Хитон
| Хитон Временной диапазон: | |
|---|---|
 | |
| Хитон с живой подкладкой Tonicella lineata , сфотографированный на месте : передний конец животного направлен вправо. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Моллюска |
| Сорт: | Полиплакофора Бленвиль , 1816 г. |
| Подгруппы | |
Хитоны ( / ˈ k aɪ t ɒ n z , ˈ k aɪ t ən z / ) — морские моллюски размера в классе Polyplacophora ( / ˌ p ɒ l i p lə k ˈ ɒ f ərə pl- разного / POL -ee- КОФ -эр-а ), [3] ранее известный как Амфиневра . [4] Около 940 [5] [6] сохранившиеся и 430 [7] ископаемые виды признаны.
Их также иногда называют морскими колыбельками , кольчужными раковинами или сосущими камнями , или, более формально, как лорикатами , полиплакофорами , а иногда и полиплакофорами .
Хитоны имеют раковину, состоящую из восьми отдельных раковинных пластинок или створок . [3] Эти пластины слегка перекрываются по переднему и заднему краям, но при этом хорошо сочленяются друг с другом. Из-за этого панцирь обеспечивает защиту и в то же время позволяет хитону изгибаться вверх, когда это необходимо для передвижения по неровным поверхностям, и даже позволяет животному сворачиваться в клубок, когда его сбивают с камней. [8] Пластины панциря окружены юбкой, известной как пояс .
среда обитания
[ редактировать ]
Хитоны живут по всему миру, от холодных вод до тропиков. Они живут на твердых поверхностях, например, на камнях или под ними, или в расщелинах скал.
Некоторые виды обитают довольно высоко в приливной зоне и подвергаются воздействию воздуха и света в течение длительного времени. Большинство видов обитают в приливных или сублиторальных зонах и не выходят за пределы фотической зоны , но некоторые виды живут в глубокой воде, на глубине до 6000 м (20 000 футов). [9]
Хитоны исключительно и полностью морские существа, в отличие от двустворчатых моллюсков , способных адаптироваться к солоноватой и пресной воде, и брюхоногих моллюсков , способных успешно переходить в пресноводную и наземную среду.
Морфология
[ редактировать ]Оболочка
[ редактировать ]Все хитоны имеют защитную спинную оболочку, разделенную на восемь сочленяющихся арагонитовых клапанов, встроенных в прочный мышечный пояс, окружающий тело хитона. По сравнению с одиночными или состоящими из двух частей раковинами других моллюсков такое расположение позволяет хитонам при смещении сворачиваться в защитный шар и плотно прилегать к неровным поверхностям. У некоторых видов створки редуцированы или покрыты опоясывающей тканью. [10] [11] Створки бывают разного цвета, с рисунком, гладкие или рельефные.
| Часть серии о |
| Ракушки |
|---|
 |
| Раковины моллюсков |
| О раковинах моллюсков |
| Другие ракушки |


Самая передняя пластинка имеет серповидную форму и известна как головная пластинка (иногда называемая головной пластинкой, несмотря на отсутствие полной головы). Самая задняя пластинка известна как анальная пластинка (иногда называемая хвостовой пластинкой, хотя у хитонов хвостов нет).
Внутренний слой каждой из шести промежуточных пластинок образуется спереди в виде сочленяющегося фланца, называемого суставом. Этот внутренний слой также может быть изготовлен сбоку в виде вставных пластин с надрезами. Они функционируют как крепление пластин клапанов к мягкому телу. Аналогичный ряд вставочных пластинок может быть прикреплен к выпуклому переднему краю головной пластинки или выпуклому заднему краю анальной пластинки. [12]
Скульптура створок является одним из таксономических признаков наряду с грануляцией или шиповчатостью пояска. [12]
После отмирания хитона отдельные створки, составляющие панцирь, состоящий из восьми частей, распадаются, потому что пояс больше не удерживает их вместе, и тогда пластины иногда смываются пляжным наносом. Отдельные пластинки панциря хитона иногда называют раковинами-бабочками из-за их формы.
