Круговая перестановка в белках

Циклическая перестановка — это взаимосвязь между белками , при которой в белках изменяется порядок аминокислот в их пептидной последовательности . В результате получается белковая структура с различной связностью, но в целом схожая трехмерная (3D) форма. первая пара белков с циркулярной перестановкой — конканавалин А и лектин В 1979 году была открыта ; Сейчас известно более 2000 таких белков.
Круговая перестановка может происходить в результате эволюционных событий, посттрансляционных модификаций или искусственно созданных мутаций. Двумя основными моделями, предложенными для объяснения эволюции белков с циркулярной перестановкой, являются пермутация путем дупликации , а также деление и слияние . Перестановка путем дупликации происходит, когда ген подвергается дупликации с образованием тандемного повтора до удаления избыточных участков белка; эта связь обнаружена между сапозином и свопозином. Деление и слияние происходят, когда частичные белки сливаются с образованием одного полипептида, например, в трансгидрогеназах никотинамиднуклеотидов .
Круговые перестановки обычно разрабатываются в лаборатории для улучшения их каталитической активности или термостабильности или для исследования свойств исходного белка.
Традиционные алгоритмы выравнивания последовательностей и структур не способны обнаруживать циклические перестановки между белками. Были разработаны новые нелинейные подходы, которые преодолевают эту проблему и способны обнаруживать сходства, независимые от топологии .
История
[ редактировать ]
В 1979 году Брюс Каннингем и его коллеги обнаружили первый в природе экземпляр белка с циркулярной перестановкой. [ 1 ] Определив пептидную последовательность лектинового белка фавина, они заметили его сходство с известным белком – конканавалином А – за исключением того, что концы были переставлены по кругу. Более поздняя работа подтвердила круговую перестановку между парой. [ 2 ] и показали, что конканавалин А подвергается посттрансляционной перестановке. [ 3 ] путем расщепления и необычного лигирования белка. [ 4 ]
После открытия природного белка с циклической перестановкой исследователи начали искать способ имитировать этот процесс. В 1983 году Дэвид Гольденберг и Томас Крейтон смогли создать версию белка с циркулярной перестановкой путем химического лигирования концов для создания циклического белка , а затем введения новых концов в другое место с помощью трипсина . [ 5 ] В 1989 году Каролин Люгер и ее коллеги представили генетический метод создания кольцевых перестановок путем тщательного фрагментирования и лигирования ДНК. [ 6 ] Этот метод позволял вводить перестановки в произвольных местах. [ 6 ]
Несмотря на раннее открытие посттрансляционных кольцевых перестановок и предположение о возможном генетическом механизме развития кольцевых пермутантов, только в 1995 году была открыта первая пара генов с кольцевыми перестановками. Сапозины представляют собой класс белков, участвующих в сфинголипидов и презентации антигенов липидов катаболизме у человека. Крис Понтинг и Роберт Рассел идентифицировали циркулярно перестановочную версию сапозина, встроенную в аспарагиновую протеиназу растений , которую они назвали свопозин . [ 7 ] Сапозин и свопозин были первым известным случаем двух природных генов, связанных кольцевой перестановкой. [ 7 ]
Сотни примеров пар белков, связанных кольцевой перестановкой, были впоследствии обнаружены в природе или получены в лаборатории. По состоянию на февраль 2012 г. база данных циклических перестановок [ 8 ] содержит 2238 пар белков с известной структурой, перестановленных по кругу, и многие другие известны без структур. [ 9 ] В базе данных CyBase собраны циклические белки, некоторые из которых представляют собой пермутированные варианты циклических белков дикого типа. [ 10 ] SISYPHUS — это база данных, содержащая коллекцию вручную созданных выравниваний белков с нетривиальными отношениями, некоторые из которых имеют циклические перестановки. [ 11 ]
Эволюция
[ редактировать ]В настоящее время для объяснения эволюции белков с циркулярной перестановкой используются две основные модели: перестановка путем дупликации , а также деление и слияние . Обе модели имеют убедительные примеры, подтверждающие их, но относительный вклад каждой модели в эволюцию все еще обсуждается. [ 12 ] Были предложены и другие, менее распространенные механизмы, такие как «вырезать и вставить». [ 13 ] или « перетасовка экзонов ». [ 14 ]
Перестановка путем дублирования
[ редактировать ]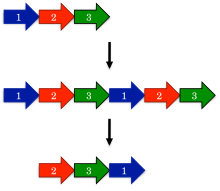
Самая ранняя модель, предложенная для эволюции круговых перестановок, - это перестановка по механизму дублирования. [ 1 ] В этой модели ген-предшественник сначала подвергается дупликации и слиянию с образованием большого тандемного повтора . Затем стартовый и стоп-кодоны вводятся в соответствующие места дуплицированного гена, удаляя избыточные участки белка.
