6] [24] Исследования, проведенные в заповеднике Беза Махафали и национальном парке Киринди Митеа, показывают, что уровень хищничества составляет около 25% (процент популяции, съедаемый хищниками в год), что является самым высоким показателем среди всех видов приматов. Однако, учитывая высокий репродуктивный потенциал этого вида, хищничество, по-видимому, не оказывает радикального влияния на его популяции. [24] Другие хищники , такие как ястреб-тетеревятник Хенста и другие совы, также охотятся на этот вид. Змеи , такие как Ithycyphus miniatus (аборигенная колубридная змея) и мадагаскарский древесный удав , а также хищники-млекопитающие, такие как кошачий мангуст , узкополосый мангуст , фосса и домашние собаки , также известны тем, что охотятся на серых змей. мышиный лемур. [9] [24] Хищники-млекопитающие часто обнаруживают дупла деревьев, служащие гнездами, и расширяют отверстие, чтобы поймать и съесть обитателя. Это оказывает сильное избирательное давление на выбор диаметра и глубины отверстия гнезда. [24]
Из-за своего нестайного характера серый мышиный лемур, как и другие ночные лемуры, в первую очередь защищается от хищников, используя стратегии избегания. [24] групповая защита с помощью сигналов тревоги и нападения хищников . хотя у этого вида наблюдалась [28] хищников и нехищников Исследования, включающие распространение запаха фекалий среди содержащихся в неволе популяций, продемонстрировали генетическую предрасположенность к распознаванию хищников посредством обнаружения метаболитов в результате переваривания мяса. Было показано, что мышиные лемуры избегают мест, где они обычно получают вознаграждение, а также проявляют повышенную активность и поведение против хищников, когда запах фекалий хищников присутствует в этой близости, но не когда запах фекалий малагасийских нехищников был подарок. [29]
Днем, когда они наиболее уязвимы, особи укрываются в дуплах деревьев, иногда образуя гнезда. Они могут использовать от трех до девяти различных дупл на деревьях в пределах своего домашнего ареала, но отдельные люди могут использовать одну конкретную нору до пяти дней подряд. [6] [9] Мышиные лемуры, как правило, предпочитают дупла деревьев, но также обычно строят сферические конструкции из листьев. Гнезда обычно находятся в дуплах деревьев с минимальным диаметром 5 см (2,0 дюйма) и средним диаметром 13 см (5,1 дюйма), что позволяет предположить, что этот диапазон диаметров дупл деревьев может иметь решающее значение для поддержания здоровой среды обитания. [7] Серый мышиный лемур также проводит большую часть времени в густой растительности, что ограничивает его видимость и доступность для хищников. Кроме того, он имеет высокую скорость размножения, что позволяет компенсировать потери от хищников. [24]
Поведение
[ редактировать ]Серый мышиный лемур ведет ночной образ жизни, днем спит в дуплах деревьев, выстланных опадом из листьев, или в специально построенных сферических гнездах, построенных из мертвых листьев, мха и веток. [9] Ночью он обычно добывает пищу в одиночку, но днем может спать группами, состав которых зависит от пола и сезона. [7] Дуплами деревьев можно поделиться с 15 другими людьми. [9] хотя самцы, как правило, спят одни, а самки, как правило, делят гнезда. [6]

Все мышиные лемуры очень активны в ночное время. [10] часто снуют, как мыши, и прыгают на высоту более 3 м (9,8 футов), [24] используя хвост как орган равновесия. [10] Передвигаясь среди конечных ветвей кустов и деревьев, они хватаются всеми четырьмя ногами и передвигаются четырьмя ногами. [10] [24] Находясь на земле, чтобы поймать насекомых или пересечь короткие открытые участки, мышиные лемуры прыгают, как лягушки. [10] Известно, что во время охоты серый мышиный лемур ловит беспозвоночных и мелких позвоночных быстрыми хватками. [19]
В засушливый сезон перед серым мышиным лемуром стоит задача эффективного использования редко распределенных кормовых ресурсов. Результаты недавнего исследования по этому вопросу показали, что серый мышиный лемур не передвигается случайно, а скорее использует пространственные сигналы для поиска пищевых ресурсов в отсутствие сенсорных сигналов, и что они, похоже, повторно используют общие, высокоэффективные маршруты для путешествий. расстояние. Считается, что вместо использования сети, основанной на маршрутах, серые мышиные лемуры обладают некоторым чувством мысленного представления своей пространственной среды, которое они используют для поиска и использования пищевых ресурсов. [30]
Поведение при поиске пищи часто бывает медленным, высота и направление постоянно меняются. Хищничество насекомых происходит преимущественно на земле. [7] Перед спуском ушные раковины попеременно движутся, помогая точно определить местонахождение добычи. Насекомые ловятся во время быстрого рывка по опавшей листве и переносятся ртом вверх, в относительную безопасность ветвей. [24] Исследования содержащихся в неволе серых лемуров-мышей показали, что зрение в основном используется для обнаружения добычи, хотя другие чувства, безусловно, играют роль в поиске пищи. [31]
Серый мышиный лемур всеяден. [9] Питаются преимущественно фруктами и беспозвоночными . [7] [24] Местное население, судя по всему, специализируется на местных фруктах. И в Маросалазе , и в Мандене жуки являются основной добычей насекомых, хотя мотыльков, богомолов , фульгоридных жуков также едят , сверчков, тараканов и пауков. Менее половины рациона составляют насекомые, немного большую часть составляют фрукты. [24] Этот лемур также потребляет цветы, десны и нектар деревьев молочай и терминалия , листья ( Uapaca sp.), экссудаты ( секреты личинок равнокрылых ) и мелких позвоночных , таких как древесные лягушки , гекконы и хамелеоны . [6] [7] [9] Его диета сезонно разнообразна и разнообразна по содержанию, что дает ему очень широкую кормовую нишу по сравнению с другими видами, такими как мышиный лемур мадам Берты. Следовательно, на него больше влияет наличие пищи, чем разделение ниш, в которых возникает симпатия. [32]
Покой
[ редактировать ]Как и все представители рода мышиных лемуров, серый мышиный лемур, как известно, впадает в кратковременное ежедневное состояние оцепенения, особенно во время прохладной и сухой в Южном полушарии . зимы [8] [14] [33] Эта редкая черта у приматов [34] в сочетании с легкостью наблюдения за этим видом в пределах его широкого географического распространения. [6] и его хорошее представительство в неволе, [35] делает его популярным объектом для исследований в качестве модельного организма .

