Хемостат
| Хемостат | |
|---|---|
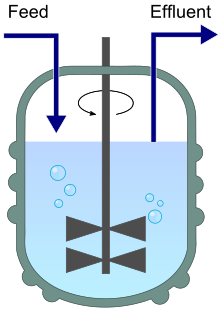 Диаграмма хемостата, показывающая приток (питание) и отток (выход). | |
| Промышленность | Биологическая инженерия |
| Приложение | Исследования и промышленность |

Хемостат в ( химическая среда статична ) представляет собой биореактор, который непрерывно добавляют свежую среду, а культуральную жидкость, содержащую остатки питательных веществ, конечные продукты метаболизма и микроорганизмы, непрерывно удаляют с той же скоростью, чтобы поддерживать постоянный объем культуры. [2] [3] Изменяя скорость добавления среды в биореактор, удельную скорость роста микроорганизма в определенных пределах можно легко контролировать .
Операция
[ редактировать ]Устойчивое состояние
[ редактировать ]Одной из наиболее важных особенностей хемостатов является то, что микроорганизмы можно выращивать в физиологически устойчивом состоянии при постоянных условиях окружающей среды. В этом устойчивом состоянии рост происходит с постоянной удельной скоростью роста , и все параметры культуры остаются постоянными (объем культуры, концентрация растворенного кислорода, концентрации питательных веществ и продуктов, pH, плотность клеток и т. д.). Кроме того, условия окружающей среды могут контролироваться экспериментатором. [4] Микроорганизмы, растущие в хемостатах, обычно достигают устойчивого состояния из-за отрицательной обратной связи между скоростью роста и потреблением питательных веществ: если в биореакторе присутствует небольшое количество клеток, клетки могут расти со скоростью, превышающей скорость разбавления, поскольку они потребляют мало питательных веществ. поэтому рост в меньшей степени ограничивается добавлением ограничивающего питательного вещества с поступающей свежей средой. Лимитирующее питательное вещество – это питательное вещество, необходимое для роста, присутствующее в среде в предельной концентрации (все остальные питательные вещества обычно поставляются в избытке). Однако чем больше становится количество клеток, тем больше питательных веществ потребляется, что снижает концентрацию лимитирующего питательного вещества. В свою очередь, это снизит удельную скорость роста клеток, что приведет к снижению количества клеток, поскольку они продолжают удаляться из системы с оттоком. Это приводит к устойчивому состоянию. Благодаря саморегуляции стационарное состояние является стабильным. Это позволяет экспериментатору контролировать удельную скорость роста микроорганизмов, изменяя скорость насоса, подающего в сосуд свежую среду.
Хорошо смешанный
[ редактировать ]Другой важной особенностью хемостатов и других систем непрерывного культивирования является то, что они хорошо перемешаны, поэтому условия окружающей среды являются однородными или однородными, а микроорганизмы беспорядочно рассредоточены и случайно сталкиваются друг с другом. Следовательно, конкуренция и другие взаимодействия в хемостате носят глобальный характер, в отличие от биопленок .
Скорость разбавления
[ редактировать ]Скорость обмена питательных веществ выражается как степень D. разбавления В устойчивом состоянии удельная скорость роста µ микроорганизма D. равна скорости разбавления Скорость разведения определяется как поток среды F в единицу времени по объему V культуры в биореакторе.

Максимальная скорость роста и критическая скорость разбавления
[ редактировать ]Удельная скорость роста µ обратно пропорциональна времени, необходимому для удвоения биомассы, называемому временем удвоения t d , следующим образом:

Следовательно, время удвоения t d становится функцией степени разбавления D в установившемся состоянии:

Каждый микроорганизм, растущий на определенном субстрате, имеет максимальную удельную скорость роста μ max (скорость роста, наблюдаемая, если рост ограничен внутренними ограничениями, а не внешними питательными веществами). Если выбрана скорость разбавления, превышающая μ max , клетки не могут расти со скоростью, такой же быстрой, как скорость их удаления, поэтому культура не сможет поддерживать себя в биореакторе и вымывается.
Однако, поскольку концентрация ограничивающего питательного вещества в хемостате не может превышать концентрацию в корме, удельная скорость роста, которой могут достичь клетки в хемостате, обычно немного ниже максимальной удельной скорости роста, поскольку удельная скорость роста обычно увеличивается с увеличением питательного вещества. концентрация, описываемая кинетикой уравнения Моно . [ нужна ссылка ] Наивысшая удельная скорость роста ( μmax ) , которую могут достичь клетки, равна критической скорости разведения ( D' c ):

где S — концентрация субстрата или питательного вещества в хемостате, а KS — константа полунасыщения (это уравнение предполагает кинетику Моно).
