Инвертированный повтор
Инвертированный повтор (или IR ) представляет собой одноцепочечную последовательность нуклеотидов , за которой следует обратный комплемент . [ 1 ] Промежуточная последовательность нуклеотидов между исходной последовательностью и обратным комплементом может иметь любую длину, включая нулевую. Например, 5'---TTACGnnnnnn CGTAA---3' представляет собой инвертированную повторяющуюся последовательность. Когда промежуточная длина равна нулю, составная последовательность является палиндромной последовательностью . [ 2 ]
Как инвертированные повторы, так и прямые повторы представляют собой типы нуклеотидных последовательностей , которые встречаются повторно. Эти повторяющиеся последовательности ДНК часто варьируются от пары нуклеотидов до целого гена , в то время как близость повторяющихся последовательностей варьируется между широко рассредоточенными и простыми тандемными массивами . [ 3 ] Короткие тандемные повторяющиеся последовательности могут существовать в количестве от нескольких копий в небольшом участке до тысяч копий, рассеянных по всему геному большинства эукариот . [ 4 ] Повторяющиеся последовательности, содержащие около 10–100 пар оснований , известны как минисателлиты , а более короткие повторяющиеся последовательности, содержащие в основном 2–4 пары оснований, известны как микросателлиты . [ 5 ] К наиболее распространенным повторам относятся динуклеотидные повторы, имеющие основания AC на одной цепи ДНК и GT на комплементарной цепи. [ 3 ] Некоторые элементы генома с уникальными последовательностями функционируют как экзоны , интроны и регуляторная ДНК. [ 6 ] Хотя наиболее знакомыми локусами повторяющихся последовательностей являются центромера и теломера , [ 6 ] большая часть повторяющихся последовательностей в геноме находится среди некодирующей ДНК . [ 5 ]
Инвертированные повторы выполняют ряд важных биологических функций. Они определяют границы транспозонов и указывают области, способные к самокомплементарному спариванию оснований (области внутри одной последовательности, которые могут спариваться друг с другом). Эти свойства играют важную роль в нестабильности генома. [ 7 ] и способствовать не только клеточной эволюции и генетическому разнообразию [ 8 ] но также к мутациям и болезням . [ 9 ] Для детального изучения этих эффектов был разработан ряд программ и баз данных, помогающих обнаруживать и аннотировать инвертированные повторы в различных геномах.
Понимание инвертированных повторов
[ редактировать ]Пример инвертированного повтора
[ редактировать ]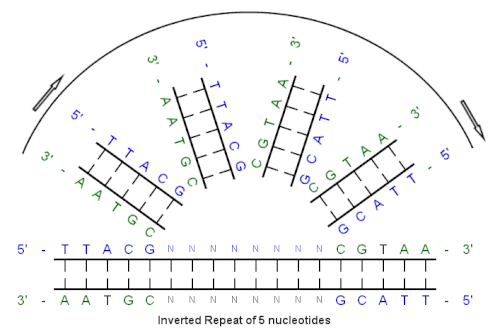
Начиная с этой начальной последовательности:
5'-TTACG-3'
Дополнение, созданное спариванием оснований:
3'-AATGC-5'
Обратное дополнение:
5'-ЦГТАА-3'
И инвертированная повторяющаяся последовательность:
5'---TTACGnnnnnn ВГТАА---3'
«nnnnnn» представляет собой любое количество промежуточных нуклеотидов.
Против. прямой повтор
[ редактировать ]Прямое повторение происходит, когда последовательность повторяется с тем же шаблоном в дальнейшем. [ 1 ] Не существует инверсии и обратного дополнения, связанного с прямым повтором. Нуклеотидная последовательность, написанная жирным шрифтом, означает повторяющуюся последовательность. Он может иметь или не иметь промежуточные нуклеотиды.