Серый мышиный лемур уникален среди изученных на данный момент мышиных лемуров, поскольку это единственный вид, демонстрирующий длительное сезонное оцепенение, но такое поведение наблюдалось только в одном месте. [33] Модели активности могут заметно различаться между полами и группами населения. На лесной станции Ампихороа в национальном парке Анкарафансика самцы и самки проявляют ежедневное, а не сезонное оцепенение. [6] [14] В лесу Киринди оба пола ежедневно находятся в одинаковом оцепенении, однако в засушливый сезон (с апреля/мая по сентябрь/октябрь) самки становятся совершенно неактивными на несколько недель или до пяти месяцев, чтобы сохранить энергию и уменьшить хищничество. Однако самцы редко остаются бездействующими более нескольких дней и становятся чрезвычайно активными до того, как самки выходят из спячки, что позволяет им устанавливать иерархию и территории на сезон размножения . [6] [9] [14] Использование альтернативных стратегий энергосбережения в тех же условиях окружающей среды наблюдалось непосредственно в 2008 году, что стало первым физиологическим подтверждением на местах. [34] Эта картина сезонного и ежедневного оцепенения может быть связана с сезонностью региона. [24] поскольку Киринди - единственное место к западу от восточных горных тропических лесов, где в зимние месяцы по ночам наблюдаются очень низкие температуры. Вхождение в длительную оцепенение, иногда называемое спячкой, позволит снизить терморегуляторный стресс у самок. [33] тогда как самцы остаются более активными в подготовке к предстоящему брачному сезону. [6] Никакой разницы в смертности между находящимися в спячке самками и активными самцами не выявлено. [36]
Во время оцепенения скорость метаболизма серого мышиного лемура замедляется, и температура его тела падает до температуры окружающей среды, всего 7 ° C (45 ° F). В более прохладные месяцы с мая по август этот вид выбирает дупла деревьев ближе к уровню земли, где температура окружающей среды остается более стабильной. Это позволяет им дольше оставаться в спячке и сохранять метаболические ресурсы. [9] Одно исследование показало, что во время сезона размножения и самцы, и самки сокращали свои затраты энергии на 20%, когда гнездились парами, а максимальный энергетический выигрыш в 40% был достигнут, когда три мышиных лемура гнездились вместе. Даже в период отсутствия размножения максимальный прирост энергии наблюдался у двух и более животных, гнездящихся вместе, поскольку скорость метаболизма в состоянии покоя уже была снижена. [37]
Хотя серый мышиный лемур встречается как в первичных, так и во вторичных лиственных лесах, во вторичных лесах плотность их населения ниже. Это связано с тем, что различия в численности серых мышиных лемуров связаны с их способностью впадать в спячку во время засушливого сезона, особенно самок, которые, как правило, впадают в спячку дольше, чем самцы. В девственных лесах они могут ежедневно находиться в оцепенении или спячке, пока температура их тела остается ниже 28 ° C (82 ° F), но во вторичных лесах, где меньше крупных деревьев, температуры выше и подавляют способность серого мышиного лемура поддерживать вялость в течение длительного периода времени. Кроме того, серые мышиные лемуры во вторичных лесах, как правило, имеют меньшую массу тела, чем лемуры в девственных лесах, а также более низкую выживаемость. Вероятно, это связано с тем, что люди с меньшей массой тела с меньшей вероятностью впадают в оцепенение и, следовательно, тратят примерно на 40% больше энергии, чем те, кто сохраняет оцепенение. [38]
Эта необычная способность приматов проявлять спячку в дополнение к миниатюрным размерам привела исследователей к предположению, что предки лемуров и, возможно, предки приматов, возможно, имели некоторые общие черты с мышиными лемурами. Следовательно, серый мышиный лемур снова был использован в качестве модельного организма для изучения эволюции лемуров и приматов. [33] исследованиям , лемуры, как полагают, колонизировали Мадагаскар, сплавившись на остров около 60 миллионов лет назад Например, согласно молекулярно-филогенетическим . [39] [40] До того, как было обнаружено, что океанские течения были противоположны нынешним, что способствовало такому событию, [41] Считалось, что любому животному, не способному входить в состояние покоя, потребовалось бы слишком много времени, чтобы выжить в путешествии. Таким образом, считалось, что мышиные лемуры, такие как серый мышиный лемур, разделяли эту плезиоморфную (наследственную) черту с предковыми лемурами. [33]
Социальные системы