Одним из удивительных предсказаний механизма перестановок посредством дублирования является то, что могут возникать промежуточные перестановки. Например, дублированная версия белка должна оставаться функциональной, поскольку в противном случае эволюция быстро отберет такие белки. Аналогично, частично дублированные промежуточные соединения, в которых был усечен только один конец, должны быть функциональными. Такие промежуточные соединения широко описаны в семействах белков, таких как ДНК-метилтрансферазы . [ 15 ]
Сапозин и свопозин
[ редактировать ]
Примером перестановки путем дупликации являются отношения между сапозином и свопозином. Сапозины представляют собой высококонсервативные гликопротеины длиной около 80 аминокислотных остатков, образующие четырехальфа -спиральную структуру. Они имеют почти идентичное расположение остатков цистеина и сайтов гликозилирования. Последовательность кДНК , кодирующая сапозин, называется просапозином . Он является предшественником четырех продуктов расщепления: сапозинов A, B, C и D. Четыре домена сапозина, скорее всего, возникли в результате двух тандемных дупликаций предкового гена. [ 16 ] Этот повтор предполагает механизм эволюции взаимоотношений со специфичной для растения вставкой (PSI). PSI представляет собой домен, обнаруженный исключительно в растениях, состоящий примерно из 100 остатков и обнаруженный в растительных аспарагиновых протеазах . [ 17 ] Он принадлежит к семейству сапозиноподобных белков (SAPLIP) и имеет «поменянные местами» N- и C-концы, так что порядок спиралей составляет 3-4-1-2 по сравнению с сапозином, что привело к названию «свапозин». ". [ 7 ] [ 18 ]
Деление и синтез
[ редактировать ]
Другая модель эволюции круговых перестановок — это модель деления и синтеза. Процесс начинается с двух частичных белков. Они могут представлять собой два независимых полипептида (например, две части гетеродимера ) или изначально представлять собой половинки одного белка, который подвергся делению и превратился в два полипептида.
Два белка позже могут сливаться вместе, образуя один полипептид. Независимо от того, какой белок стоит первым, этот слитый белок может выполнять аналогичную функцию. Таким образом, если слияние двух белков происходит в эволюции дважды (либо между паралогами внутри одного вида, либо между ортологами разных видов), но в разном порядке, полученные слитые белки будут связаны круговой перестановкой.
Доказательства того, что конкретный белок эволюционировал по механизму деления и слияния, могут быть получены путем наблюдения за половинами перестановки как независимыми полипептидами у родственных видов или путем экспериментальной демонстрации того, что две половины могут функционировать как отдельные полипептиды. [ 19 ]
Трансгидрогеназы
[ редактировать ]Пример механизма деления и слияния можно найти в трансгидрогеназах никотинамиднуклеотидов . [ 20 ] Это мембраносвязанные , ферменты которые катализируют перенос гидрид-иона между НАД(Н) и НАДФ(Н) в реакции, связанной с трансмембранной транслокацией протонов . Они состоят из трех основных функциональных единиц (I, II и III), которые можно найти в различном расположении у бактерий , простейших и высших эукариот . Филогенетический анализ предполагает, что три группы расположения доменов были приобретены и слиты независимо. [ 12 ]
Другие процессы, которые могут привести к круговым перестановкам
[ редактировать ]Посттрансляционная модификация
[ редактировать ]Две упомянутые выше эволюционные модели описывают способы, с помощью которых гены могут подвергаться циркулярной перестановке, что приводит к образованию циркулярно перестановленной мРНК после транскрипции . Белки также могут подвергаться циклической перестановке посредством посттрансляционной модификации без изменения основного гена. Циклические перестановки могут происходить спонтанно посредством автокатализа , как в случае конканавалина А. [ 4 ] С другой стороны, для перестановки могут потребоваться ферменты рестрикции и лигазы . [ 5 ]
Роль в белковой инженерии
[ редактировать ]Концы многих белков расположены близко друг к другу в трехмерном пространстве. [ 21 ] [ 22 ] Благодаря этому часто можно создавать круговые перестановки белков. Сегодня круговые перестановки регулярно генерируются в лаборатории с использованием стандартных генетических методов. [ 6 ] белка Хотя некоторые сайты перестановок препятствуют правильному сворачиванию , было создано множество пермутантов с почти идентичной структурой и функциями исходному белку.