- 5´ TTACG nnnnnnTTACG 3´
- 3´ AATGC nnnnnnAATGC 5´
С лингвистической точки зрения типичный прямой повтор сравним с рифмой, например, «время на время » .
Против. тандемный повтор
[ редактировать ]Прямой повтор без промежуточных нуклеотидов между исходной последовательностью и ее последующей копией представляет собой тандемный повтор . Нуклеотидная последовательность, написанная жирным шрифтом, означает повторяющуюся последовательность.
- 5´ ТТАЧГ ТТАЧГ 3´
- 3 ' ААТГК ААТГК 5'
С лингвистической точки зрения типичное тандемное повторение сравнимо с заиканием или намеренным повторением слов, например «пока-пока».
Против. палиндром
[ редактировать ]Инвертированная повторная последовательность без промежуточных нуклеотидов между исходной последовательностью и ее последующим обратным комплементом представляет собой палиндром . [ 1 ]
ПРИМЕР:
Шаг 1: начните с перевернутого повтора: 5' ТТАКГнннннКГТАА 3'
Шаг 2: удалите промежуточные нуклеотиды: 5' ТТАГКГТАА 3'
Полученная последовательность является палиндромной, поскольку она является обратным дополнением самой себе. [ 1 ]
- 5'-TTACGCGTAA-3'- тестовая последовательность (из этапа 2 с удалением промежуточных нуклеотидов)
- 3'-AATGCGCATT 5'- дополнение тестовой последовательности
- 5' TTACGCGTAA 3' обратный комплемент. Это то же самое, что и тестовая последовательность выше, и, следовательно, это палиндром.
Биологические особенности и функциональность
[ редактировать ]Условия, благоприятствующие синтезу
[ редактировать ]Разнообразные полногеномные повторы происходят из мобильных элементов , которые, как сейчас понимают, «перепрыгивают» в разные места генома, не перенося своих исходных копий. [ 10 ] Последующее перемещение одних и тех же последовательностей по многочисленным поколениям обеспечивает их множественность по всему геному. [ 10 ] Ограниченная рекомбинация последовательностей между двумя отдельными элементами последовательности, известная как консервативная сайт-специфическая рекомбинация (CSSR), приводит к инверсиям сегмента ДНК, основанным на расположении последовательностей распознавания рекомбинации на ДНК донора и ДНК реципиента. [ 10 ] Опять же, ориентация двух сайтов рекомбинации внутри донорной молекулы ДНК относительно асимметрии промежуточных последовательностей расщепления ДНК, известная как область кроссовера, имеет решающее значение для образования либо инвертированных повторов, либо прямых повторов. [ 10 ] Таким образом, рекомбинация, происходящая в паре инвертированных сайтов, инвертирует последовательность ДНК между двумя сайтами. [ 10 ] Наблюдались очень стабильные хромосомы со сравнительно меньшим количеством инвертированных повторов, чем с прямыми повторами, что позволяет предположить наличие связи между стабильностью хромосом и количеством повторов. [ 11 ]
Регионы, где присутствие обязательно
[ редактировать ]Концевые инвертированные повторы наблюдались в ДНК различных эукариотических транспозонов, хотя их источник остается неизвестным. [ 12 ] Инвертированные повторы в основном обнаруживаются в истоках репликации клеточного организма и органелл, от фаговых плазмид, митохондрий и эукариотических вирусов до клеток млекопитающих. [ 13 ] Истоки репликации фага G4 и других родственных фагов включают сегмент из почти 139 нуклеотидных оснований, который включает три инвертированных повтора, которые необходимы для прайминга репликации. [ 13 ]
В геноме