[ редактировать ]Серый мышиный лемур описывается как одинокий, но социальный человек . [24] Ночью добывают пищу в одиночку, но днем часто спят группами. [7] [9] Эта социальная модель варьируется в зависимости от пола, сезона и местоположения. Самки, как правило, делят гнезда с другими самками и их потомством, тогда как самцы, как правило, спят в одиночку или парами вне сезона размножения. [6] [7] [9] Группы самок, живущих в одном гнезде, могут быть относительно стабильными и состоять из двух-девяти особей, хотя самца можно встретить с группой самок вне сезона размножения. [10] В период размножения (с сентября по октябрь) самцы и самки могут спать в одной дупле дерева. [9] [10] В настоящее время могут быть распространены смешанные половые группы: одинокие самцы делят гнезда с тремя-семи самками, а одинокие самки делят гнезда с одним-тремя самцами. [7]
Исследования показали, что участки обитания серого мышиного лемура обычно невелики, возможно, менее 50 м (160 футов). [7] Самцы обычно путешествуют дальше ночью, и их ареалы в два раза больше, чем у самок. [6] [7] часто перекрываются друг с другом и всегда пересекаются по крайней мере с домашним ареалом одной самки. [9] [10] В период размножения ареал обитания самцов увеличивается в три раза. [6] [9]
Домашние ареалы самок перекрываются меньше, чем ареалы самцов, хотя локализованные концентрации, или «популяционные ядра», имеют тенденцию формироваться в некоторых областях, где соотношение полов в ядре ядра благоприятствует самкам и самцам в соотношении три или четыре к одному. [7] Генетические исследования показывают, что самки пространственно организуются в кластеры («популяционные ядра») родственных особей, в то время как самцы склонны эмигрировать из своей натальной группы . [6] Исследования показали, что самки этого вида могут поддерживать меньшие ареалы и более тесно общаться с другими самками, чем у некоторых других видов мышиных лемуров, из-за более оппортунистической пищевой ниши и, в случае популяций в Киринди, использования продолжительного сезонного оцепенения. . [42]
Коммуникация
[ редактировать ]
Вокализация и запах — основные способы общения внутри этого вида. Домашние территории отмечены запахами мочи и фекалий . [10] Вокализации сложные и очень высокие (от 10 до 36 кГц ), иногда выходящие за пределы человеческого слуха (от 0,02 до 20 кГц). [10] [43] К ним относятся призывы к поиску контакта, спариванию, дистанционному общению, тревоге и бедствию. [10]
Как и другие мышиные лемуры, серый мышиный лемур использует так называемый гармонический свист, который ниже по частоте и короче по продолжительности, чем его близкий родственник, коричневый мышиный лемур . Кроме того, было обнаружено, что типы вокализаций, издаваемых серым мышиным лемуром, могут зависеть от окружающей среды. В более открытых местах обитания сухих лесов, предпочитаемых серым мышиным лемуром, крики трелей более распространены и эффективны, поскольку они разносятся быстрее и с меньшей вероятностью маскируются ветром, в то время как щебетание чаще встречается у коричневого мышиного лемура, который предпочитает закрытые места обитания тропических лесов. [44]
Как и в случае с другими социальными млекопитающими, крики раскрывают пол и личность особи. Диалекты также были обнаружены между общинами. Трель самца, часть демонстрации спаривания самца, очень похожа на пение птиц с точки зрения упорядоченной последовательности слогов с широкополосной частотной модуляцией, высотой от 13 до 35 кГц и продолжительностью от 0,3 до 0,9 секунды, повторяющихся до 1,5 раз. в минуту. В каждой местности есть своя собственная тема трелей, отличная от таковой в соседних сообществах, и местные самцы издают индивидуально разные трели в рамках этой темы. Эти призывы не запрограммированы генетически . Во время игры молодые самцы рано пытаются издать трель, которая демонстрирует высокую степень изменчивости. Исследования показали, что самцы мышиных лемуров сознательно манипулируют диалектом, чтобы напоминать диалект своих соседей, когда их переводят из дома в новый район. Это может снизить агрессию и способствовать социальному принятию мужчин-эмигрантов, когда они переходят из своей родной группы по достижении зрелости. [43]
Поскольку мышиные лемуры относятся к загадочным видам и, следовательно, визуально неотличимы, их социальные крики помогают людям выбирать себе партнеров своего вида. Эта дифференцированная система передачи сигналов и распознавания способствовала сплочению видов за счет изоляции перед спариванием и помогла исследователям различать и идентифицировать виды. [45]
Разведение и размножение
[ редактировать ]Система спаривания описывается как многосамцовая и многоженская. Самцы устанавливают иерархию доминирования перед сезоном спаривания, однако некоторые исследования в дикой природе не выявили агрессии самцов или видимой конкуренции за восприимчивых самок. Самцы в неволе становятся очень агрессивными и формируют строгую иерархию доминирования. Эти самцы в неволе могут демонстрировать самые высокие в плазме уровни тестостерона , обнаруженные у млекопитающих, и даже запах доминирующего самца может снизить уровень тестостерона и сексуально подавлять подчиненного самца. [24] В период размножения семенники самцов значительно увеличиваются в размерах. [6] облегчение конкуренции сперматозоидов из-за женской распущенности . Исследования серого мышиного лемура показали, что оптимальный период оплодотворения, в течение которого самец с наибольшей вероятностью даст потомство, приходится на ранний период восприимчивости самки. [46] Лишь в брачный период смертность самцов превышает смертность самок. [36]