Мотивация создания кольцевого пермутанта белка может быть разной. Ученые могут захотеть улучшить некоторые свойства белка, такие как:
- Снижение протеолитической восприимчивости. Скорость расщепления белков может иметь большое влияние на их активность в клетках. Поскольку концы часто доступны для протеаз , создание белка с циркулярной перестановкой и менее доступными концами может увеличить продолжительность жизни этого белка в клетке. [ 23 ]
- Улучшить каталитическую активность . Циклическая перестановка белка иногда может увеличить скорость, с которой он катализирует химическую реакцию, что приводит к получению более эффективных белков. [ 24 ]
- Изменить связывание субстрата или лиганда . Циклическая перестановка белка может привести к потере связывания субстрата , но иногда может приводить к новой активности связывания лиганда или изменению специфичности субстрата. [ 25 ]
- Улучшите термостабильность . Повышение активности белков в более широком диапазоне температур и условий может повысить их полезность. [ 26 ]
С другой стороны, ученых могут заинтересовать свойства исходного белка, такие как:
- Порядок складывания. Определение порядка, в котором сворачиваются различные части белка, является сложной задачей из-за чрезвычайно быстрых временных масштабов. Версии белков с циркулярной перестановкой часто сворачиваются в другом порядке, предоставляя информацию о сворачивании исходного белка. [ 27 ] [ 28 ] [ 29 ]
- Основные конструктивные элементы. Искусственные белки с циркулярной перестановкой могут позволить избирательно удалять части белка. Это дает представление о том, какие структурные элементы являются существенными, а какие нет. [ 30 ]
- Изменить четвертичную структуру . Было показано, что белки с циркулярной перестановкой имеют четвертичную структуру, отличную от белков дикого типа. [ 31 ]
- Найдите места вставки других белков. Вставка одного белка в качестве домена в другой белок может оказаться полезной. Например, вставка кальмодулина в зеленый флуоресцентный белок (GFP) позволила исследователям измерить активность кальмодулина по флуоресценции расщепленного GFP. [ 32 ] Области GFP, которые допускают введение круговой перестановки, с большей вероятностью примут добавление другого белка, сохраняя при этом функцию обоих белков.
- Разработка новых биокатализаторов и биосенсоров. Введение круговых перестановок может быть использовано для создания белков, катализирующих определенные химические реакции. [ 24 ] [ 33 ] или для обнаружения присутствия определенных молекул с помощью белков. Например, описанное выше слияние GFP-кальмодулина можно использовать для определения уровня ионов кальция в образце. [ 32 ]
Алгоритмическое обнаружение
[ редактировать ]Многие алгоритмы выравнивания последовательностей и структур белков были разработаны с учетом линейного представления данных и, как таковые, не способны обнаруживать циклические перестановки между белками. [ 34 ] Двумя примерами часто используемых методов, которые имеют проблемы с правильным выравниванием белков, связанных циклической перестановкой, являются динамическое программирование и множество скрытых марковских моделей . [ 34 ] В качестве альтернативы этому ряд алгоритмов построен на основе нелинейных подходов и способен обнаруживать независимые от топологии сходства или использовать модификации, позволяющие им обойти ограничения динамического программирования. [ 34 ] [ 35 ] В таблице ниже представлена коллекция таких методов.