Орнамент пояса
[ редактировать ]Пояс может быть украшен чешуйками или спикулами, которые, как и пластинки раковины, минерализованы арагонитом, хотя в спикулах протекает другой процесс минерализации, чем в зубах или раковинах (что подразумевает независимое эволюционное нововведение). [11] Этот процесс кажется довольно простым по сравнению с другими тканями оболочки; у некоторых таксонов кристаллическая структура отложенных минералов очень напоминает неупорядоченную природу кристаллов, образующихся неорганическим путем, хотя в других таксонах наблюдается больший порядок. [11]
Белковый компонент чешуи и склеритов мизерен по сравнению с другими биоминерализованными структурами, тогда как общая доля матрикса «выше», чем в раковинах моллюсков. Это означает, что полисахариды составляют основную часть матрицы. [11] Поясные шипы часто несут продольно-параллельные исчерченности. [11]
Широкая форма орнамента пояса предполагает, что он выполняет второстепенную роль; хитоны прекрасно могут выжить и без них. Камуфляж или защита — две вероятные функции. [11] Конечно, такие виды, как некоторые представители рода Acanthochitona, имеют на поясе заметные парные пучки спикул. Спикулы острые и при неосторожном обращении легко проникают в кожу человека, где отслаиваются и остаются болезненным раздражителем. [13]
Спикулы секретируются клетками, которые не экспрессируют engrailed , но эти клетки окружены клетками, экспрессирующими engrailed. [14] Эти соседние клетки выделяют органическую пелликулу на внешней стороне развивающейся спикулы, арагонит которой откладывается центральной клеткой; последующее деление этой центральной клетки позволяет у некоторых таксонов выделять более крупные шипы. [15] Органическая пелликула обнаружена у большинства полиплакофор (но не у базальных хитонов, таких как Hanleya ). [15] но необычен для аплакофоры. [16] В процессе развития клетки, секретирующие склериты, возникают из претрохальных и построхальных клеток: клеток 1a, 1d, 2a, 2c, 3c и 3d. [16] Пластинки раковины возникают преимущественно из 2d-микромера, хотя в его секреции участвуют также клетки 2a, 2b, 2c, а иногда и 3c. [16]
Внутренняя анатомия
[ редактировать ]Пояс часто украшен спикулами, щетинками, волосистыми пучками, шипами или змеевидными чешуйками. Большая часть тела представляет собой стопу, напоминающую улитку , но с дорсальной стороны не видно ни головы, ни других мягких частей за поясом.Мантийная полость состоит из узких каналов с каждой стороны, лежащих между телом и поясом. Вода поступает в полость через отверстия по обе стороны рта, затем течет по каналу ко второму, выдыхающему отверстию, расположенному вблизи ануса . [17] Множественные жабры свисают в мантийную полость вдоль части или всей латеральной мантийной борозды, каждая из которых состоит из центральной оси с множеством уплощенных нитей, через которые может поглощаться кислород. [18]
Трехкамерное сердце расположено ближе к заднему концу животного. Каждое из двух предсердий собирает кровь из жабр с одной стороны, в то время как мышечный желудочек перекачивает кровь через аорту и вокруг тела.
Выделительная система состоит из двух нефридиев , которые соединяются с полостью перикарда вокруг сердца и выводят экскременты через пору, открывающуюся вблизи задней части мантийной полости. Единственная гонада расположена перед сердцем и выпускает гаметы через пару пор непосредственно перед теми, которые используются для выделения. [18]

Рот радулой расположен на нижней стороне животного и содержит языкообразную структуру, называемую , которая имеет многочисленные ряды по 17 зубов в каждом. Зубы покрыты магнетитом , твёрдым минералом из оксида железа/железа. Радула используется для соскребания микроскопических водорослей с субстрата. Сама ротовая полость выстлана хитином и связана с парой слюнных желез . В задней части рта открываются два мешочка, один из которых содержит радулу, а другой — выступающий сенсорный субрадулярный орган , который прижимается к субстрату для определения вкуса пищи. [18]
Реснички вытягивают пищу через рот в виде струи слизи и через пищевод , где она частично переваривается ферментами пары крупных глоточных желез. Пищевод, в свою очередь, открывается в желудок , где ферменты пищеварительной железы завершают расщепление пищи. Питательные вещества всасываются через слизистую оболочку желудка и первый отдел кишечника . Кишечник разделен на две части сфинктером , причем последняя часть сильно извита и функционирует для уплотнения отходов в фекальные шарики . Анус открывается сразу за стопой. [18]
У хитонов отсутствует четко очерченная голова; их нервная система напоминает рассыпанную лестницу. [19] , как у других моллюсков, нет Настоящих ганглиев , хотя вокруг пищевода имеется кольцо плотной нервной ткани. От этого кольца нервы отходят вперед, иннервируя полость рта и субрадулу, а две пары основных нервных тяжей проходят назад через тело. Одна пара, педальные связки, иннервируют стопу, а паллиовисцеральные связки иннервируют мантию и остальные внутренние органы. [18]
Некоторые виды несут перед головой множество щупалец. [20]
Чувства
[ редактировать ]Основными органами чувств хитонов являются субрадулярный орган и большое количество уникальных органов, называемых эстетами . Эстеты состоят из светочувствительных клеток чуть ниже поверхности панциря, хотя они не способны к истинному зрению. Однако в некоторых случаях они модифицируются, образуя глазки , при этом скопление отдельных фоторецепторных клеток лежит под небольшой арагонита на основе линзой . [21] Каждая линза может формировать четкое изображение и состоит из относительно крупных зерен с высокой кристаллографической ориентацией, что позволяет минимизировать рассеяние света. [22] Отдельный хитон может иметь тысячи таких глазков. [18] Эти арагонита глаза на основе [23] сделать их способными к истинному видению; [24] хотя исследования продолжаются в отношении степени их остроты зрения. Известно, что они могут различать тень хищника и изменения света, вызванные облаками. Эволюционный компромисс привел к компромиссу между глазами и панцирем; по мере увеличения размера и сложности глаз механические характеристики их оболочек уменьшаются, и наоборот. [25]
Существует относительно хорошая летопись окаменелостей хитонных панцирей, но глазки присутствуют только в тех, которые датируются 10 миллионами лет назад или моложе; это сделало бы глазки, точная функция которых неясна, вероятно, самыми последними эволюционировавшими глазами. [19]
Хотя у хитонов отсутствуют осфрадии , статоцисты и другие органы чувств, общие для других моллюсков, у них есть многочисленные осязательные нервные окончания, особенно на поясе и внутри мантийной полости.