[ редактировать ]В значительной степени участки нуклеотидных повторов нередко наблюдаются в составе редких комбинаций ДНК. [ 14 ] Три основных повтора, которые в основном встречаются в конкретных конструкциях ДНК, включают очень точные гомопурин-гомопиримидиновые инвертированные повторы, которые иначе называются H-палиндромами, что является обычным явлением в тройных спиральных H-конформациях, которые могут включать либо триады нуклеотидов TAT, либо CGC. Остальные можно описать как длинные инвертированные повторы, имеющие тенденцию к образованию шпилек и крестообразных повторов, и, наконец, как прямые тандемные повторы, которые обычно существуют в структурах, описываемых как скользящая петля, крестообразная и левосторонняя Z-ДНК. [ 14 ]
Общие у разных организмов
[ редактировать ]Прошлые исследования показывают, что повторы являются общей чертой эукариот, в отличие от прокариот и архей . [ 14 ] Другие сообщения позволяют предположить, что, несмотря на сравнительную нехватку повторяющихся элементов в геномах прокариот, они, тем не менее, содержат сотни и даже тысячи крупных повторов. [ 15 ] Текущий геномный анализ, по-видимому, предполагает существование большого избытка идеальных инвертированных повторов во многих геномах прокариот по сравнению с геномами эукариот. [ 16 ]

Количественную оценку и сравнение инвертированных повторов у нескольких видов, а именно у архей, см. [ 17 ]
Инвертированные повторы в псевдоузлах
[ редактировать ]Псевдоузлы — это распространенные структурные мотивы, встречающиеся в РНК. Они образованы двумя вложенными друг в друга петлями стебля, так что стержень одной структуры образуется из петли другой. Среди псевдоузлов существует множество складчатых топологий и большие различия в длине петель, что делает их структурно разнообразной группой. [ 18 ]
Инвертированные повторы являются ключевым компонентом псевдоузлов, как можно видеть на иллюстрации встречающегося в природе псевдоузла, обнаруженного в компоненте теломеразной РНК человека . [ 19 ] В эту структуру вовлечены четыре разных набора инвертированных повторов. Наборы 1 и 2 являются основой стебля-петли A и являются частью петли для стебля-петли B. Аналогично, наборы 3 и 4 являются основой для стебля-петли B и частью петли для стебля-петли A.
Псевдоузлы играют множество различных ролей в биологии. Псевдоузел теломеразы на иллюстрации имеет решающее значение для активности этого фермента. [ 19 ] Рибозим сворачивается вируса гепатита дельта (HDV) в структуру с двойным псевдоузлом и саморасщепляет свой кольцевой геном с образованием РНК длиной в один геном. Псевдоузлы также играют роль в запрограммированном сдвиге рамки рибосом, обнаруженном у некоторых вирусов и необходимом при репликации ретровирусов . [ 18 ]
В рибопереключателях
[ редактировать ]Инвертированные повторы играют важную роль в рибопереключателях , которые являются регуляторными элементами РНК, контролирующими экспрессию генов, продуцирующих мРНК, частью которой они являются. [ 10 ] Упрощенный пример рибопереключателя флавинмононуклеотида (FMN) показан на иллюстрации. Этот рибопереключатель существует в транскрипте мРНК и имеет несколько структур «стебель-петля» выше кодирующей области . Однако на иллюстрации показаны только ключевые стебли-петли, которые были значительно упрощены, чтобы показать роль перевернутых повторов. В этом рибопереключателе имеется несколько инвертированных повторов, которые обозначены зеленым (желтый фон) и синим (оранжевый фон).