Алгоритмы классифицируются по типу требуемых входных данных. Алгоритмы, основанные на последовательностях, требуют только последовательности двух белков для создания выравнивания. [ 36 ] Методы секвенирования, как правило, быстры и подходят для поиска во всем геноме пар белков с циклической перестановкой. [ 36 ] Структурные методы требуют трехмерных структур обоих рассматриваемых белков. [ 37 ] Они часто медленнее, чем методы, основанные на последовательностях, но способны обнаруживать круговые перестановки между отдаленно родственными белками с низким сходством последовательностей. [ 37 ] Некоторые структурные методы не зависят от топологии , а это означает, что они также способны обнаруживать более сложные перестановки, чем круговые перестановки. [ 38 ]
| ИМЯ | Тип | Описание | Автор | Год | Доступность | Ссылка |
|---|---|---|---|---|---|---|
| ФБПЛОТ | Последовательность | Рисует точечные графики неоптимального выравнивания последовательностей. | Цукер | 1991 | [ 39 ] | |
| Бачар и др. | Структура, независимая от топологии | Использует геометрическое хеширование для независимого от топологии сравнения белков. | Бачар и др. | 1993 | [ 35 ] | |
| Улиэль и др. | Последовательность | Первое предложение о том, как может работать алгоритм сравнения последовательностей для обнаружения циклических перестановок. | Улиэль и др. | 1999 | [ 36 ] | |
| СМОТРЕТЬ | Структура | Использует алгоритм SHEBA для создания структурных выравниваний для различных точек перестановки, одновременно итеративно улучшая точку отсечения. | Юнг и Ли | 2001 | [ 14 ] | |
| Мультипрот | Структура, независимая от топологии | Вычисляет выравнивание структуры нескольких белков независимо от порядка последовательности. | Шацкий | 2004 | сервер, скачать | [ 38 ] |
| ПО РАСПРЕДЕЛЕНИЮ | Последовательность | Модифицированный алгоритм сравнения последовательностей Нидлмана и Вунша | Вайнер и др. | 2005 | скачать | [ 34 ] |
| ЦПСАРСТ | Структура | Описывает белковые структуры как одномерные текстовые строки с использованием алгоритма последовательного преобразования Рамачандрана (RST). Обнаруживает циклические перестановки посредством дублирования представления последовательности и стратегии «двойной фильтрации и уточнения». | Ло, Лю | 2008 | сервер | [ 40 ] |
| ГАНГСТА + | Структура | Работает в два этапа: на первом этапе определяются грубые выравнивания на основе элементов вторичной структуры. Второй этап уточняет выравнивание на уровне остатков и распространяется на области петель. | Шмидт-Геннер и др. | 2009 | сервер , скачать | [ 41 ] |
| МНОГО | Структура | Обнаружение первоначально выровненных пар фрагментов (AFP). Создайте сеть возможных AFP. Используйте алгоритм случайного совмещения для соединения компонентов в граф. | Ван и др. | 2010 | скачать | [ 42 ] |
| ИК-КП | Структура | Построен на основе алгоритма комбинаторного расширения . Дублирует атомы перед выравниванием, усекает результаты после выравнивания | Блайв и др. | 2015 | сервер , скачать | [ 43 ] |
| ТопМатч | Структура | Имеет возможность расчета выравнивания структуры белка, независимого от топологии. | Зиппл и Видерштейн | 2012 | сервер , скачать | [ 44 ] |
Ссылки
[ редактировать ]![]() Эта статья была адаптирована из следующего источника под лицензией CC BY 4.0 ( 2012 г. ) ( отчеты рецензента ):
Спенсер Бливен; Андреас Прлич (2012). «Круговая перестановка в белках» . PLOS Вычислительная биология . 8 (3): e1002445. doi : 10.1371/JOURNAL.PCBI.1002445 . ISSN 1553-734X . ПМК 3320104 . ПМИД 22496628 . Викиданные Q5121672 .