Приложения
[ редактировать ]Исследовать
[ редактировать ]Хемостаты в исследованиях используются для исследований в области клеточной биологии в качестве источника больших объемов однородных клеток или белка. Хемостат часто используется для сбора данных об устойчивом состоянии организма с целью создания математической модели, относящейся к его метаболическим процессам. Хемостаты также используются в качестве микрокосмов в экологии. [5] [6] и эволюционная биология. [7] [8] [9] [10] В одном случае мутация/отбор — это помеха, в другом — желаемый изучаемый процесс. Хемостаты также можно использовать для обогащения культур определенных типов бактериальных мутантов, таких как ауксотрофы или устойчивые к антибиотикам или бактериофагам , для дальнейшего научного изучения. [11] Вариации скорости разведения позволяют изучать метаболические стратегии, реализуемые организмами при разных скоростях роста. [12] [13]
Конкуренция за одиночные и множественные ресурсы, эволюция путей приобретения и использования ресурсов, перекрестное питание/симбиоз, [14] [15] антагонизм, хищничество и конкуренция среди хищников изучались в экологии и эволюционной биологии с использованием хемостатов. [16] [17] [18]
Промышленность
[ редактировать ]Хемостаты часто используются при промышленном производстве этанола . В этом случае последовательно используются несколько хемостатов, каждый из которых поддерживает понижение концентрации сахара. [ нужна ссылка ] Хемостат также служит экспериментальной моделью непрерывных культур клеток в биотехнологической промышленности. [13]
Технические проблемы
[ редактировать ]- Вспенивание приводит к переливу, при этом объем жидкости не совсем постоянный.
- Некоторые очень хрупкие клетки разрываются во время перемешивания и аэрации .
- Клетки могут расти на стенках или прилипать к другим поверхностям. [19] Эту проблему можно преодолеть, обработав стеклянные стенки сосуда силаном, чтобы сделать их гидрофобными. Однако ячейки будут выбраны для прикрепления к стенам, поскольку те, которые это сделают, не будут удалены из системы. Те бактерии, которые прочно прикрепляются к стенкам, образуя биопленку , трудно изучать в условиях хемостата.
- Смешивание может быть не совсем равномерным, что нарушает «статические» свойства хемостата.
- Капание среды в камеру фактически приводит к небольшим импульсам питательных веществ и, следовательно, к колебаниям концентраций, что снова нарушает «статические» свойства хемостата.
- Бактерии довольно легко перемещаются вверх по течению. Они быстро достигнут резервуара со стерильной средой, если путь жидкости не будет прерван воздушным разрывом, при котором среда падает каплями через воздух.
Постоянные усилия по устранению каждого дефекта довольно регулярно приводят к изменениям в базовом хемостате. Примеров в литературе множество.
- Для подавления пенообразования используются пеногасители.
- Перемешивание и аэрацию можно проводить осторожно.
- Было предпринято множество подходов для уменьшения роста стенок. [20] [21]
- В различных приложениях для смешивания используются лопасти, барботаж и другие механизмы. [22]
- Капание можно сделать менее резким за счет меньших капель и большего объема сосуда.
- Многие улучшения направлены на устранение угрозы загрязнения.
Соображения экспериментального дизайна
[ редактировать ]Этот раздел в значительной степени или полностью опирается на один источник . ( декабрь 2019 г. ) |
Выбор и настройка параметров
[ редактировать ]- Стационарная концентрация лимитирующего субстрата в хемостате не зависит от концентрации притока. Концентрация притока будет влиять на концентрацию клеток и, следовательно, на OD в устойчивом состоянии.
- Несмотря на то, что предельная концентрация субстрата в хемостате обычно очень низка и поддерживается дискретными высококонцентрированными импульсами притока, на практике временные изменения концентрации внутри хемостата невелики (несколько процентов или меньше) и, таким образом, могут рассматриваться как квазистационарное состояние.
- Время, необходимое для того, чтобы плотность клеток (ОП) приблизилась к установившемуся значению (превышение/недостаток), часто бывает длительным (несколько оборотов хемостата), особенно когда исходный инокулят большой. Но время можно минимизировать при правильном выборе параметров.