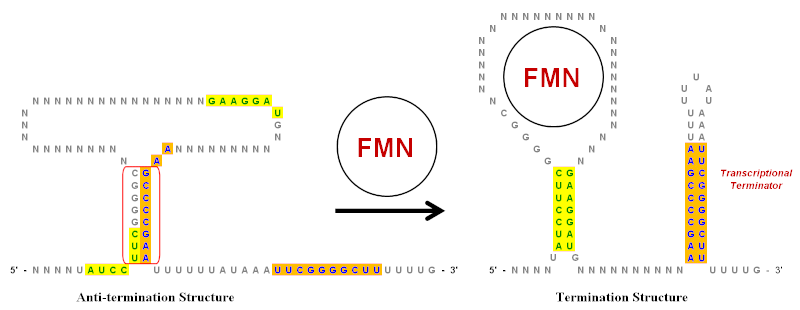
В отсутствие FMN структура антитерминации является предпочтительной конформацией транскрипта мРНК. Он создается путем спаривания оснований области инвертированного повтора, обведенной красным. Когда присутствует FMN, он может связываться с петлей и предотвращать образование структуры антитерминации. Это позволяет двум различным наборам инвертированных повторов образовывать пару оснований и формировать структуру завершения. [ 20 ] Стебель-петля на 3'-конце является терминатором транскрипции, поскольку последовательность, следующая сразу за ней, представляет собой цепочку урацилов (U). Если эта стебель-петля образуется (из-за присутствия FMN) при выходе растущей цепи РНК из комплекса РНК-полимеразы , это создаст достаточное структурное напряжение, чтобы заставить цепь РНК диссоциировать и, таким образом, прекратить транскрипцию. Диссоциация происходит легко, поскольку пары оснований между буквами U в РНК и буквами А в цепи матрицы являются самыми слабыми из всех пар оснований. [ 10 ] Таким образом, при более высоких уровнях концентрации FMN подавляет собственную транскрипцию за счет увеличения образования терминационной структуры.
Мутации и болезни
[ редактировать ]Инвертированные повторы часто называют «горячими точками» эукариотической и прокариотической геномной нестабильности. [ 7 ] Считается, что длинные инвертированные повторы сильно влияют на стабильность генома различных организмов. [ 21 ] Это проиллюстрировано на примере E. coli , где геномные последовательности с длинными инвертированными повторами редко реплицируются, а скорее быстро удаляются. [ 21 ] Опять же, длинные инвертированные повторы, наблюдаемые у дрожжей, в значительной степени способствуют рекомбинации внутри одной и той же и соседних хромосом, что приводит к столь же очень высокой частоте делеций. [ 21 ] Наконец, очень высокая скорость делеции и рекомбинации наблюдалась также в участках хромосом млекопитающих с инвертированными повторами. [ 21 ] Сообщаемые различия в стабильности геномов взаимосвязанных организмов всегда являются показателем несоответствия инвертированных повторов. [ 11 ] Нестабильность возникает из-за тенденции инвертированных повторов сворачиваться в структуры ДНК, похожие на шпильки или крестообразные. Эти специальные структуры могут препятствовать или сбивать с толку репликацию ДНК и другие виды геномной деятельности. [ 7 ] Таким образом, инвертированные повторы приводят к особым конфигурациям как РНК , так и ДНК , которые в конечном итоге могут вызывать мутации и заболевания . [ 9 ]

На иллюстрации показан инвертированный повтор, подвергшийся крестообразной экструзии. ДНК в области инвертированного повтора раскручивается, а затем рекомбинируется, образуя четырехстороннее соединение с двумя «стебель-петля» структурами . Крестообразная структура возникает из-за того, что инвертированные повторяющиеся последовательности самоспариваются друг с другом на своей собственной цепи. [ 22 ]