Эта статья была адаптирована из следующего источника под лицензией CC BY 4.0 ( 2012 г. ) ( отчеты рецензента ):
Спенсер Бливен; Андреас Прлич (2012). «Круговая перестановка в белках» . PLOS Вычислительная биология . 8 (3): e1002445. doi : 10.1371/JOURNAL.PCBI.1002445 . ISSN 1553-734X . ПМК 3320104 . ПМИД 22496628 . Викиданные Q5121672 .
- ^ Перейти обратно: а б с Каннингем Б.А., Хемперли Дж.Дж., Хопп Т.П., Эдельман Г.М. (июль 1979 г.). «Фавин против конканавалина А: аминокислотные последовательности с круговой перестановкой» . Труды Национальной академии наук Соединенных Штатов Америки . 76 (7): 3218–22. Бибкод : 1979PNAS...76.3218C . дои : 10.1073/pnas.76.7.3218 . ПМЦ 383795 . ПМИД 16592676 .
- ^ Эйншпар Х., Паркс Э.Х., Сугуна К., Субраманиан Э., Суддат Флорида (декабрь 1986 г.). «Кристаллическая структура горохового лектина при разрешении 3,0 А» . Журнал биологической химии . 261 (35): 16518–27. дои : 10.1016/S0021-9258(18)66597-4 . ПМИД 3782132 .
- ^ Кэррингтон Д.М., Оффрет А., Ханке Д.Е. (1985). «Лигирование полипептида происходит во время посттрансляционной модификации конканавалина А». Природа . 313 (5997): 64–7. Бибкод : 1985Natur.313...64C . дои : 10.1038/313064a0 . ПМИД 3965973 . S2CID 4359482 .
- ^ Перейти обратно: а б Боулз-ди-джей, Паппин-ди-джей (февраль 1988 г.). «Трафик и сборка конканавалина А». Тенденции биохимических наук . 13 (2): 60–4. дои : 10.1016/0968-0004(88)90030-8 . ПМИД 3070848 .
- ^ Перейти обратно: а б Гольденберг Д.П., Крейтон Т.Э. (апрель 1983 г.). «Круглые и циркулярно переставленные формы ингибитора бычьего трипсина поджелудочной железы». Журнал молекулярной биологии . 165 (2): 407–13. дои : 10.1016/S0022-2836(83)80265-4 . ПМИД 6188846 .
- ^ Перейти обратно: а б с Люгер К., Хоммель У., Херольд М., Хофстинге Дж., Киршнер К. (январь 1989 г.). «Правильное сворачивание циркулярно переставленных вариантов фермента бета-альфа-барреля in vivo». Наука . 243 (4888): 206–10. Бибкод : 1989Sci...243..206L . дои : 10.1126/science.2643160 . ПМИД 2643160 .
- ^ Перейти обратно: а б с д Понтинг КП, Рассел РБ (май 1995 г.). «Свапозины: круговые перестановки внутри генов, кодирующих гомологи сапозина». Тенденции биохимических наук . 20 (5): 179–80. дои : 10.1016/S0968-0004(00)89003-9 . ПМИД 7610480 .
- ^ Ло У.К., Ли СиСи, Ли Сай, Лю ПК. «База данных круговых перестановок» . Институт биоинформатики и структурной биологии Национального университета Цин Хуа . Проверено 16 февраля 2012 г.
- ^ Ло WC, Ли CC, Ли Сай, Лю ПК (январь 2009 г.). «CPDB: база данных круговых перестановок в белках» . Исследования нуклеиновых кислот . 37 (Проблема с базой данных): D328–32. дои : 10.1093/нар/gkn679 . ПМЦ 2686539 . ПМИД 18842637 .
- ^ Каас Кью, Крейк DJ (2010). «Анализ и классификация кольцевых белков в CyBase» . Биополимеры . 94 (5): 584–91. дои : 10.1002/bip.21424 . ПМИД 20564021 .