Стабильный рост
[ редактировать ]- Может показаться, что хемостат находится в устойчивом состоянии, но захваты мутантных штаммов могут происходить непрерывно, даже если они не обнаруживаются путем мониторинга параметров макромасштаба, таких как OD или концентрации продукта.
- Ограничивающий субстрат обычно находится в таких низких концентрациях, что его невозможно обнаружить. В результате концентрация лимитирующего субстрата может сильно меняться со временем (в процентном отношении), поскольку разные штаммы захватывают популяцию, даже если результирующие изменения OD слишком малы, чтобы их можно было обнаружить.
- «Импульсный» хемостат (с очень большими импульсами притока) имеет существенно меньшую селективную способность, чем стандартный квазинепрерывный хемостат, для мутантного штамма с повышенной приспособленностью в лимитирующих условиях.
- Резко снижая концентрацию субстрата, ограничивающую приток, можно временно подвергнуть клетки относительно более жестким условиям до тех пор, пока хемостат не стабилизируется обратно в устойчивое состояние (во временном порядке степени разбавления D).
Мутация
[ редактировать ]- Некоторые типы мутантных штаммов появятся быстро:
- Если существует SNP , который может повысить приспособленность, он должен появиться в популяции всего лишь после нескольких удвоений хемостата для характерно больших хемостатов (например, 10^11 клеток E. coli ).
- Штамм, которому необходимы два конкретных SNP, где только их комбинация дает преимущество в приспособленности (тогда как каждый из них по отдельности нейтрален), вероятно, появится только в том случае, если целевой размер (количество различных местоположений SNP, которые приводят к выгодной мутации) для каждого SNP очень большой.
- Появление других типов мутантных штаммов (например, двух SNP с небольшим размером мишени, большего количества SNP или хемостатов меньшего размера) маловероятно.
- Эти другие мутации ожидаются только в результате последовательных выборок мутантов с преимуществом в приспособленности. Можно ожидать возникновения множественных мутантов только в том случае, если каждая мутация полезна независимо, а не в тех случаях, когда мутации нейтральны по отдельности, но вместе выгодны. Последовательные поглощения — единственный надежный способ продолжения эволюции в хемостате.
- Казалось бы, экстремальный сценарий, когда нам требуется, чтобы каждый возможный SNP хотя бы один раз сосуществовал в хемостате, на самом деле вполне вероятен. Большой хемостат с большой вероятностью достигнет этого состояния.
- Для большого хемостата ожидаемое время до появления выгодной мутации порядка времени оборота хемостата. Обратите внимание, что обычно это значительно короче, чем время, необходимое для захвата популяции хемостата выгодным штаммом. Это не обязательно так в небольшом хемостате.
- Ожидается, что вышеуказанные моменты будут одинаковыми для разных видов бесполого размножения ( E. coli , S. cerevisiae и т. д.).
- Более того, время до появления мутации не зависит от размера генома, но зависит от частоты мутаций каждого BP.
- Для характерно больших хемостатов гипермутирующий штамм не дает достаточного преимущества, чтобы оправдать его использование. Кроме того, у него недостаточно селективного преимущества, чтобы можно было ожидать, что он всегда появится в результате случайной мутации и возьмет на себя управление хемостатом.
Единое поглощение
[ редактировать ]- Время перехода предсказуемо с учетом соответствующих параметров деформации.
- Различные степени разведения избирательно способствуют захвату популяции хемостата разными мутантными штаммами, если такой штамм существует. Например:
- Высокая скорость разведения создает давление отбора для мутантного штамма с повышенной максимальной скоростью роста;
- Средняя степень разведения создает давление отбора для мутантного штамма с более высоким сродством к лимитирующему субстрату;
- Медленная скорость разведения создает давление отбора для мутантного штамма, который может расти в среде без ограничивающего субстрата (предположительно за счет потребления другого субстрата, присутствующего в среде);
- Время захвата превосходящего мутанта будет довольно постоянным в диапазоне рабочих параметров. Для характеристических рабочих значений время срабатывания составляет от нескольких дней до недель.
Последовательные поглощения
[ редактировать ]- При подходящих условиях (достаточно большая популяция и множество мишеней в геноме для простых полезных мутаций) ожидается, что несколько штаммов последовательно захватят популяцию, и сделают это относительно синхронно и в темпе. Сроки зависят от типа мутации.
- В последовательности поглощений, даже если избирательное улучшение каждого из штаммов остается постоянным (например, каждый новый штамм лучше предыдущего в постоянный коэффициент) – скорость поглощения не остается постоянной, а скорее уменьшается от штамма к штамму.