Отряд Lepidopleurida также имеет пигментированный орган чувств, называемый органом Швабе. [26] Его функция остается в значительной степени неизвестной, и предполагается, что она связана с функцией личиночного глаза. [27]
Однако у хитонов отсутствует мозговой ганглий. [28]
Способность самонаведения
[ редактировать ]Подобно многим видам морских блюдец , некоторые виды хитонов, как известно, демонстрируют поведение самонаведения , путешествуя в поисках корма, а затем возвращаясь в то место, где они ранее обитали. [29] Метод, который они используют для такого поведения, в некоторой степени исследован, но остается неизвестным. По одной из теорий, хитоны запоминают топографический профиль региона и, таким образом, могут вернуться к своему домашнему шраму с помощью физического знания горных пород и визуальной информации от своих многочисленных примитивных глазных пятен. [30] Морская улитка Nerita textilis (как и все брюхоногие моллюски ) при движении оставляет слизистый след, который хеморецептивный орган способен обнаружить и направить улитку обратно на родное место. [31] Неясно, действует ли хитоновое самонаведение таким же образом, но они могут оставлять химические сигналы вдоль поверхности камня и в шраме дома, которые их обонятельные чувства могут обнаружить и нацелиться на них. Кроме того, могут быть обнаружены и более старые следы, что дает хитону дополнительный стимул найти свой дом. [30]
Радулярные зубцы хитонов состоят из магнетита , и находящиеся в них кристаллы железа могут участвовать в магниторецепции . [32] Земли способность чувствовать полярность и наклон магнитного поля . Экспериментальные работы показали, что хитоны могут обнаруживать магнетизм и реагировать на него. [33]
Кулинарное использование
[ редактировать ]Хитоны едят в нескольких частях света. Сюда входят острова в Карибском бассейне, такие как Тринидад , Тобаго , Багамские острова , Синт-Мартен, Аруба, Бонайре, Ангилья и Барбадос , а также на Бермудских островах. Их также традиционно едят в некоторых частях Филиппин , где их называют кибет, если они сырые, и хитон, если они жареные. Коренные жители тихоокеанского побережья Северной Америки питаются хитонами. Они являются обычным продуктом питания на тихоокеанском побережье Южной Америки и на Галапагосских островах . Ножку хитона готовят так же, как и морское ушко . Некоторые островитяне, живущие в Южной Корее, также едят хитон, слегка отваренный и смешанный с овощами и острым соусом. Аборигены Австралии также едят хитон; например, они записаны в Соглашении о традиционном рыболовстве народа Нарунгга .
Жизненные привычки
[ редактировать ]
Хитон медленно ползет по мускулистой стопе. Он обладает значительной силой сцепления и может очень сильно цепляться за камни, как блюдечко .
Хитоны, как правило, травоядные травоядные животные, хотя некоторые из них всеядны, а некоторые плотоядны. [34] [35] Они поедают водоросли , мшанки , диатомовые водоросли , ракушки , а иногда и бактерии , соскребая каменистый субстрат своими хорошо развитыми радулами .
Некоторые виды хитонов являются хищными , например, небольшой западно- тихоокеанский вид Placiphorella velata . Эти хищные хитоны имеют увеличенные передние пояса. Они ловят других мелких беспозвоночных , таких как креветки и, возможно, даже мелкую рыбу, удерживая увеличенный, похожий на капюшон передний конец пояса над поверхностью, а затем зажимая ничего не подозревающую и ищущую укрытие добычу. [36]
Размножение и жизненный цикл
[ редактировать ]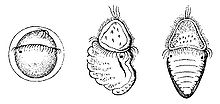
Хитоны раздельнополы, оплодотворение обычно наружное . Самец выпускает сперму в воду, а самка выпускает икру либо по отдельности, либо длинной цепочкой. В большинстве случаев оплодотворение происходит либо в окружающей воде, либо в мантийной полости самки. Некоторые виды высиживают яйца в мантийной полости, а вид Callistochiton viviparus даже сохраняет их в яичнике и рождает живых детенышей, что является примером яйцеживорождения .
Яйцо имеет жесткую колючую оболочку, из него обычно вылупляется свободно плавающая личинка трохофора , что типично для многих других групп моллюсков. В некоторых случаях трохофор остается внутри яйца (и тогда его называют лецитотрофным — получающим питание из желтка), из которого вылупляется миниатюрная взрослая особь. нет промежуточной стадии, или велигера В отличие от большинства других моллюсков, между трохофором и взрослой особью . Вместо этого на одной стороне личинки образуется сегментированная панцирная железа, а на противоположной стороне – ножка. Когда личинка готова стать взрослой, тело удлиняется, и панцирная железа выделяет пластинки панциря. В отличие от взрослой взрослой особи, у личинки есть пара простых глаз, хотя они могут оставаться некоторое время у неполовозрелой взрослой особи. [18]
Хищники
[ редактировать ]Хитонами питаются люди, чайки , морские звезды , крабы , омары и рыбы . [ нужна ссылка ]
Эволюционное происхождение
[ редактировать ]Хитоны имеют относительно хорошую летопись окаменелостей, начиная с кембрия. [1] [2] с родом Preacanthochiton , известным по окаменелостям, найденным в позднекембрийских отложениях в штате Миссури , который классифицируется как самый ранний известный полиплакофоран. Однако точное филогенетическое положение предполагаемых кембрийских хитонов весьма спорно, и вместо этого некоторые авторы утверждают, что самые ранние подтвержденные полиплакофоры относятся к раннему ордовику . [37] Кимберелла и Виваксия докембрия и кембрия могут быть связаны с предковыми полиплакофорами. Маттевия - полиплакофор позднего кембрия, сохранившийся в виде отдельных заостренных створок, иногда его рассматривают как хитон. [1] хотя, в лучшем случае, это может быть только член стволовой группы группы. [38]

Основываясь на этой и сопутствующих окаменелостях, одна из правдоподобных гипотез происхождения полиплакофор состоит в том, что они образовались, когда родился аберрантный моноплакофоран с несколькими центрами кальцификации, а не с обычным. Отбор быстро воздействовал на полученные конические оболочки, формируя из них защитную броню; их оригинальные шишки гомологичны кончикам пластинок современных хитонов. [1]
Хитоны произошли от мультиплакофор в палеозое, а их относительно консервативный современный план тела был зафиксирован в мезозое. [38]
Самые ранние ископаемые свидетельства присутствия эстетов в хитонах датируются примерно 400 млн лет назад, в раннем девоне . [19]
История научных исследований
[ редактировать ]Хитоны были впервые изучены Карлом Линнеем в его 10-м издании «Системы природы» 1758 года . С момента описания первых четырех видов хитоны были классифицированы по-разному. В начале 19 века их называли циклобраранхами они сгруппировались с аплакофорами в подтипе Amphineura (круглыми руками), а затем в 1876 году . Класс Polyplacophora был назван де Бленвиллем в 1816 году.