- ^ Андреева А., Прлич А., Хаббард Т.Дж., Мурзин А.Г. (январь 2007 г.). «СИЗИФ - структурное выравнивание белков с нетривиальными отношениями» . Исследования нуклеиновых кислот . 35 (Проблема с базой данных): D253–9. дои : 10.1093/nar/gkl746 . ПМЦ 1635320 . ПМИД 17068077 .
- ^ Перейти обратно: а б Вайнер Дж., Борнберг-Бауэр Э. (апрель 2006 г.). «Эволюция круговых перестановок в многодоменных белках» . Молекулярная биология и эволюция . 23 (4): 734–43. дои : 10.1093/molbev/msj091 . ПМИД 16431849 .
- ^ Буйницкий Ю.М. (март 2002 г.). «Перестановки последовательностей в молекулярной эволюции ДНК-метилтрансфераз» . Эволюционная биология BMC . 2 (1): 3. дои : 10.1186/1471-2148-2-3 . ПМК 102321 . ПМИД 11914127 .
- ^ Перейти обратно: а б Юнг Дж, Ли Б (сентябрь 2001 г.). «Циркулярно перестановленные белки в базе данных структур белков» . Белковая наука . 10 (9): 1881–6. дои : 10.1110/ps.05801 . ПМК 2253204 . ПМИД 11514678 .
- ^ Джельч А. (июль 1999 г.). «Круговые перестановки в молекулярной эволюции ДНК-метилтрансфераз». Журнал молекулярной эволюции . 49 (1): 161–4. Бибкод : 1999JMolE..49..161J . дои : 10.1007/pl00006529 . PMID 10368444 . S2CID 24116226 .
- ^ Хазкани-Ково Э., Альтман Н., Горовиц М., Граур Д. (январь 2002 г.). «Эволюционная история просапозина: два последовательных события тандемного дублирования привели к появлению четырех доменов сапозина у позвоночных». Журнал молекулярной эволюции . 54 (1): 30–4. Бибкод : 2002JMolE..54...30H . дои : 10.1007/s00239-001-0014-0 . ПМИД 11734895 . S2CID 7402721 .
- ^ Гурупрасад К., Тормякангас К., Кервинен Дж., Бланделл Т.Л. (сентябрь 1994 г.). «Сравнительное моделирование аспарагиновой протеиназы ячменного зерна: структурное обоснование наблюдаемой гидролитической специфичности» . Письма ФЭБС . 352 (2): 131–6. Бибкод : 1994FEBSL.352..131G . дои : 10.1016/0014-5793(94)00935-X . ПМИД 7925961 . S2CID 32524531 .
- ^ Брюн Х (июль 2005 г.). «Краткая экскурсия по функциональным и структурным особенностям сапозиноподобных белков» . Биохимический журнал . 389 (Часть 2): 249–57. дои : 10.1042/BJ20050051 . ПМЦ 1175101 . ПМИД 15992358 .
- ^ Ли Дж., Блабер М. (январь 2011 г.). «Экспериментальная поддержка эволюции симметричной архитектуры белка из простого пептидного мотива» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (1): 126–30. Бибкод : 2011PNAS..108..126L . дои : 10.1073/pnas.1015032108 . ПМК 3017207 . ПМИД 21173271 .
- ^ Перейти обратно: а б Хатефи Ю., Ямагути М. (март 1996 г.). «Никотинамиднуклеотидтрансгидрогеназа: модель использования энергии связывания субстрата для транслокации протонов» . Журнал ФАСЭБ . 10 (4): 444–52. дои : 10.1096/fasebj.10.4.8647343 . ПМИД 8647343 . S2CID 21898930 .
- ^ Торнтон Дж. М., Сибанда Б. Л. (июнь 1983 г.). «Амино- и карбокси-концевые области глобулярных белков». Журнал молекулярной биологии . 167 (2): 443–60. дои : 10.1016/S0022-2836(83)80344-1 . ПМИД 6864804 .
- ^ Ю Ю, Лутц С (январь 2011 г.). «Круговая перестановка: другой способ создания структуры и функции фермента». Тенденции в биотехнологии . 29 (1): 18–25. дои : 10.1016/j.tibtech.2010.10.004 . ПМИД 21087800 .