- Бывают случаи, когда последовательные поглощения происходят настолько быстро, что отличить штаммы очень трудно даже при изучении частоты аллелей. Таким образом, линия множественных поглощений последовательных штаммов может выглядеть как поглощение одного штамма с группой мутаций.
Вариации
[ редактировать ]Установки для ферментации, тесно связанные с хемостатами, - это турбидостат , ауксостат и ретентостат . В ретентостатах культуральная жидкость также удаляется из биореактора, но биомасса удерживается фильтром. В этом случае концентрация биомассы увеличивается до тех пор, пока потребность в питательных веществах для поддержания биомассы не станет равна количеству лимитирующего питательного вещества, которое может быть потреблено.
См. также
[ редактировать ]- Бактериальный рост
- Биохимическая инженерия
- Реактор непрерывного действия с мешалкой (CSTR)
- Эксперимент по долгосрочной эволюции E. coli
- Фед-пакет
Ссылки
[ редактировать ]- ^ Мэдиган, Майкл (2015). Брок Биология микроорганизмов . Пирсон. стр. 152–153. ISBN 978-0-321-89739-8 .
- ^ Новик А., Сцилард Л. (1950). «Описание хемостата». Наука . 112 (2920): 715–6. Бибкод : 1950Sci...112..715N . дои : 10.1126/science.112.2920.715 . ПМИД 14787503 .
- ^ Джеймс Т.В. (1961). «Непрерывная культура микроорганизмов». Ежегодный обзор микробиологии . 15 : 27–46. дои : 10.1146/annurev.mi.15.100161.000331 .
- ^ Д Герберт; Р. Элсворт; Р. К. Теллинг (1956). «Непрерывная культура бактерий: теоретическое и экспериментальное исследование» . Дж. Генерал Микробиол . 14 (3): 601–622. дои : 10.1099/00221287-14-3-601 . ПМИД 13346021 .
- ^ Бекс Л., Хилкер Ф.М., Мальхов Х., Юргенс К., Арндт Х. (2005). «Экспериментальная демонстрация хаоса в микробной пищевой сети». Природа . 435 (7046): 1226–9. Бибкод : 2005Natur.435.1226B . дои : 10.1038/nature03627 . ПМИД 15988524 . S2CID 4380653 .
- ^ Павлу С., Кеврекидис И.Г. (1992). «Микробное хищничество в периодически работающем хемостате: глобальное исследование взаимодействия между естественными и внешними частотами». Математические бионауки . 108 (1): 1–55. дои : 10.1016/0025-5564(92)90002-E . ПМИД 1550993 .
- ^ Вичман Х.А., Миллштейн Дж., Булл Дж.Дж. (2005). «Адаптивная молекулярная эволюция для 13 000 поколений фагов: возможная гонка вооружений» . Генетика . 170 (1): 19–31. дои : 10.1534/genetics.104.034488 . ПМК 1449705 . ПМИД 15687276 .
- ^ Дыхейзен Д.Э., Дин А.М. (2004). «Эволюция специалистов в экспериментальном микромире» . Генетика . 167 (4): 2015–26. doi : 10.1534/genetics.103.025205 . ПМК 1470984 . ПМИД 15342537 .
- ^ Вик Л.М., Вайленманн Х., Эгли Т. (2002). «Очевидная часовая эволюция Escherichia coli в хемостатах с ограниченным содержанием глюкозы воспроизводится при больших, но не при малых размерах популяции, и может быть объяснена кинетикой Моно» . Микробиология . 148 (Часть 9): 2889–902. дои : 10.1099/00221287-148-9-2889 . ПМИД 12213934 .
- ^ Джонс Л.Е., Эллнер С.П. (2007). «Влияние быстрой эволюции добычи на циклы хищник-жертва». J Математика Биол . 55 (4): 541–73. arXiv : q-bio/0609032 . дои : 10.1007/s00285-007-0094-6 . ПМИД 17483952 . S2CID 16927689 .
- ^ Шлегель Х.Г., Яннаш Х.В. (1967). «Обогащающие культуры». Анну. Преподобный Микробиол . 21 : 49–70. дои : 10.1146/annurev.mi.21.100167.000405 . ПМИД 4860267 .
- ^ Варма, А.; Палссон, Б.О. (1 октября 1994 г.). «Модели баланса стехиометрических потоков количественно предсказывают рост и секрецию побочных продуктов метаболизма у Escherichia coli W3110 дикого типа» . Прикладная и экологическая микробиология . 60 (10): 3724–3731. Бибкод : 1994ApEnM..60.3724V . дои : 10.1128/aem.60.10.3724-3731.1994 . ISSN 0099-2240 . ЧВК 201879 . ПМИД 7986045 .