Этимология
[ редактировать ]Название хитон имеет неолатинское происхождение от древнегреческого слова «хитон» , что означает «туника» (от которого также произошло слово «хитин» ). Древнегреческое слово «хитон» происходит от центрально-семитского слова *киттан , которое происходит от аккадских слов «киту» или «кита'ум» , означающих лен или лен, и первоначально шумерского слова «гада» или «гида» . [ нужна ссылка ]
Греческое название «Полиплакофора» происходит от слов «поли-» («много»), «плако-» («таблетка») и «-форос» («несущий»), что указывает на восемь пластин панциря хитона.
Таксономия
[ редактировать ]Большинство схем классификации, используемых сегодня, основаны, по крайней мере частично, на «Руководстве по конхологии» Пилсбри (1892–1894), расширенном и исправленном Каасом и Ван Беллем (1985–1990).
С тех пор как хитоны были впервые описаны Линнеем (1758), были проведены обширные таксономические исследования на видовом уровне. Однако таксономическая классификация на более высоких уровнях группы осталась несколько неурегулированной.
Самая последняя классификация Сиренко (2006): [39] основан не только на морфологии скорлупы, как обычно, но и на других важных особенностях, включая эстетику , поясок, радулу, жабры, железы, выступы скорлупы яйца и сперматозоиды. В него входят все ныне живущие и вымершие роды хитонов.
Дальнейшее разрешение хитонид было восстановлено с помощью молекулярного анализа. [40]
Эта система сейчас общепринята.
- Класс Polyplacophora de Blainville, 1816 г.
- † Подкласс Paleoloricata Bergenhayn, 1955 г.
- † Орден Челодида Бергенхайн, 1943 г.
- † Семейство Chelodidae Bergenhayn, 1943.
- † Челодес Дэвидсон и Кинг, 1874 г.
- † Эвхелодес Марек, 1962 г.
- † Кальцеохитона , 1968 г. Цветок
- † Семейство Chelodidae Bergenhayn, 1943.
- † Орден Септемхитонида Бергенхайн, 1955 г.
- † Семейство Gotlandochitonidae Bergenhayn, 1955.
- † Готландохитон Бергенхайн, 1955 г.
- † Семейство Helminthochitonidae Van Belle, 1975.
- † Киндбладохитон Ван Белль, 1975 г.
- † Диаделохитон Хоар, 2000 г.
- † Гельминтохитон Солтер в Griffith & M'Coy, 1846 г.
- † Эхинохитон Пожета, Эрнисс, Хоар и Хендерсон, 2003 г.
- † Семейство Septemchitonidae Bergenhayn, 1955.
- † Септемхитон Бергенхайн, 1955 г.
- † Палеохитон А.Г. Смит, 1964 г.
- † Тайроплакс Чернс, 1998 г.
- † Семейство Gotlandochitonidae Bergenhayn, 1955.
- † Орден Челодида Бергенхайн, 1943 г.
- Подкласс Лориката Шумахер, 1817 г.
- Орден Лепидоплевриды Тиле, 1910 г.
- Подотряд Cymatochitonina Sirenko & Starobogatov, 1977.
- Семейство Acutichitonidae Hoare, Mapes & Atwater, 1983 г.
- Акутичитон Хоар, Осетр и Хоар, 1972 г.
- Элахихитон Хоар, Осетр и Хоар, 1972 г.
- Гарпидохитон Хоар и Кук, 2000 г.
- Аркохитон Хоар, Осетр и Хоар, 1972 г.
- Кратерохитон Хоар, 2000 г.
- Солеахитон Хоар, Осетр и Хоар, 1972 г.
- Аскетохитон Хоар и Сабаттини, 2000 г.
- Семейство † Cymatochitonidae Сиренко, Старобогатов, 1977.
- † Циматохитон Далл , 1882 г.
- † Компсохитон Хоар и Кук, 2000 г.
- Семейство Gryphochitonidae Pilsbry , 1900.
- Грифохитон Грей, 1847 г.
- Семейство Lekiskochitonidae Smith & Hoare, 1987.
- Лекискохитон Хоар и Смит, 1984 г.
- † Family Permochitonidae Sirenko & Starobogatov, 1977
- † Пермохитон Иредейл и Халл, 1926 г.
- Семейство Acutichitonidae Hoare, Mapes & Atwater, 1983 г.
- Подотряд Lepidopleurina Thiele, 1910 г.
- Семейство Abyssochitonidae (синоним: Ferreiraellidae) Dell 'Angelo & Palazzi, 1991 г.
- Глафурохитон Раймонд, 1910 г.
- ? Пикнохитон Хоар, 2000 г.