- ^ Уайтхед Т.А., Бержерон Л.М., Кларк Д.С. (октябрь 2009 г.). «Связывая концы с концами: круговая перестановка снижает протеолитическую восприимчивость рекомбинантных белков» . Белковая инженерия, проектирование и отбор . 22 (10): 607–13. дои : 10.1093/протеин/gzp034 . ПМИД 19622546 .
- ^ Перейти обратно: а б Чельцов А.В., Барбер М.Ю., Феррейра Г.К. (июнь 2001 г.). «Круговая перестановка 5-аминолевулинатсинтазы. Сопоставление полипептидной цепи с ее функцией» . Журнал биологической химии . 276 (22): 19141–9. дои : 10.1074/jbc.M100329200 . ПМЦ 4547487 . ПМИД 11279050 .
- ^ Цянь З, Лутц С (октябрь 2005 г.). «Улучшение каталитической активности липазы B Candida antarctica путем круговой перестановки». Журнал Американского химического общества . 127 (39): 13466–7. дои : 10.1021/ja053932h . ПМИД 16190688 . (основной источник)
- ^ Топелл С., Хеннеке Дж., Глокшубер Р. (август 1999 г.). «Циркулярно перестановленные варианты зеленого флуоресцентного белка». Письма ФЭБС . 457 (2): 283–9. Бибкод : 1999FEBSL.457..283T . дои : 10.1016/S0014-5793(99)01044-3 . ПМИД 10471794 . S2CID 43085373 . (основной источник)
- ^ Вигера А.Р., Серрано Л., Вильманс М. (октябрь 1996 г.). «Различные переходные состояния складывания могут привести к одной и той же собственной структуре». Структурная биология природы . 3 (10): 874–80. дои : 10.1038/nsb1096-874 . ПМИД 8836105 . S2CID 11542397 . (основной источник)
- ^ Капраро Д.Т., Рой М., Онучич Дж.Н., Дженнингс П.А. (сентябрь 2008 г.). «Отступление к складчатому ландшафту белка бета-трилистника интерлейкина-1бета?» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (39): 14844–8. Бибкод : 2008PNAS..10514844C . дои : 10.1073/pnas.0807812105 . ПМК 2567455 . ПМИД 18806223 .
- ^ Чжан П., Шахман Х.К. (июль 1996 г.). «Образование in vivo аллостерической аспартат-транскарбамоилазы, содержащей циркулярно перестановленные каталитические полипептидные цепи: значение для сворачивания и сборки белков» . Белковая наука . 5 (7): 1290–300. дои : 10.1002/pro.5560050708 . ПМК 2143468 . ПМИД 8819162 . (основной источник)
- ^ Хуан Ю.М., Наяк С., Быстрофф С. (ноябрь 2011 г.). «Количественная растворимость in vivo и восстановление усеченных кольцевых пермутантов зеленого флуоресцентного белка» . Белковая наука . 20 (11): 1775–80. дои : 10.1002/pro.735 . ПМЦ 3267941 . ПМИД 21910151 . (основной источник)
- ^ Бирнинк П.Т., Ян Ю.Р., Граф Р., Кинг Д.С., Шах С.С., Шахман Х.К. (март 2001 г.). «Случайная круговая перестановка, приводящая к разрыву цепи внутри и вблизи альфа-спиралей в каталитических цепях аспартат-транскарбамолазы: влияние на сборку, стабильность и функцию» . Белковая наука . 10 (3): 528–37. дои : 10.1110/ps.39001 . ПМЦ 2374132 . ПМИД 11344321 .
- ^ Перейти обратно: а б Бэрд Г.С., Захариас Д.А., Цянь Р.Ю. (сентябрь 1999 г.). «Круговая перестановка и вставка рецепторов в зеленые флуоресцентные белки» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (20): 11241–6. Бибкод : 1999PNAS...9611241B . дои : 10.1073/pnas.96.20.11241 . ЧВК 18018 . ПМИД 10500161 .
- ^ Тернер, Нью-Джерси (август 2009 г.). «Направленная эволюция стимулирует новое поколение биокатализаторов». Химическая биология природы . 5 (8): 567–73. дои : 10.1038/nchembio.203 . ПМИД 19620998 .