- ^ Jump up to: а б Фернандес-де-Коссио-Диас, Хорхе; Леон, Калет; Муле, Роберто (13 ноября 2017 г.). «Характеристика устойчивого состояния метаболических сетей масштаба генома в непрерывных клеточных культурах» . PLOS Вычислительная биология . 13 (11): e1005835. arXiv : 1705.09708 . Бибкод : 2017PLSCB..13E5835F . дои : 10.1371/journal.pcbi.1005835 . ISSN 1553-7358 . ПМК 5703580 . ПМИД 29131817 .
- ^ Дотон К.Г., Се Д.П. (1977). «Утилизация паратиона бактериальными симбионтами в хемостате» . Прил. Окружающая среда. Микробиол . 34 (2): 175–84. Бибкод : 1977ApEnM..34..175D . doi : 10.1128/aem.34.2.175-184.1977 . ПМК 242618 . ПМИД 410368 .
- ^ Пфайффер Т., Бонхёффер С. (2004). «Эволюция перекрестного питания в микробных популяциях». Являюсь. Нат . 163 (6): E126–35. дои : 10.1086/383593 . ПМИД 15266392 . S2CID 31110741 .
- ^ Дж. Дж. Батлер; ГСК Волкович (июль 1986 г.). «Конкуренция, опосредованная хищником, в хемостате». J Математика Биол . 24 (2): 67–191. дои : 10.1007/BF00275997 . S2CID 120858390 .
- ^ Дыхуйзен Д.Э., Хартл Д.Л. (июнь 1983 г.). «Отбор в хемостатах» . Микробиол. Преподобный . 47 (2): 150–68. дои : 10.1128/г.47.2.150-168.1983 . ПМК 281569 . ПМИД 6308409 .
- ^ Дыхейзен Д.Э., Хартл Д.Л. (май 1981 г.). «Эволюция конкурентоспособности Escherichia coli». Эволюция . 35 (3): 581–94. дои : 10.2307/2408204 . JSTOR 2408204 . ПМИД 28563589 .
- ^ Бономи А., Фредриксон АГ (1976). «Питание простейших и рост бактериальных стенок». Биотехнология. Биоинж. 18 (2): 239–52. дои : 10.1002/бит.260180209 . ПМИД 1267931 . S2CID 41343643 .
- ^ де Креси Э., Мецгар Д., Аллен С., Пенико М., Лайонс Б., Хансен С.Дж., де Креси-Лагард В. (2007). «Разработка нового устройства непрерывного культивирования для экспериментальной эволюции бактериальных популяций». Прил. Микробиол. Биотехнология. 77 (2): 489–96. дои : 10.1007/s00253-007-1168-5 . ПМИД 17896105 . S2CID 25787277 .
- ^ Чжан З., Боккацци П., Чой Х.Г., Пероцциелло Дж., Сински А.Дж., Дженсен К.Ф. (2006). «Микрохемостат - непрерывная культура микробов в инструментальном микробиореакторе на полимерной основе». Лабораторный чип . 6 (7): 906–13. дои : 10.1039/b518396k . ПМИД 16804595 .
- ^ Ван Хюлле С.В., Ван Ден Брук С., Мартенс Дж., Виллес К., Шелстрете Г., Волке Э.И., Ванроллегем П.А. (2003). «Практический опыт запуска и эксплуатации лабораторного реактора ШАРОН с непрерывной аэрацией». Коммун. Сельское хозяйство. Прил. Биол. наук. 68 (2 часть А): 77–84. ПМИД 15296140 .
- ^ Jump up to: а б с д и Уайдс А., Майло Р. (2018). «Понимание динамики и оптимизация эффективности экспериментов по выбору хемостата». arXiv : 1806.00272 [ q-bio.PE ].
Внешние ссылки
[ редактировать ]- http://www.pererikstrandberg.se/examensarbete/chemostat.pdf
- https://web.archive.org/web/20060504172359/http://www.rpi.edu/dept/chem-eng/Biotech-Environ/Contin/chemosta.htm
- Заключительная диссертация, включающая математические модели хемостата и других биореакторов.
- Страница об одной конструкции лабораторного хемостата.
- Подробное руководство по хемостату (лаборатория Данэма). Процедуры и принципы являются общими.