Рибосомальная ДНК
Рибосомальная ДНК (сокращенно рДНК ) состоит из группы рибосомальных РНК, кодирующих гены и родственные регуляторные элементы, и широко распространена в сходной конфигурации во всех сферах жизни. Рибосомальная ДНК кодирует некодирующую рибосомальную РНК , неотъемлемые структурные элементы в сборке рибосом , ее важность делает ее наиболее распространенным участком РНК, обнаруживаемым в клетках эукариот . [ 1 ] Кроме того, эти сегменты включают регуляторные участки, такие как промотор, специфичный для РНК-полимеразы I , а также как транскрибируемые, так и нетранскрибируемые спейсерные сегменты.
Из-за их высокой важности в сборке рибосом для биосинтеза белка гены рДНК обычно высоко консервативны в молекулярной эволюции . Количество копий может значительно варьироваться в зависимости от вида. [ 1 ] Рибосомальная ДНК широко используется в филогенетических исследованиях. [ 2 ] [ 3 ]
Структура
[ редактировать ]| Тип | СГУ рРНК | LSU рРНК |
|---|---|---|
| Эукариоты | 18S рРНК | 28S рРНК 5.8S рРНК 5S рРНК |
| Бактерии | 16S рРНК | 23S рРНК 5S рРНК |
| Митохондриальный | MT-RNR1 (12S рРНК) | MT-RNR2 (16S рРНК) |
| Пластид | 16S рРНК | 23S рРНК 4.5S рРНК 5S рРНК |
Рибосомальная ДНК включает все гены, кодирующие некодирующие структурные молекулы рибосомальной РНК . Во всех сферах жизни это структурные последовательности малой субъединицы ( 16S или 18S рРНК ) и большой субъединицы ( 23S или 28S рРНК ). Сборка последнего также включает 5S рРНК , а также дополнительную 5,8S рРНК у эукариот.
Гены рДНК обычно присутствуют в геноме в нескольких копиях, где у большинства видов они организованы в связанные группы, разделенные внутренним транскрибируемым спейсером (ITS) и которому предшествует внешний транскрибируемый спейсер (ETS) . также 5S рРНК связана с этим участком рДНК у прокариот она расположена в отдельных повторяющихся участках , тогда как у большинства эукариот . [ 4 ] Они вместе транскрибируются в РНК-предшественник, которая затем обрабатывается до равных количеств каждой рРНК.
Прокариоты
[ редактировать ]Первичные структурные молекулы рРНК у бактерий и архей меньше, чем их аналоги у эукариот, и сгруппированы как 16S рРНК и 23S рРНК . Между тем, 5S рРНК, также присутствующая у прокариот, имеет такой же размер, как и у эукариот.
Заметное количество бактерий и архей отклоняются от канонической структуры оперона, содержащего гены рДНК, неся «несвязанные» гены в разных местах своего генома. [ 5 ]
Пластиды
[ редактировать ]Рибосомальная ДНК в хлоропластах повторяет структуру рибосомальной ДНК прокариот.
Эукариоты
[ редактировать ]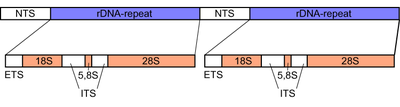

Кластер генов рДНК эукариот состоит из генов 18S , 5.8S и 28S рРНК , разделенных двумя спейсерами ITS-1 и ITS-2 . Активный геном эукариот содержит несколько сотен копий транскрипционной единицы рДНК в виде тандемных повторов , они организованы в области ядрышковых организаторов (ЯОР) . [ 4 ] которые, в свою очередь, могут присутствовать в нескольких локусах генома. [ 6 ]
Подобно структуре прокариот, 5S рРНК присоединяется к кластеру рДНК у сахаромицетов (гемиаскомицетов). [ 6 ] такие как Saccharomyces cerevisiae . [ 4 ] Однако большинство эукариот несут ген 5S рРНК в отдельных повторах гена в разных локусах генома. [ 6 ] [ 4 ]
5S рДНК также присутствует в независимых тандемных повторах, как у дрозофилы . [ 6 ] Поскольку повторяющиеся участки ДНК часто подвергаются событиям рекомбинации, повторы рДНК имеют множество регуляторных механизмов, которые удерживают ДНК от мутаций. [ нужен пример ] таким образом сохраняя консервацию рДНК. [ 1 ]
В ядре районы-организаторы ядрышка дают начало ядрышку , где участки рДНК хромосомы образуют расширенные хромосомные петли, доступные для транскрипции рРНК . В рДНК тандемные повторы преимущественно обнаруживаются в ядрышке; но гетерохроматическая рДНК находится вне ядрышка. Однако транскрипционно активная рДНК находится внутри самого ядрышка. [ 1 ]
Люди
[ редактировать ]Геном человека содержит в общей сложности 560 копий. [ 4 ] транскрипционной единицы рДНК, распределенной по пяти хромосомам с областями ядрышкового организатора . Кластеры повторов расположены на акроцентрических хромосомах 13 ( RNR1 ), 14 ( RNR2 ), 15 ( RNR3 ), 21 ( RNR4 ) и 22 ( RNR5 ). [ 7 ]
Инфузории