Хотя серый мышиный лемур демонстрирует модели спаривания, состоящие из нескольких самцов и нескольких самок, исследования показали, что самки действительно демонстрируют непрямой выбор партнера (форма избранной полиандрии). В ходе исследования самки спаривались с 1–7 самцами до 11 раз за одну ночь восприимчивости, но избегали или противодействовали самцам, которые пытались монополизировать спаривание. Доминирующие самцы, которые пытаются монополизировать, как правило, являются более крупными и тяжелыми особями. Однако было показано, что женский отбор помогает увеличить генетическое разнообразие потомства. [47]
Серый мышиный лемур считается в целом мономорфным по половому признаку, но сезонные колебания полового диморфизма наблюдаются по массе тела. Хотя масса тела обоих полов колеблется в течение года в зависимости от наличия пищи, будучи самой высокой в сезон дождей, различные поведенческие репертуары полов приводят к специфичным для пола закономерностям в этих колебаниях. Например, масса тела самцов увеличивается перед сезоном спаривания из-за значительного увеличения объема семенников, что, вероятно, повышает успех самцов в конкуренции спермы. [48] [49]
Самки восприимчивы в течение 45–55 дней с сентября по октябрь, а течка длится от 1 до 5 дней. [6] [9] [10] Самки сигнализируют о течке характерными высокочастотными криками и запаховой маркировкой. [9] Беременность длится от 54 до 68 дней, [10] в среднем 60 дней, в результате чего обычно рождается 2 или 3 потомка весом 5 г (0,18 унции) каждый. [6] [7] [9] [10] Младенцы рождаются в гнезде из листьев или дупла в ноябре, перед наступлением сезона дождей. [9] Отлучение происходит через 25 дней, [10] и детенышей либо оставляют в гнезде, либо носят во рту матери и кладут на ветку, пока она добывает пищу. [9] [24] Детеныши мышиных лемуров не цепляются за шерсть матери. Независимость достигается за 2 месяца, [7] [9] тогда как половая зрелость достигается в возрасте от 10 до 29 месяцев у женщин и от 7 до 19 месяцев у мужчин. [7] [10] Близкородственные самки остаются слабо связанными после созревания (женская филопатрия ), тогда как самцы расходятся из своей натальной области. В дикой природе репродуктивная продолжительность жизни серого мышиного лемура составляет не более 5 лет. [9] хотя, как сообщается, экземпляры в неволе жили до 15 лет и 5 месяцев, [10] или даже до 18,2 лет. [50]
Серый мышиный лемур использует совместное разведение как форму семейной страховки. Самка регулярно переносит свое потомство в гнезда других самок, а также ухаживает за потомством, кроме своего собственного. Хотя это может иметь высокие физиологические издержки для кормящей самки, которая и так тратит много энергии, в целом это может быть полезно для обеспечения выживания среди близкородственных групп с высоким риском смертности. Исследование, проводившееся в течение трех сезонов размножения, показало, что близкородственные самки образуют гнездовые группы главным образом при нехватке подходящих насестов; когда есть преимущество общего гнезда для защиты или когда есть преимущества терморегуляции. В случае усыновления потомства, когда родитель умирает и заботу о нем берет на себя близкородственная самка, считается, что это выгодно группам с высоким риском смертности. [51]
Статус сохранения
[ редактировать ]Серый мышиный лемур был внесен в Приложение 1 Конвенции о международной торговле видами, находящимися под угрозой исчезновения (СИТЕС) в 1975 году. [52] объявив его находящимся под угрозой исчезновения и запретив международную торговлю образцами, за исключением некоммерческого использования, например, для научных исследований. [2] (МСОП) 2008 года В оценке Красного списка Международного союза охраны природы он внесен в список видов, вызывающих наименьшее беспокойство (LC) с тенденцией к сокращению численности популяции. [1]