Выдавленные крестообразные формы могут привести к мутациям сдвига рамки считывания , когда последовательность ДНК имеет инвертированные повторы в форме палиндрома в сочетании с областями прямых повторов с обеих сторон. Во время транскрипции проскальзывание и частичная диссоциация полимеразы от цепи матрицы могут привести как к делеционным, так и к инсерционным мутациям. [ 9 ] Делеция происходит, когда часть раскрученной цепи матрицы образует стебель-петлю, которая «пропускается» механизмом транскрипции. Вставка происходит, когда в диссоциированной части возникающей (вновь синтезированной) цепи образуется стебель-петля, в результате чего часть цепи матрицы транскрибируется дважды. [ 9 ]
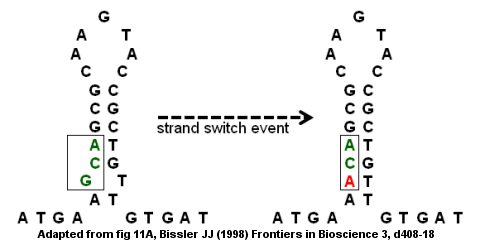
Дефицит антитромбина вследствие точечной мутации
[ редактировать ]Несовершенные инвертированные повторы могут приводить к мутациям посредством внутрицепочечных и межцепочечных переключений. [ 9 ] Кодирующая область гена антитромбина III является примером несовершенного инвертированного повтора , как показано на рисунке справа. Структура стебля -петли образуется с выступом внизу, потому что буквы G и T не соединяются в пары. Событие переключения цепи может привести к замене буквы G (на выступе) на букву A, что устраняет «несовершенство» в инвертированном повторе и обеспечивает более прочную структуру стебель-петля. Однако замена также создает точечную мутацию, превращающую кодон GCA в ACA. Если за событием переключения цепи последует второй раунд репликации ДНК , мутация может закрепиться в геноме и привести к заболеванию. В частности, миссенс-мутация приведет к дефектному гену и дефициту антитромбина, что может привести к развитию венозной тромбоэмболии (сгустков крови в венах). [ 9 ]

Несовершенный остеогенез вследствие мутации сдвига рамки считывания
[ редактировать ]Мутации в гене коллагена могут привести к заболеванию несовершенного остеогенеза , которое характеризуется ломкостью костей. [ 9 ] На иллюстрации ствол-петля, образованная из несовершенного инвертированного повтора, мутирована с вставкой нуклеотида тимина (Т) в результате меж- или внутрицепочечного переключения. Добавление буквы Т создает «совпадение» пары оснований с аденином (А), который ранее был «выпуклостью» на левой стороне стебля. Хотя это добавление делает стебель более сильным и совершенствует инвертированный повтор, оно также создает мутацию сдвига рамки считывания в нуклеотидной последовательности, которая изменяет рамку считывания и приводит к неправильной экспрессии гена. [ 9 ]
Программы и базы данных
[ редактировать ]В следующем списке представлена информация и внешние ссылки на различные программы и базы данных для инвертированных повторов:
- Non-B DB База данных для интегрированных аннотаций и анализа мотивов, образующих ДНК, отличных от B. [ 23 ] Эта база данных предоставлена Центром передовых биомедицинских вычислений (ABCC) тогдашней Национальной лаборатории исследования рака имени Фредерика (FNLCR). Он охватывает конформации A-ДНК и Z-ДНК, также известные как «ДНК, отличные от B», поскольку они не являются более распространенной формой B-ДНК правосторонней двойной спирали Уотсона-Крика . Эти «не-B ДНК» включают левостороннюю Z-ДНК, крестообразную , триплексную , тетраплексную и шпильчную структуры. [ 23 ] Поиск может осуществляться по множеству «типов повторов» (включая инвертированные повторы) и по нескольким видам.