- ? Хадрохитон Хоар, 2000 г.
- Феррейрелла Сиренко, 1988 г.
- † Семейство Glyptochitonidae Старобогатов и Сиренко, 1975.
- † Глиптохитон Коннинк, 1883 г.
- Семейство Leptochitonidae Dall , 1889 г.
- Колаптерохитон Хоар и Мейпс, 1985 г.
- Кориссохитон ДеБрок, Хоар и Мейпс, 1984 г.
- Proleptochiton Sirenko & Starobogatov, 1977
- Схематохитон Хоара, 2002 г.
- Птерохитон ( Карпентер М.С.) Далл , 1882 г.
- Лептохитон Грей, 1847 г.
- Парахитон Тиле, 1909 год.
- Теренохитон Иредейл , 1914 год.
- Трахиплевра Якель, 1900 г.
- Псевдоишнохитон Эшби, 1930 год.
- Лепидоплеврус Риссо , 1826 г.
- Ханлейелла Сиренко, 1973 г.
- Семейство † Camptochitonidae Сиренко, 1997.
- Кэмпточитон ДеБрок, Хоар и Мейпс, 1984 г.
- Педанохитон ДеБрок, Хоар и Мэйпс, 1984 г.
- Эулептохитон Хоар и Мейпс, 1985 г.
- Пилеохитон ДеБрок, Хоар и Мейпс, 1984 г.
- Чаулиохитон Хоар и Смит, 1984 г.
- Стегохитон Хоар и Смит, 1984 г.
- Семейство Nierstraszellidae Сиренко, 1992 г.
- Ниерстразелла Сиренко, 1992 г.
- Семейство Mesochitonidae Dell 'Angelo & Palazzi, 1989.
- Мезохитон Ван Белль, 1975 г.
- Птеригохитон Рошбрюн, 1883 г.
- Семейство Protochitonidae Эшби, 1925 г.
- Протохитон Эшби, 1925 год.
- Дешайезиелла ( Карпентер М.С.) Далл , 1879 г.
- Олдройдия Далл , 1894 г.
- Семейство Hanleyidae Bergenhayn, 1955 г.
- Ханлея Грей, 1857 г.
- Гемиартрум Далл , 1876 г.
- Семейство Abyssochitonidae (синоним: Ferreiraellidae) Dell 'Angelo & Palazzi, 1991 г.
- Подотряд Cymatochitonina Sirenko & Starobogatov, 1977.
- Орден Хитониды Тиле, 1910 г.
- Подотряд Chitonina Thiele, 1910 г.
- Надсемейство Chitonoidea Rafinesque , 1815 г.
- Семейство Ochmazochitonidae Hoare & Smith, 1984.
- Охмазохитон Хоар и Смит, 1984 г.
- Семейство Ischnochitonidae Dall , 1889.
- Ищнохитон Грей, 1847 г.
- Стенохитон Х. Адамс и Ангас, 1864 г.
- Стеноплакс ( Карпентер М.С. ) Далл , 1879 г.
- Лепидозона Пилсбри , 1892 г.
- Стеноз Миддендорфа, 1847 г.
- Субтеренохитон Иредейл и Халл, 1924 год.
- Термохитон Сайто и Окутани, 1990 г.
- Сыр коннексохитон , 1979 г.
- Тоницина Тиле, 1906 год.
- Семейство Callistoplacidae Pilsbry , 1893 г.
- Ишноплакс Далл , 1879 г.
- Каллистохитон Карпентер М.С., Далл , 1879 г.
- Каллистоплакс Далл , 1882 г.
- Цератозона Далл , 1882 г.
- Каллоплакс Тиле, 1909 год.
- семейства Chaetopleuridae , 1899 г. Тарелка
- Четоплевра Шаттлворт, 1853 г.
- Диноплакс Карпентер М.С., Далл , 1882 г. [41]
- Семья Loricidae Iredale & Hull, 1923 г.
- Тарелка семейства Callochitonidae , 1901 г.
- Каллохитон Грей, 1847 г.
- Евдоксохитон Шаттлворт, 1853 г.
- Сыр Вермихитон , 1979 год.
- Семейство Chitonidae Rafinesque , 1815 г.
- Подсемейство Chitoninae Rafinesque , 1815 г.
- Хитон Линней , 1758 г.
- Амаурохитон Тиле, 1893 г.
- Радсия Грей, 1847 г.
- Сифарохитон Тиле, 1893 г.
- Нодиплакс Бью, 1967 г.
- Риссоплакс Тиле, 1893 г.
- Тегулоаплакс Иредейл и Халл, 1926 год.
- Макросквама Иредейл , 1893 г.
- Подсемейство Toniciinae Pilsbry , 1893.
- Тоника Грей, 1847 г.
- Онитохитон Грей, 1847 г.
- Подсемейство Acanthopleurinae Dall , 1889.
- Гильдия Акантоплевры , 1829 г.
- Лиолофура Пилсбри , 1893 г.
- Эноплохитон Грей, 1847 г.
- Сквамоплевра Нирстраз, 1905 г.
- Подсемейство Chitoninae Rafinesque , 1815 г.
- Семейство Ochmazochitonidae Hoare & Smith, 1984.
- Надсемейство Schizochitonoidea Dall , 1889 г.
- Семейство Schizochitonidae Dall , 1889 г.
- Инциссиохитон Ван Белль, 1985 г.
- Шизохитон Грей, 1847 г.
- Семейство Schizochitonidae Dall , 1889 г.
- Надсемейство Chitonoidea Rafinesque , 1815 г.