[ редактировать ]У инфузорий наличие генеративного микроядра рядом с вегетативным макронуклеусом позволяет редуцировать гены рДНК в зародышевой линии. Точное количество копий в ядре генома микроядра варьируется от нескольких копий у Paramecium. [ 8 ] всего один экземпляр у Tetrahymena thermophila. [ 4 ] и другие виды Tetrahymena . Во время формирования макронуклеуса участки, содержащие кластеры генов рДНК, амплифицируются, резко увеличивая количество доступных матриц для транскрипции до нескольких тысяч копий. У некоторых родов инфузорий, например Tetrahymena или Hypotrich рода Oxytricha , [ 8 ] обширная фрагментация амплифицированной ДНК приводит к образованию микрохромосом, сосредоточенных на транскрипционной единице рДНК. [ 8 ] Подобные процессы наблюдаются у Glaucomachattoni и в меньшей степени у Paramecium . [ 8 ]
Однородность последовательности
[ редактировать ]В большом массиве рДНК полиморфизм между повторяющимися единицами рДНК очень низок, что указывает на то, что тандемные массивы рДНК развиваются в результате согласованной эволюции . [ 6 ] Однако механизм согласованной эволюции несовершенен, так что полиморфизмы между повторами внутри особи могут возникать на значительных уровнях и могут затруднить филогенетический анализ близкородственных организмов. [ 9 ] [ 10 ]
Последовательности 5S-тандемных повторов у нескольких дрозофил сравнили друг с другом; результат показал, что вставки и делеции часто происходили между видами и часто сопровождались консервативными последовательностями. [ 11 ] Они могут произойти в результате проскальзывания вновь синтезированной цепи во время репликации ДНК или в результате конверсии гена. [ 11 ]
Расхождение последовательностей
[ редактировать ]Транскрипционные пути рДНК имеют низкий уровень полиморфизма среди видов, что позволяет проводить межвидовые сравнения для выяснения филогенетических взаимоотношений, используя лишь несколько образцов. Кодирующие области рДНК высококонсервативны среди видов, но области ITS изменчивы из-за инсерций, делеций и точковых мутаций. Сравнение последовательностей в участках ITS между отдаленными видами, такими как человек и лягушка, нецелесообразно. [ 12 ] Консервативные последовательности кодирующих областей рДНК позволяют сравнивать отдаленные виды, даже между дрожжами и человеком. 5.8S рРНК человека на 75% идентична дрожжевой 5.8S рРНК. [ 13 ] В случае видов-двойников сравнение сегмента рДНК, включая участки ITS, среди видов и филогенетический анализ проведены удовлетворительно. [ 14 ] [ 15 ] Различные кодирующие области повторов рДНК обычно демонстрируют разную скорость эволюции. В результате эта ДНК может предоставлять филогенетическую информацию о видах, принадлежащих к широким систематическим уровням. [ 2 ]
Рекомбинационно-стимулирующая активность
[ редактировать ]Фрагмент дрожжевой рДНК, содержащий ген 5S, нетранскрибируемую спейсерную ДНК и часть гена 35S, обладает локализованной цис-действующей митотическую рекомбинацию . активностью, стимулирующей [ 16 ] Этот фрагмент ДНК содержит горячую точку митотической рекомбинации , называемую HOT1. HOT1 проявляет активность, стимулирующую рекомбинацию, когда он вставляется в новые места генома дрожжей . HOT1 включает транскрипции РНК-полимеразы I (PolI) промотор , который катализирует транскрипцию гена 35S рибосомальной рРНК . У мутанта с дефектом PolI активность, стимулирующая рекомбинацию горячих точек HOT1, отменена. Уровень транскрипции PolI в HOT1, по-видимому, определяет уровень рекомбинации . [ 17 ]
Клиническое значение
[ редактировать ]Заболевания могут быть связаны с мутациями ДНК, при которых ДНК может расширяться, например болезнь Хантингтона , или утрачиваться из-за делеционных мутаций. То же самое относится и к мутациям, возникающим в повторах рДНК; Было обнаружено, что если гены, связанные с синтезом рибосом, нарушаются или мутируют, это может привести к различным заболеваниям, связанным со скелетом или костным мозгом. Кроме того, любое повреждение или нарушение работы ферментов, защищающих тандемные повторы рДНК, может привести к снижению синтеза рибосом, что также приводит к другим дефектам в клетке. Неврологические заболевания также могут возникать из-за мутаций в тандемных повторах рДНК, например, синдром Блума , который возникает, когда количество тандемных повторов увеличивается почти в сто раз; по сравнению с нормальным количеством тандемных повторов. Различные типы рака также могут возникать в результате мутаций тандемных повторов рибосомальной ДНК. Клеточные линии могут стать злокачественными либо в результате перестройки тандемных повторов, либо в результате расширения повторов в рДНК. [ 18 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д Вармердам, Даниэль О.; Волтуис, Роб МФ (01 марта 2019 г.). «Сохранение рибосомальной ДНК нетронутой: повторяющаяся задача» . Хромосомные исследования . 27 (1): 57–72. дои : 10.1007/s10577-018-9594-z . ISSN 1573-6849 . ПМК 6394564 . ПМИД 30556094 .