- ^ Перейти обратно: а б с д Вайнер Дж., Томас Г., Борнберг-Бауэр Э. (апрель 2005 г.). «Быстрое предсказание круговых перестановок в многодоменных белках на основе мотивов» . Биоинформатика . 21 (7): 932–7. doi : 10.1093/биоинформатика/bti085 . ПМИД 15788783 .
- ^ Перейти обратно: а б Бачар О., Фишер Д., Нусинов Р., Вольфсон Х. (апрель 1993 г.). «Техника, основанная на компьютерном зрении, для трехмерного структурного сравнения белков, независимого от последовательности». Белковая инженерия . 6 (3): 279–88. дои : 10.1093/протеин/6.3.279 . ПМИД 8506262 .
- ^ Перейти обратно: а б с Улиэль С., Флисс А., Амир А., Унгер Р. (ноябрь 1999 г.). «Простой алгоритм обнаружения круговых перестановок в белках» . Биоинформатика . 15 (11): 930–6. дои : 10.1093/биоинформатика/15.11.930 . ПМИД 10743559 .
- ^ Перейти обратно: а б Прлик А., Бливен С., Роуз П.В., Блюм В.Ф., Бизон С., Годзик А., Борн П.Е. (декабрь 2010 г.). «Предварительно рассчитанное выравнивание структуры белка на веб-сайте RCSB PDB» . Биоинформатика . 26 (23): 2983–5. doi : 10.1093/биоинформатика/btq572 . ПМК 3003546 . ПМИД 20937596 .
- ^ Перейти обратно: а б Шацкий М., Нусинов Р., Вольфсон Х.Дж. (июль 2004 г.). «Метод одновременного выравнивания нескольких белковых структур». Белки . 56 (1): 143–56. дои : 10.1002/прот.10628 . ПМИД 15162494 . S2CID 14665486 .
- ^ Цукер М. (сентябрь 1991 г.). «Субоптимальное выравнивание последовательностей в молекулярной биологии. Выравнивание с анализом ошибок». Журнал молекулярной биологии . 221 (2): 403–20. дои : 10.1016/0022-2836(91)80062-Y . ПМИД 1920426 .
- ^ Ло WC, Лю ПК (январь 2008 г.). «CPSARST: эффективный инструмент поиска циклических перестановок, применяемый для обнаружения новых структурных взаимосвязей белков» . Геномная биология . 9 (1): Р11. дои : 10.1186/gb-2008-9-1-r11 . ПМЦ 2395249 . ПМИД 18201387 .
- ^ Шмидт-Геннер Т., Герлер А., Кольбек Б., Кнапп Э.В. (май 2010 г.). «Круговые перестановочные белки во вселенной белковых складок». Белки . 78 (7): 1618–30. дои : 10.1002/прот.22678 . ПМИД 20112421 . S2CID 20673981 .
- ^ Ван Л, Ву ЛИ, Ван Ю, Чжан XS, Чен Л (июль 2010 г.). «SANA: алгоритм последовательного и непоследовательного выравнивания структуры белка». Аминокислоты . 39 (2): 417–25. дои : 10.1007/s00726-009-0457-y . ПМИД 20127263 . S2CID 2292831 .
- ^ Бливен С.Е., Борн П.Е., Прлич А. (апрель 2015 г.). «Обнаружение круговых перестановок внутри белковых структур с использованием CE-CP» . Биоинформатика . 31 (8): 1316–8. doi : 10.1093/биоинформатика/btu823 . ПМЦ 4393524 . ПМИД 25505094 .
- ^ Сиппл М.Ю., Видерштейн М. (апрель 2012 г.). «Обнаружение пространственных корреляций в белковых структурах и молекулярных комплексах» . Структура . 20 (4): 718–28. дои : 10.1016/j.str.2012.01.024 . ПМК 3320710 . ПМИД 22483118 .
Дальнейшее чтение
[ редактировать ]- Дэвид Гудселл (апрель 2010 г.) Конканавалин А и банк данных белков кольцевых перестановок (PDB) Молекула месяца