Наибольшей угрозой для него являются утрата среды обитания в результате подсечно-огневого земледелия и выпаса скота , а также отлов живых животных для местной торговли домашними животными в северной и южной частях его ареала. Хотя этот вид населяет вторичные леса, исследования показали, что снижение качества среды обитания отрицательно влияет на его популяцию, поскольку меньшее количество дупл в деревьях дает меньше возможностей для сохранения энергии, увеличивая стресс и смертность. [1] [6] Одно исследование обнаружило девять видов паразитов в фекалиях серого мышиного лемура, живущего в лесах, подвергшихся деградации и фрагментации. На небольших фрагментах леса хорошего качества, а также в лесах с высокой степенью деградации у серого мышиного лемура наблюдалась более высокая распространенность паразитических нематод и простейших, чем на более крупных фрагментах леса хорошего качества. [53] Исследования конца 1960-х и 1970-х годов показали, что интенсивные лесозаготовки в период с 1968 по 1970 годы, по-видимому, привели к снижению массы тела, использованию меньших деревьев в качестве мест гнездования и меньшему максимальному размеру гнездовой группы самок (до 7 из 15). [7] Существует также обеспокоенность тем, что, хотя ежедневное оцепенение может помочь сохранить энергию и ресурсы во время умеренной нехватки продовольствия, продолжительная нехватка продовольствия, вызванная изменением климата, может создать слишком сильный стресс и серьезно повлиять на выживаемость вида. [54] [55]
Серый мышиный лемур считается одним из самых распространенных мелких местных млекопитающих Мадагаскара. Он обитает в семи национальных парках, пяти специальных заповедниках, частном заповеднике Беренти и других частных охраняемых лесах в пределах заповедной зоны Мандена. [6]
Этот вид мышиных лемуров очень хорошо размножается в неволе, хотя в зоопарках его обычно не показывают, как некоторых более крупных дневных лемуров. В 1989 году более 370 особей были размещены в 14 учреждениях Международной системы информации о видах (ISIS) и учреждениях, не входящих в ИГИЛ, в Соединенных Штатах и Европе, 97% из которых родились в неволе. [7] В марте 2009 года 167 человек были зарегистрированы в 29 учреждениях ИГИЛ, включая Центр Дюка Лемура . [1] [35]
Примечания
[ редактировать ]- ^ Перейти обратно: а б с д и ж Рейтер, Кентукки; Бланко, М.; Ганцхорн, Дж.; Швитцер, К. (2020). « Микроцебус муринус » . Красный список видов, находящихся под угрозой исчезновения МСОП . 2020 : e.T163314248A182239898. doi : 10.2305/IUCN.UK.2020-3.RLTS.T163314248A182239898.en . Проверено 19 ноября 2021 г.
- ^ Перейти обратно: а б «Контрольный список видов СИТЕС» . СИТЕС . ЮНЕП-ВЦМК . Проверено 18 марта 2015 г.
- ^ Перейти обратно: а б с д Гроувс, CP (2005). « Микроцебус муринус » . В Уилсоне, Делавэр ; Ридер, Д.М. (ред.). Виды млекопитающих мира: таксономический и географический справочник (3-е изд.). Балтимор: Издательство Университета Джонса Хопкинса. п. 113. ИСБН 0-801-88221-4 . OCLC 62265494 .
- ^ Аллен, генеральный директор (1939). «Контрольный список африканских млекопитающих». Вестник Музея сравнительной зоологии . 83 : 1–763.
- ^ Палмер, Т. (1904). Указатель рода млекопитающих: список родов и семейств млекопитающих . Правительственная печать США. стр. 421 , 435.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа Миттермайер, РА; и др. (2006). « М. мышиная », с. 104–107
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и Харкорт, К.; Торнбэк, Дж. (1990). Лемуры Мадагаскара и Коморских островов. Красная книга МСОП . МСОП. стр. 32–38 . ISBN 978-2-88032-957-0 .
- ^ Перейти обратно: а б Миттермайер, РА; и др. (2006). «Мышиные лемуры», стр. 91–128.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и Гарбутт, Н. (2007). Млекопитающие Мадагаскара, Полное руководство . А&С Черный . стр. 86–88. ISBN 978-0-300-12550-4 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с Новак, Р.М. (1999). Приматы мира Уокера . Издательство Университета Джонса Хопкинса. стр. 66–67, 126 . ISBN 978-0-8018-6251-9 .
- ^ Луи-младший, Э.; Энгберг, С.; Макгуайр, С.; Маккормик, М.; Рандриамампионона, Р.; Ранайвоарисоа, Дж.; Бейли, К.; Миттермайер, Р.; Лей, Р. (2008). «Пересмотр мышиных лемуров, Microcebus (приматов, Lemuriformes) Северного и Северо-Западного Мадагаскара с описаниями двух новых видов в национальном парке Монтань д'Амбр и классифицированном лесу Антафондро» (PDF) . Сохранение приматов . 23 :19–38. дои : 10.1896/052.023.0103 . S2CID 84533039 . Архивировано из оригинала (PDF) 5 февраля 2009 г.
- ^ Миллер, Джон Фредерик (1777). Icones Animalium et Plantarum. Различные предметы естествознания, в которых изображены птицы, животные и множество любопытных растений и т. д. (на латыни). Лондон. Часть 3, Табличка 13. Издано в 10 частях по 6 пластин в каждой. Видеть: Шерборн, CD ; Иредейл, Т. (1921). Дж. Ф. Миллера « Иконы » . Ибис . 11-я серия. 3 : 302–309. дои : 10.1111/j.1474-919X.1921.tb00801.x .
- ^ Маккенна, MC; Белл, СК (1997). Классификация млекопитающих: выше уровня вида . Издательство Колумбийского университета. п. 335. ИСБН 978-0-231-11013-6 .
- ^ Перейти обратно: а б с д Каппелер, премьер-министр; Расолоарисон, РМ (2003). « Микроцебусы , Мышиные Лемуры, Циды ». В Гудмане, С.М.; Бенстед, JP (ред.). Естественная история Мадагаскара . Издательство Чикагского университета. стр. 1310–1315. ISBN 978-0-226-30306-2 .