- Подотряд Acanthochitonina Bergenhayn, 1930 г.
- Надсемейство Mopalioidea Dall , 1889 г.
- Семейство Tonicellidae Simroth, 1894 г.
- Подсемейство Tonicellinae Simroth, 1894 г.
- Лепидохитона Грей, 1821 г.
- Сыр Особаязона , 1993 г.
- Бореохитон Сарс, 1878 г.
- Тоникелла Карпентер , 1873 г.
- Нутталлина ( Карпентер М.С.) Далл, 1871 г.
- Спонгиорадсия Пилсбри , 1894 г.
- Олигохитон Берри, 1922 г.
- Подсемейство Juvenichitoninae Сиренко, 1975.
- Ювенихитон Сиренко, 1975 г.р.
- Мичичитон Сиренко, 1975 г.
- Наничитон Сиренко, 1975 г.
- Подсемейство Tonicellinae Simroth, 1894 г.
- Семейство Schizoplacidae Bergenhayn, 1955 г.
- Семейство Mopaliidae Dall , 1889 г.
- Подсемейство Heterochitoninae Van Belle, 1978.
- Гетерохитон Фучини, 1912 г.
- Аллохитон Фучини, 1912 г.
- Подсемейство Mopaliinae Dall , 1889 г.
- Аэриламма Халл, 1924 год.
- Гильдингия Пилсбри , 1893 г.
- Фремблея Х. Адамс, 1866 г.
- Диафороплакс Иредейл , 1914 г.
- Плаксифора Грей, 1847 г.
- Плацифорина Каас и Ван Белль, 1994 г.
- Плита Нутталлохитон , 1899 г.
- Мопалия Грей, 1847 г.
- Маорихитон Иредейл , 1914 год.
- Плацифорелла ( Карпентер М.С.) Далл , 1879 г.
- Катарина Грей, 1847 г.
- Амикула Грей, 1847 г.
- Подсемейство Heterochitoninae Van Belle, 1978.
- Семейство Tonicellidae Simroth, 1894 г.
- Надсемейство Cryptoplacoidea H. & A. Adams , 1858 г.
- Семейство Acanthochitonidae Pilsbry , 1893.
- Подсемейство Acanthochitoninae Pilsbry , 1893.
- Акантохитона Грей, 1921 год.
- Краспедохитон Шаттлворт, 1853 г.
- Губгиохитон ( Карпентер М.С.) Далл , 1882 г.
- Нотоплакс Х. Адамс, 1861 г.
- Псевдотоника Эшби, 1928 г.
- Бассетуллия Пилсбри , 1928 год.
- Америхитон Уоттерс, 1990 г.
- Чонеплакс ( Карпентер М.С.) Далл , 1882 г.
- Криптоконх ( де Бленвиль М.С.) Берроу, 1815 г.
- Подсемейство Cryptochitoninae Pilsbry , 1893 г.
- Криптохитон Миддендорф, 1847 г.
- Подсемейство Acanthochitoninae Pilsbry , 1893.
- Семейство Hemiarthridae Сиренко, 1997 г.
- Hemiarthrum Carpenter в Далле , 1876 г.
- Видингия Каас, 1988 год.
- Семейство Choriplacidae Ashby, 1928 г.
- Семейство Cryptoplacidae H. & A. Adams , 1858 г.
- Криптоплакс из Бленвилля , 1818 г.
- Семейство Acanthochitonidae Pilsbry , 1893.
- Надсемейство Mopalioidea Dall , 1889 г.
- Подотряд Chitonina Thiele, 1910 г.
- Орден Лепидоплевриды Тиле, 1910 г.
- Неуверенное сиденье
- Семейство Scanochitonidae Bergenhayn, 1955.
- Сканохитон Бергенхайн, 1955 год.
- Семейство Olingechitonidae Старобогатов и Сиренко, 1977.
- Олингечитон Бергенхайн, 1943 год.
- Семейство Haeggochitonidae Сиренко и Старобогатов, 1977.
- Хэггохитон Бергенхайн, 1955 год.
- Семейство Ivoechitonidae Сиренко и Старобогатов, 1977.
- Ивоехитон Бергенхайн, 1955 год.
- Семейство Scanochitonidae Bergenhayn, 1955.
- † Подкласс Paleoloricata Bergenhayn, 1955 г.
Ссылки
[ редактировать ]- ^ Jump up to: а б с д Раннегар, Б.; Пожета, младший (октябрь 1974 г.). «Филогения моллюсков: палеонтологическая точка зрения». Наука . 186 (4161): 311–317. Бибкод : 1974Sci...186..311R . дои : 10.1126/science.186.4161.311 . JSTOR 1739764 . ПМИД 17839855 . S2CID 46429653 .
- ^ Jump up to: а б Чернс, Лесли (2 января 2007 г.). «Раннепалеозойская диверсификация хитонов (Polyplacophora, Mollusca) на основе новых данных из силура Готланда, Швеция» . Летайя . 37 (4): 445–456. дои : 10.1080/00241160410002180 . Проверено 25 ноября 2022 г.
- ^ Jump up to: а б Чисхолм, Хью , изд. (1911). » энциклопедия Британская Том. 6 (11-е изд.). Издательство Кембриджского университета. стр. 100-1 247–251.
- ^ «Полиплакофора» . Интегрированная таксономическая информационная система .
- ^ Швабе, Э (2005). «Каталог современных и ископаемых хитонов (Mollusca: Polyplacophora)». Новапекс . 6 : 89–105.