- База данных инвертированных повторов Бостонского университета. Эта база данных представляет собой веб-приложение, которое позволяет запрашивать и анализировать повторы, содержащиеся в проекте PUBLIC DATABASE. Ученые также могут анализировать свои собственные последовательности с помощью алгоритма поиска инвертированных повторов. [ 24 ]
- P-MITE: база данных растительных MITE — эта база данных миниатюрных мобильных элементов с инвертированными повторами (MITE) содержит последовательности из геномов растений . Последовательности можно искать или загружать из базы данных. [ 25 ]
- EMBOSS — это «Европейский пакет открытого программного обеспечения для молекулярной биологии», который работает на UNIX и UNIX-подобных операционных системах. [ 26 ] Документация и исходные файлы программы доступны на сайте EMBOSS . Приложения, специально связанные с инвертированными повторами, перечислены ниже:
- EMBOSS einverted : Находит инвертированные повторы в нуклеотидных последовательностях . Пороговые значения могут быть установлены для ограничения области поиска. [ 26 ]
- Палиндром EMBOSS : находит палиндромы, такие как области стеблевой петли в нуклеотидных последовательностях. Программа найдет последовательности, которые включают в себя участки несовпадений и пробелов, которые могут соответствовать выпуклостям в петле стебля. [ 26 ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д Ассери, Дэвид В.; Вассенаар, Труди; Борини, Стефано (22 декабря 2008 г.). «Частоты слов, повторы и структуры, связанные с повторами, в бактериальных геномах». Вычисления для сравнительной микробной геномики: биоинформатика для микробиологов . Вычислительная биология. Том. 8 (1-е изд.). Спрингер. стр. 133–144. ISBN 978-1-84800-254-8 .
- ^ Йе, Конгтинг; Цзи, Гуоли; Лян, Чун (2014). «detectIR: новая программа для обнаружения идеальных и несовершенных инвертированных повторов с использованием комплексных чисел и векторных вычислений» . ПЛОС ОДИН . 9 (11): e113349. Бибкод : 2014PLoSO...9k3349Y . дои : 10.1371/journal.pone.0113349 . ПМЦ 4237412 . ПМИД 25409465 .
- ^ Перейти обратно: а б Ричардс, Греция; Ричардс, Род-Айленд (25 апреля 1995 г.). «Простые тандемные повторы ДНК и генетические заболевания человека» . Труды Национальной академии наук Соединенных Штатов Америки . 92 (9): 3636–41. Бибкод : 1995PNAS...92.3636S . дои : 10.1073/pnas.92.9.3636 . ПМК 42017 . ПМИД 7731957 .
- ^ ван Белкум, А; Шерер, С; ван Альфен, Л; Вербру, Х. (июнь 1998 г.). «Короткие последовательности ДНК-повторов в геномах прокариот» . Обзоры микробиологии и молекулярной биологии . 62 (2): 275–93. дои : 10.1128/MMBR.62.2.275-293.1998 . ПМЦ 98915 . ПМИД 9618442 .
- ^ Перейти обратно: а б Рамель, К. (июнь 1997 г.). «Мини- и микроспутники» . Перспективы гигиены окружающей среды . 105 (Приложение 4): 781–9. дои : 10.2307/3433284 . JSTOR 3433284 . ПМК 1470042 . ПМИД 9255562 .
- ^ Перейти обратно: а б Эйхлер, Э.Э. (август 1998 г.). «Маскарад повторяется: паралогические ловушки человеческого генома» . Геномные исследования . 8 (8): 758–62. дои : 10.1101/гр.8.8.758 . ПМИД 9724321 .
- ^ Перейти обратно: а б с Миркин, И; Нараянан, В; Лобачев К.С.; Миркин С.М. (22 июля 2008 г.). «Остановка репликации при нестабильных инвертированных повторах: взаимодействие между шпильками ДНК и белками, стабилизирующими вилку» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (29): 9936–41. Бибкод : 2008PNAS..105.9936V . дои : 10.1073/pnas.0804510105 . ПМЦ 2481305 . ПМИД 18632578 .
- ^ Лин, Коннектикут; Лин, Вашингтон; Лю, ЮЛ; Ванг-Пэн, Дж. (1 сентября 2001 г.). «Инвертированные повторы как генетические элементы, способствующие инвертированной дупликации ДНК: значение амплификации генов» . Исследования нуклеиновых кислот . 29 (17): 3529–38. дои : 10.1093/нар/29.17.3529 . ПМК 55881 . ПМИД 11522822 .