- ^ Миттермайер, Р. ; Ганцхорн, Дж.; Констант, В.; Гландер, К.; Таттерсолл, И .; Гровс, К. ; Райландс, А.; Хапке, А.; и др. (2008). «Разнообразие лемуров на Мадагаскаре» . Международный журнал приматологии . 29 (6): 1607–1656. дои : 10.1007/s10764-008-9317-y . hdl : 10161/6237 . S2CID 17614597 . Архивировано из оригинала (PDF) 15 февраля 2021 г. Проверено 20 сентября 2019 г.
- ^ Брауне, П.; Шмидт, С.; Циммерманн, Э. (2008). «Акустическая дивергенция в общении загадочных видов ночных приматов ( Microcebus ssp. )» . БМК Биология . 6 : 1–10. дои : 10.1186/1741-7007-6-19 . ПМК 2390514 . ПМИД 18462484 .
- «Все началось со скрипа: серенада лунного света помогает лемурам выбирать себе пару подходящего вида» . ScienceDaily (пресс-релиз). 14 мая 2008 г.
- ^ Оливьери, Г.; Циммерманн, Э.; Рандрианамбинина, Б.; Рассолохарияона, С.; Ракотондравони, Д.; Гущанский, К.; Радешпиль, У. (2006). «Постоянно растущее разнообразие мышиных лемуров: три новых вида на севере и северо-западе Мадагаскара». Молекулярная филогенетика и эволюция . 43 (1): 309–327. дои : 10.1016/j.ympev.2006.10.026 . ПМИД 17197200 .
- ^ Луис-младший, Э.Э.; Энгберг, SE; МакГуайр, С.М.; Маккормик, MJ; Рандриамампионона, Р.; Ранайвоарисоа, Дж. Ф.; Бейли, Калифорния; Миттермайер, РА; Лей, Р. (2008). «Пересмотр мышиных лемуров, Microcebus (приматов, Lemuriformes) Северного и Северо-Западного Мадагаскара с описаниями двух новых видов в национальном парке Монтань д'Амбр и классифицированном лесу Антафондро» (PDF) . Сохранение приматов . 23 :19–38. дои : 10.1896/052.023.0103 . S2CID 84533039 . Архивировано из оригинала (PDF) 5 февраля 2009 г.
- ^ Перейти обратно: а б Флигл, Дж. (1999). Адаптация и эволюция приматов (2-е изд.). Академическая пресса. п. 86 . ISBN 978-0-12-260341-9 .
- ^ Малдун, КМ; де Блье, Д.Д.; Саймонс, Эл.; Чатрат, PS (2009). «Распространение субископаемых и палеоэкологическое значение мелких млекопитающих в пещере Анкилитело, юго-запад Мадагаскара» . Журнал маммологии . 90 (5): 1111–1131. doi : 10.1644/08-МАММ-А-242.1 .
- ^ Миттермайер, РА ; Таттерсолл, И .; Констант, WR; Мейерс, DM; Маст, РБ (1994). Лемуры Мадагаскара . Иллюстрировано С.Д. Нэшем (1-е изд.). Интернационал Консервации . п. 34. ISBN 1-881173-08-9 . OCLC 32480729 .
- ^ Перейти обратно: а б Анкель-Саймонс, Ф. (2007). Анатомия приматов (3-е изд.). Академическая пресса. п. 591. ИСБН 978-0-12-372576-9 .
- ^ Дютрилья, Б.; Румплер, Ю. (2005). «Аналогия хромосомных полос между полуобезьянами ( Microcebus murinus ), платирринами ( Cebus capucinus ) и человеком». Американский журнал физической антропологии . 52 (1): 133–137. дои : 10.1002/ajpa.1330520116 . ПМИД 6768303 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т Сассман, Р. (1999). Экология приматов и социальная структура. Том 1: Лори, лемуры и долгопяты . Pearson Custom Publishing. стр. 107–148 . ISBN 978-0-536-02256-1 .
- ^ Ракотондравони, Р.; Радешпиль, У. (2009). «Различные модели сосуществования двух видов мышиных лемуров ( Microcebus ravelobensis и M. murinus ) в гетерогенном ландшафте». Американский журнал приматологии . 71 (11): 928–938. дои : 10.1002/ajp.20732 . ПМИД 19676112 . S2CID 38075778 .
- ^ Даммхан, М.; Каппелер, премьер-министр (2008). «Маломасштабное сосуществование двух видов мышиных лемуров ( Microcebus berthae и M. murinus ) в однородной конкурентной среде» . Экология . 157 (3): 473–483. Бибкод : 2008Oecol.157..473D . дои : 10.1007/s00442-008-1079-x . ПМК 2515545 . ПМИД 18574599 .
- ^ Хямяляйнен, А. (2012). «Случай каннибализма у взрослой особи серого мышиного лемура ( Microcebus murinus )». Американский журнал приматологии . 74 (9): 783–787. дои : 10.1002/ajp.22034 . ПМИД 22623272 . S2CID 13022858 .
- Чарльз Чой (8 июня 2012 г.). «ScienceShot: каннибализм замечен у серого мышиного лемура» . Наука .