- ^ Стеббинс, Т.Д.; Эрнисс, диджей (2009). «Хитоны (Mollusca: Polyplacophora), известные из программ мониторинга бентоса в заливе Южная Калифорния». Фестиваль . 41 : 53–100.
- ^ Пухальский, С.; Эрниссе, диджей; Джонсон, CC (2008). «Влияние систематической ошибки выборки на летопись окаменелостей хитонов (Mollusca, Polyplacophora)» . Американский малакологический бюллетень . 25 : 87–95. дои : 10.4003/0740-2783-25.1.87 . S2CID 59485784 . Архивировано из оригинала 26 июля 2020 года . Проверено 4 октября 2021 г.
- ^ Коннорс, MJ; Эрлих, Х.; Хог, М.; Годфруа, К.; Арайя, С.; Каллай, И.; Газит, Д.; Бойс, М.; Ортис, К. (2012). «Трехмерная структура сборки панцирной пластинки хитона Tonicella Marmorea и ее биомеханические последствия». Журнал структурной биологии . 177 (2): 314–328. дои : 10.1016/j.jsb.2011.12.019 . ПМИД 22248452 .
- ^ Линдберг, Дэвид Р. (1 июня 2009 г.). «Моноплакофоры, происхождение и родство моллюсков» . Эволюция: образование и информационно-пропагандистская деятельность . 2 (2): 191–203. дои : 10.1007/s12052-009-0125-4 . ISSN 1936-6434 . S2CID 26108547 .
- ^ Винтер, Дж.; Нильсен, К. (2005). «Раннекембрийские халкиерии — моллюски». Зоологика Скрипта . 34 : 81–89. дои : 10.1111/j.1463-6409.2005.00177.x . S2CID 84493997 .
- ^ Jump up to: а б с д и ж Тревес, К.; Трауб, В.; Вайнер, С.; Аддади, Л. (2003). «Арагонитовые образования в поясе хитона (моллюска)». Helvetica Chimica Acta . 86 (4): 1101–1112. дои : 10.1002/hlca.200390096 .
- ^ Jump up to: а б П. Дж. Хейворд и Дж. С. Райланд (1996). Справочник морской фауны Северо-Западной Европы . Издательство Оксфордского университета. п. 485. ИСБН 978-0-19-854055-7 .
- ^ Филиал, генеральный директор; Гриффитс, CL; Бранч, М.Л. и Бекли, Ю.Л. (2010). Два океана: Путеводитель по морской жизни Южной Африки . Кейптаун: Природа Струика.
- ^ Джейкобс, ДК; Рэй, К.Г.; Ведин, CJ; Кострикен, Р.; Десаль, Р.; Стейтон, Дж.Л.; Гейтс, РД; Линдберг, Д.Р. (2000). «Закрепленное выражение моллюсков, серийная организация и эволюция раковины». Эволюция и развитие . 2 (6): 340–347. дои : 10.1046/j.1525-142x.2000.00077.x . ПМИД 11256378 . S2CID 25274057 .
- ^ Jump up to: а б Хаас, В. (1981). «Эволюция известковых твердых частей у примитивных моллюсков малакология». Малакология . 21 : 403–418.
- ^ Jump up to: а б с Генри, Дж.; Окусу, А.; Мартиндейл, М. (2004). «Клеточная линия полиплакофорана Chaetopleura apiculata: вариации спиральной программы и значение для эволюции моллюсков» . Биология развития . 272 (1): 145–160. дои : 10.1016/j.ydbio.2004.04.027 . ПМИД 15242797 .
- ^ Animalnetwork.com. Архивировано 16 марта 2003 г. на Wayback Machine.
- ^ Jump up to: а б с д и ж г Барнс, Роберт Д. (1982). Зоология беспозвоночных . Филадельфия, Пенсильвания: Холт-Сондерс Интернэшнл. стр. 381–389. ISBN 978-0-03-056747-6 .
- ^ Jump up to: а б с Серб, Ю.М.; Эрнисс, диджей (2008). «Изложение траектории эволюции: использование разнообразия глаз моллюсков для понимания параллельной и конвергентной эволюции» . Эволюция: образование и информационно-пропагандистская деятельность . 1 (4): 439–447. дои : 10.1007/s12052-008-0084-1 .
- ^ Джеймс Х. Маклин (1 апреля 1962 г.). «Пищевое поведение хитонов Placiphorella» . Журнал исследований моллюсков . 35 (1): 23. Архивировано из оригинала 19 июля 2012 года.
- ^ Спейзер, Д.И.; Эрниссе, диджей; Джонсен, С.Н. (2011). «Хитон использует линзы из арагонита для формирования изображений» . Современная биология . 21 (8): 665–670. Бибкод : 2011CBio...21..665S . дои : 10.1016/j.cub.2011.03.033 . ПМИД 21497091 . S2CID 10261602 .
- ^ Ли, Л.; Коннорс, MJ; Колле, М.; Англия, GT; Спейзер, Д.И.; Сяо, X.; Айзенберг, Дж.; Ортис, К. (2015). «Многофункциональность хитоновой биоминерализованной брони с интегрированной зрительной системой» (PDF) . Наука . 350 (6263): 952–956. дои : 10.1126/science.aad1246 . hdl : 1721.1/100035 . ПМИД 26586760 . S2CID 217544572 . Архивировано (PDF) из оригинала 4 ноября 2018 года . Проверено 4 ноября 2018 г.
- ^ «Странный морской моллюск с сотнями глаз, сделанных из доспехов» . Живая наука . 19 ноября 2015 года. Архивировано из оригинала 17 августа 2016 года . Проверено 28 июля 2016 г.