- ^ Перейти обратно: а б с д и ж г час Бисслер, Джей-Джей (27 марта 1998 г.). «Инвертированные повторы ДНК и болезни человека» (PDF) . Границы бионауки . 3 (4): д408–18. дои : 10.2741/a284 . ПМИД 9516381 . S2CID 12982 . Архивировано из оригинала (PDF) 3 марта 2019 г.
- ^ Перейти обратно: а б с д и ж г Школа, Джеймс Д. Уотсон, Лаборатория Колд-Спринг-Харбор, Таня А. Бейкер, Массачусетский технологический институт, Стивен П. Белл, Массачусетский технологический институт, Александр Ганн, Лаборатория Колд-Спринг-Харбор, Майкл Левин, Калифорнийский университет, Беркли, Ричард Лосик, Гарвардский университет; со Стивеном К. Харрисоном, Гарвардский медицинский университет (2014). Молекулярная биология гена (Седьмое изд.). Бостон: Издательская компания Бенджамина-Каммингса. ISBN 9780321762436 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Перейти обратно: а б Ачаз, Г; Куассак, Э; Неттер, П; Роча, EP (август 2003 г.). «Связь между инвертированными повторами и структурной эволюцией бактериальных геномов» . Генетика . 164 (4): 1279–89. дои : 10.1093/генетика/164.4.1279 . ПМЦ 1462642 . ПМИД 12930739 .
- ^ Чжан, Х.Х.; Сюй, ХЭ; Шен, Ю.Х.; Хан, MJ; Чжан, Z (январь 2013 г.). «Происхождение и эволюция шести миниатюрных перевернуто-повторяющихся мобильных элементов у Bombyx mori и Rhodnius prolixus» . Геномная биология и эволюция . 5 (11): 2020–31. дои : 10.1093/gbe/evt153 . ПМЦ 3845634 . ПМИД 24115603 .
- ^ Перейти обратно: а б Пирсон, CE; Зорбас, Х; Цена, ГБ; Заннис-Хаджопулос, М (октябрь 1996 г.). «Инвертированные повторы, стеблевые петли и крестообразные формы: значение для инициации репликации ДНК». Журнал клеточной биохимии . 63 (1): 1–22. doi : 10.1002/(SICI)1097-4644(199610)63:1<1::AID-JCB1>3.0.CO;2-3 . ПМИД 8891900 . S2CID 22204780 .
- ^ Перейти обратно: а б с Херинга, Дж. (июнь 1998 г.). «Обнаружение внутренних повторов: насколько они распространены?». Современное мнение в области структурной биологии . 8 (3): 338–45. дои : 10.1016/S0959-440X(98)80068-7 . ПМИД 9666330 .
- ^ Треанген, Ти Джей; Авраам, Алабама; Тушон, М; Роча, EP (май 2009 г.). «Генезис, эффекты и судьба повторов в геномах прокариот» (PDF) . Обзоры микробиологии FEMS . 33 (3): 539–71. дои : 10.1111/j.1574-6976.2009.00169.x . ПМИД 19396957 .
- ^ Ладукакис, Эд; Эйр-Уокер, А. (сентябрь 2008 г.). «Избыток малых инвертированных повторов у прокариот» (PDF) . Журнал молекулярной эволюции . 67 (3): 291–300. Бибкод : 2008JMolE..67..291L . CiteSeerX 10.1.1.578.7466 . дои : 10.1007/s00239-008-9151-z . ПМИД 18696026 . S2CID 29953202 .
- ^ Хоссейни, М; Пратас, Д; Пиньо, Эй Джей (2017). «О роли инвертированных повторов в сходстве последовательностей ДНК». 11-я Международная конференция по практическому применению вычислительной биологии и биоинформатики . Достижения в области интеллектуальных систем и вычислений. Том. 616. Спрингер. стр. 228–236. дои : 10.1007/978-3-319-60816-7_28 . ISBN 978-3-319-60815-0 .