- ^ Эберле, М.; Каппеле, премьер-министр (2008). «Мутуализм, взаимность или родственный отбор? Совместное спасение особи вида от удава у серого мышиного лемура, ведущего ночной одиночный образ жизни» (PDF) . Американский журнал приматологии . 70 (4): 410–414. дои : 10.1002/ajp.20496 . ПМИД 17972271 . S2CID 464977 . Архивировано из оригинала (PDF) 19 июля 2011 г.
- ^ Зюндерманн, Д.; Шойман, М.; Циммерманн, З. (2008). «Распознавание обонятельных хищников у серых мышиных лемуров, не привыкших к хищникам ( Microcebus murinus )». Журнал сравнительной психологии . 122 (2): 146–155. дои : 10.1037/0735-7036.122.2.146 . ПМИД 18489230 .
- ^ Лурс, МЛ; Даммхан, М.; Каппелер, премьер-министр; Фихтель, К. (2009). «Пространственная память у серого мышиного лемура ( Microcebus murinus )» . Познание животных . 12 (4): 599–609. дои : 10.1007/s10071-009-0219-y . ПМЦ 2698973 . ПМИД 19263100 .
- ^ Пип, М.; Радешпиль, У.; Циммерманн, Э.; Шмидт, С.; Симерс, Б.М. (2008). «Сенсорные основы обнаружения добычи у рожденных в неволе серых мышиных лемуров Microcebus murinus ». Поведение животных . 73 (3): 871–878. дои : 10.1016/j.anbehav.2007.07.008 . S2CID 53164748 .
- ^ Даммхан, М.; Каппелер, премьер-министр (2008). «Сравнительная экология питания симпатрических Microcebus berthae и M. murinus ». Международный журнал приматологии . 29 (6): 1567–1589. дои : 10.1007/s10764-008-9312-3 . S2CID 37601666 .
- ^ Перейти обратно: а б с д и Радешпиль, У. (2006). «Глава 10: Экологическое разнообразие и сезонная адаптация мышиных лемуров ( Microcebus spp.)». Ин Гулд, Л.; Заутер, М.Л. (ред.). Лемуры: экология и адаптация . Спрингер. стр. 211–234 . ISBN 978-0-387-34585-7 .
- ^ Перейти обратно: а б Шмид, Дж.; Ганцхорн, Ю. (2009). «Дополнительные стратегии снижения метаболизма у серых мышиных лемуров». Naturwissenschaften . 96 (6): 737–741. Бибкод : 2009NW.....96..737S . дои : 10.1007/s00114-009-0523-z . ПМИД 19277596 . S2CID 23107653 .
- ^ Перейти обратно: а б «Холдинг видов ИГИЛ, Microcebus murinus » . Международная информационная система видов (ISIS). 2009. Архивировано из оригинала 15 мая 2006 года . Проверено 25 марта 2009 г.
- ^ Перейти обратно: а б Краус, К.; Эберле, Э.; Каппелер, премьер-министр (2008). «Цена рискованного поведения самцов: половые различия в сезонном выживании мелких мономорфных по половому признаку приматов» . Учеб. Р. Сок. Б. 275 (1643): 1635–1644. дои : 10.1098/rspb.2008.0200 . ПМК 2602817 . ПМИД 18426751 .
- ^ Перре, М. (1998). «Энергетическое преимущество совместного гнездования у одиночного примата, малого мышиного лемура (Microcebus murinus)» . Журнал маммологии . 49 (4): 1093–1102. дои : 10.2307/1383001 . JSTOR 1383001 .
- ^ Ганцхорн, Дж.; Шмид, Дж. (1998). «Различная динамика популяций Microcebus murinus в первичных и вторичных лиственных сухих лесах Мадагаскара». Международный журнал приматологии . 19 (5): 785–796. дои : 10.1023/A:1020337211827 . S2CID 19896069 .
- ^ Миттермайер, РА; и др. (2006) «Глава 1: Происхождение лемуров», стр. 23–26.
- ^ Йодер, AD; Ян, З. (2004). «Даты дивергенции малагасийских лемуров оценены на основе нескольких генных локусов: геологический и эволюционный контекст» (PDF) . Молекулярная экология . 13 (4): 757–773. Бибкод : 2004MolEc..13..757Y . дои : 10.1046/j.1365-294X.2004.02106.x . ПМИД 15012754 . S2CID 14963272 .
- ^ Али, младший; Хубер, М. (2010). «Биоразнообразие млекопитающих на Мадагаскаре, контролируемое океанскими течениями». Природа . 463 (7281): 653–6. Бибкод : 2010Natur.463..653A . дои : 10.1038/nature08706 . ПМИД 20090678 . S2CID 4333977 .
- «Животные заселили Мадагаскар, сплавляясь по нему» . ScienceDaily (пресс-релиз). 21 января 2010 г.
- ^ Даммхан, М.; Каппелер, премьер-министр (2009). «Схватка или соревнование за еду у одиночных мышей-лемуров ( Microcebus spp.): Новые идеи на основе стабильных изотопов». Американский журнал физической антропологии . 141 (2): 181–189. дои : 10.1002/ajpa.21129 . ПМИД 19591211 .
- ^ Перейти обратно: а б Макдональд, Д. (2006). «Приматы». Энциклопедия млекопитающих . Компания Brown Reference Group plc. п. 319. ИСБН 978-0-681-45659-4 .