- ^ «Глаза, сделанные из камня, действительно могут видеть, говорят исследования» . 14 апреля 2011 года. Архивировано из оригинала 20 декабря 2013 года . Проверено 10 декабря 2013 г.
- ^ «Хитоны видят керамическими глазами, показывают новые исследования» . Архивировано из оригинала 15 мая 2019 года . Проверено 15 мая 2019 г.
- ^ Сигварт, доктор медицинских наук; Самнер-Руни, Л.Х.; Швабе, Э.; Хесс, М.; Бреннан, врач общей практики; Шредль, М. (2014). «Новый орган чувств у примитивных моллюсков (Polyplacophora: Lepidopleurida) и его контекст в нервной системе хитонов» . Границы в зоологии . 11 (1): 7. дои : 10.1186/1742-9994-11-7 . ПМЦ 3916795 . ПМИД 24447393 .
- ^ Самнер-Руни, Л.Х.; Сигварт, доктор юридических наук (2015). «Является ли орган Швабе сохранившимся личиночным глазом? Анатомические и поведенческие исследования нового органа чувств у взрослых Leptochiton asellus (Mollusca, Polyplacophora) указывают на связь с личиночными фоторецепторами» . ПЛОС ОДИН . 10 (9): e0137119. Бибкод : 2015PLoSO..1037119S . дои : 10.1371/journal.pone.0137119 . ПМК 4569177 . ПМИД 26366861 .
- ^ (Thorne. J. M, 1968; Moroz. L, et al., 1993).
- ^ Челацци; и др. (1983). «Сравнительное исследование характера движения двух симпатрических тропических хитонов Mollusca: Polyplacophora» . Морская биология . 74 (2): 115–125. Бибкод : 1983MarBi..74..115C . дои : 10.1007/bf00413914 . S2CID 56141764 . ; Челацци, Дж; и др. (1990). «Роль следования по следу в возвращении хитонов приливной зоны: сравнение трех видов Acanthopleura». Морская биология . 105 (3): 445–450. Бибкод : 1990МарБи.105..445С . дои : 10.1007/bf01316316 . S2CID 83889350 .
- ^ Jump up to: а б (Челацци Г. и др., 1987; Торн Дж. М., 1968).
- ^ (Челацци, Г. и др., 1985).
- ^ Киршвинк, Дж.Л.; Лоуэнстам, штат Джорджия (1 августа 1979 г.). «Минерализация и намагниченность хитоновых зубов: палеомагнитные, седиментологические и биологические последствия органического магнетита». Письма о Земле и планетологии . 44 (2): 193–204. Бибкод : 1979E&PSL..44..193K . дои : 10.1016/0012-821X(79)90168-7 . ISSN 0012-821X .
- ^ Самнер-Руни, Лорен Х.; Мюррей, Джеймс А.; Каин, Шон Д.; Сигварт, Джулия Д. (2014). «Есть ли у хитонов компас? Доказательства магнитной чувствительности полиплакофоры». Журнал естественной истории . 48 (45–48): 45–48. Бибкод : 2014JNatH..48.3033S . дои : 10.1080/00222933.2014.959574 . S2CID 84896224 .
- ^ Кангас, Мерви; Шеперд, ЮАР (1984). «Распространение и питание хитонов в среде обитания валунов на Западном острове, Южная Австралия». Журнал Малакологического общества Австралии . 6 (3–4): 101–111. дои : 10.1080/00852988.1984.10673963 .
- ^ Барнавелл, Э.Б. (1960). Плотоядность полиплакофоры.
- ^ «Реакция на кормление Placiphorella velata» . Ютуб . Архивировано из оригинала 18 октября 2020 года . Проверено 17 октября 2020 г.
- ^ Сигварт, доктор медицинских наук; Саттон, доктор медицины (октябрь 2007 г.). «Глубинная филогения моллюсков: синтез палеонтологических и неонтологических данных» . Труды Королевского общества B: Биологические науки . 274 (1624): 2413–2419. дои : 10.1098/rspb.2007.0701 . ПМК 2274978 . ПМИД 17652065 . Краткое описание см. «Моллюска» . Музей палеонтологии Калифорнийского университета. Архивировано из оригинала 15 декабря 2012 года . Проверено 2 октября 2008 г.
- ^ Jump up to: а б Вендраско, MJ; Вуд, TE; Раннегар, Б.Н. (2004). «Сочлененная палеозойская окаменелость с 17 пластинами значительно расширяет неравенство ранних хитонов». Природа . 429 (6989): 288–291. Бибкод : 2004Natur.429..288V . дои : 10.1038/nature02548 . ПМИД 15152250 . S2CID 4428441 .
- ^ Сиренко Б.И. (2006). «Новый взгляд на систему хитонов (Mollusca: Polyplacophora)». Венера . 65 (1–2): 27–49.
- ^ Сигварт, Джулия Д.; Штегер, Изабелла; Кнебельсбергер, Томас; Швабе, Энрико (2013). «Филогения хитона (Mollusca: Polyplacophora) и размещение загадочного вида Choriplax Grayi (H. Adams & Angas) » . Систематика беспозвоночных . 27 (6): 603. дои : 10.1071/IS13013 . S2CID 86845236 . [ постоянная мертвая ссылка ]
- ^ «WoRMS — Всемирный реестр морских видов» . www.marinespecies.org . Архивировано из оригинала 29 мая 2010 года . Проверено 7 апреля 2010 г.
Внешние ссылки
[ редактировать ]| Базы данных органов управления : Национальные |
|---|