- ^ Перейти обратно: а б Степлер, Д.В.; Мясник, SE (июнь 2005 г.). «Псевдоузлы: структуры РНК с разнообразными функциями» . ПЛОС Биология . 3 (6): е213. дои : 10.1371/journal.pbio.0030213 . ПМЦ 1149493 . ПМИД 15941360 .

- ^ Перейти обратно: а б Чен, Дж.Л.; Грейдер, CW (7 июня 2005 г.). «Функциональный анализ структуры псевдоузла теломеразной РНК человека» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (23): 8080–5, обсуждение 8077–9. Бибкод : 2005PNAS..102.8080C . дои : 10.1073/pnas.0502259102 . ПМЦ 1149427 . ПМИД 15849264 .
- ^ Винклер, WC; Коэн-Чаламиш, С; Брейкер, Р.Р. (10 декабря 2002 г.). «Структура мРНК, которая контролирует экспрессию генов путем связывания FMN» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (25): 15908–13. Бибкод : 2002PNAS...9915908W . дои : 10.1073/pnas.212628899 . ПМК 138538 . ПМИД 12456892 .
- ^ Перейти обратно: а б с д Стормо, Дж; Чанг, Кентукки; Варлей, К; Стормо, Джорджия (28 февраля 2007 г.). Холл, Нил (ред.). «Доказательства активного поддержания инвертированных повторяющихся структур, выявленных с помощью сравнительного геномного подхода» . ПЛОС ОДИН . 2 (2): е262. Бибкод : 2007PLoSO...2..262Z . дои : 10.1371/journal.pone.0000262 . ПМК 1803023 . ПМИД 17327921 .
- ^ Рамредди, Т; Сачиданандам, Р; Стрик, Т.Р. (май 2011 г.). «Обнаружение крестообразной экструзии в режиме реального времени с помощью наноманипуляции одиночной молекулы ДНК» . Исследования нуклеиновых кислот . 39 (10): 4275–83. дои : 10.1093/nar/gkr008 . ПМК 3105387 . ПМИД 21266478 .
- ^ Перейти обратно: а б Цер, РЗ; Донохью, Делавэр; Мудунури, США; Темиз, Н.А.; Лосс, Массачусетс; Старнер, Нью-Джерси; Халуса, Г.Н.; Вольфовский, Н; Йи, М; Люк, БТ; Баколла, А; Коллинз-младший; Стивенс, Р.М. (январь 2013 г.). «БД данных Non-B v2.0: база данных предсказанных мотивов, образующих ДНК, отличных от B, и связанных с ней инструментов» . Исследования нуклеиновых кислот . 41 (Проблема с базой данных): D94–D100. дои : 10.1093/nar/gks955 . ПМЦ 3531222 . ПМИД 23125372 .
- ^ Гельфанд Ю.; Родригес, А; Бенсон, Дж. (январь 2007 г.). «TRDB — база данных тандемных повторов» . Исследования нуклеиновых кислот . 35 (Проблема с базой данных): D80–7. дои : 10.1093/нар/gkl1013 . ПМЦ 1781109 . ПМИД 17175540 .
- ^ Чен, Дж; Ху, Кью; Чжан, Ю; Лу, С; Куанг, Х. (29 октября 2013 г.). «P-MITE: база данных миниатюрных растительных мобильных элементов с инвертированным повторением» . Исследования нуклеиновых кислот . 42 (1): Д1176–81. дои : 10.1093/нар/gkt1000 . ПМЦ 3964958 . ПМИД 24174541 .
- ^ Перейти обратно: а б с Райс, П; Лонгден, я; Блисби, А. (июнь 2000 г.). «EMBOSS: Европейский пакет открытого программного обеспечения для молекулярной биологии» . Тенденции в генетике . 16 (6): 276–7. дои : 10.1016/S0168-9525(00)02024-2 . ПМИД 10827456 .
Внешние ссылки
[ редактировать ]- Инвертированный + повтор + последовательность Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)