- ^ Циммерманн, Э.; Воробьева Е.; Рогеманн, Д.; Хафен, Т. (2000). «Использование голосовых отпечатков пальцев для специфической дискриминации серых ( Microcebus murinus ) и рыжих мышиных лемуров ( Microcebus rufus )». Международный журнал приматологии . 21 (9): 837–852. дои : 10.1023/А:1005594625841 . S2CID 43736948 .
- ^ Брауне, П.; Шмидт, С.; Циммерманн, Э. (2008). «Акустическая дивергенция в общении загадочных видов ночных приматов ( Microcebus ssp. )» . БМК Биология . 6:19 . дои : 10.1186/1741-7007-6-19 . ПМК 2390514 . ПМИД 18462484 .
- ^ Эберле, М.; Перре, М.; Каппелер, премьер-министр (2007). «Конкуренция сперматозоидов и оптимальное время спаривания у Microcebus murinus ». Международный журнал приматологии . 28 (6): 1267–1278. дои : 10.1007/s10764-007-9220-y . S2CID 25738388 .
- ^ Эберле, М.; Каппелер, П. (2004). «Избранная полиандрия: женский выбор и межполовой конфликт у маленького ночного одиночного примата ( Microcebus murinus )». Поведенческая экология и социобиология . 57 (1): 91–100. дои : 10.1007/s00265-004-0823-4 . S2CID 43994124 .
- ^ Хямяляйнен, А.; Даммхан, М.; Ожар, Ф.; Эберле, М.; Харди, И.; Каппелер, премьер-министр; Перре, М.; Шлихе-Дикс, С.; Краус, К. (2014). «Старение или избирательное исчезновение? Возрастные траектории массы тела в диких и содержащихся в неволе популяциях приматов с небольшим телом» . Труды Королевского общества B: Биологические науки . 281 (1791): 20140830. doi : 10.1098/rspb.2014.0830 . ПМЦ 4132673 . ПМИД 25100693 .
- ^ Шмид, Дж.; Каппелер, премьер-министр (1998). «Колебательный половой диморфизм и дифференциальная спячка в зависимости от пола у примата, серого мышиного лемура ( Microcebus murinus )». Поведенческая экология и социобиология . 43 (2): 125–132. дои : 10.1007/s002650050474 . S2CID 36249151 .
- ^ Леман, Шон М.; Радешпиль, Юте; Циммерманн, Эльке (7 апреля 2016 г.). Карликовые и мышиные лемуры Мадагаскара: биология, поведение и биогеография сохранения Cheirogaleidae . Издательство Кембриджского университета. ISBN 978-1-316-55278-0 .
- ^ Эберле, М.; Каппелер, П. (2006). «Семейное страхование: родственный отбор и совместное разведение одиночного примата ( Microcebus murinus )». Поведенческая экология и социобиология . 60 (4): 582–588. дои : 10.1007/s00265-006-0203-3 . S2CID 22186719 .
- ^ «База данных ЮНЕП-WCMC по видам: виды, внесенные в список СИТЕС, Microcebus murinus » . Программа ООН по окружающей среде – Всемирный центр мониторинга охраны природы. 2009 . Проверено 29 марта 2009 г. [ постоянная мертвая ссылка ]
- ^ Рахаривололона, М.; Ганцхорн, Дж. (2009). «Желудочно-кишечная паразитарная инфекция серого мышиного лемура ( Microcebus murinus ) в прибрежном лесу Мандена, Мадагаскар: последствия фрагментации и деградации леса» . Сохранение и развитие Мадагаскара . 4 (2): 103–112. дои : 10.4314/mcd.v4i2.48650 .
- ^ Жиру, С.; Перре, М.; Штейн, П.; Гудэбл, Дж.; Ожар, Ф.; Гилберт, К.; Робин, JP; Махо, ЮЛ; и др. (2010). Бартелл, Пол А. (ред.). «Серый мышиный лемур использует сезонно-зависимые стратегии экономии жира или белка, чтобы справиться с хроническим ограничением в еде» . ПЛОС ОДИН . 5 (1): е8823. Бибкод : 2010PLoSO...5.8823G . дои : 10.1371/journal.pone.0008823 . ПМК 2809095 . ПМИД 20098678 .
- ^ Жиру, С.; Блан, С.; Ожар, Ф.; Бертран, Ф.; Гилберт, К.; Перре, М. (2008). «Хроническая нехватка пищи и сезонные модуляции суточной оцепенения и двигательной активности у серого мышиного лемура ( Microcebus murinus )». Американский журнал физиологии. Регуляторная, интегративная и сравнительная физиология . 294 (6): R1958–R1967. дои : 10.1152/ajpregu.00794.2007 . ПМИД 18434438 .
Ссылки
[ редактировать ]- Миттермайер, РА ; Констант, WR; Хокинс, Ф.; Луи, EE ; и др. (2006). Лемуры Мадагаскара . Иллюстрировано С.Д. Нэшем (2-е изд.). Интернационал Консервации . ISBN 1-881173-88-7 . OCLC 883321520 .
Внешние ссылки
[ редактировать ] СМИ, связанные с Microcebus murinus, на Викискладе?
СМИ, связанные с Microcebus murinus, на Викискладе? - Просмотрите геном мышиного лемура в Ensembl .
- Просмотрите сборку генома micMur2 в браузере генома UCSC .
| Базы данных органов управления : Национальные |
|---|