- ^ Jump up to: а б Хиллис DM, Диксон MT (декабрь 1991 г.). «Рибосомальная ДНК: молекулярная эволюция и филогенетический вывод». Ежеквартальный обзор биологии . 66 (4): 411–53. дои : 10.1086/417338 . ПМИД 1784710 . S2CID 32027097 .
- ^ Вайсбург В.Г., Барнс С.М., Пеллетье Д.А., Лейн DJ (январь 1991 г.). «Амплификация рибосомальной ДНК 16S для филогенетических исследований» . Журнал бактериологии . 173 (2): 697–703. дои : 10.1128/jb.173.2.697-703.1991 . ПМК 207061 . ПМИД 1987160 .
- ^ Jump up to: а б с д и ж Грау, Йохен (2015). Genetik [ Генетика ] (на немецком языке) (6-е изд.). Берлин, Гейдельберг: Springer-Verlag Berlin Heidelberg. дои : 10.1007/978-3-662-44817-5 . ISBN 978-3-662-44816-8 .
- ^ Брюэр Т.Э., Альбертсен М., Эдвардс А., Киркегор Р.Х., Роча Э.П., Фирер Н. (февраль 2020 г.). «Несвязанные гены рРНК широко распространены среди бактерий и архей» . Журнал ISME . 14 (2): 597–608. Бибкод : 2020IMAGE..14..597B . дои : 10.1038/ s41396-019-0552-3 ПМК 6976660 . ПМИД 31712737 .
- ^ Jump up to: а б с д и Ричард Г.Ф., Керрест А., Дужон Б. (декабрь 2008 г.). «Сравнительная геномика и молекулярная динамика повторов ДНК у эукариот» . Обзоры микробиологии и молекулярной биологии . 72 (4): 686–727. дои : 10.1128/MMBR.00011-08 . ПМЦ 2593564 . ПМИД 19052325 .
- ^ Шмидт, Олаф (2017). Фриче, Олаф (ред.). и Генетика молекулярная биология Компактные знания биологии (на немецком языке). Берлин, Гейдельберг: Springer-Verlag Berlin Heidelberg. дои : 10.1007/978-3-662-50274-7 . ISBN 978-3-662-50273-0 .
- ^ Jump up to: а б с д Прескотт, Дэвид М. (июнь 1994 г.). «ДНК реснитчатых простейших» . Микробиологические обзоры . 58 (2): 233–267. дои : 10.1128/мр.58.2.233-267.1994 . ПМК 372963 . ПМИД 8078435 .
- ^ Альварес I, Вендел Дж. Ф. (декабрь 2003 г.). «Рибосомальные последовательности ITS и филогенетический вывод растений». Молекулярная филогенетика и эволюция . 29 (3): 417–34. Бибкод : 2003МОЛПЭ..29..417А . дои : 10.1016/S1055-7903(03)00208-2 . ПМИД 14615184 .
- ^ Вайтемир К., Штрауб С.К., Фишбейн М., Листон А. (2015). «Внутригеномные полиморфизмы среди высококопийных локусов: общеродовое исследование ядерной рибосомальной ДНК у Asclepias (Apocynaceae)» . ПерДж . 3 : е718. дои : 10.7717/peerj.718 . ПМК 4304868 . ПМИД 25653903 .
- ^ Jump up to: а б Пакес Ф., Самсон М.Л., Джордан П., Вегнес М. (ноябрь 1995 г.). «Структурная эволюция рибосомальных генов 5S дрозофилы». Журнал молекулярной эволюции . 41 (5): 615–21. Бибкод : 1995JMolE..41..615P . дои : 10.1007/bf00175820 . ПМИД 7490776 . S2CID 6911205 .
- ^ Сумида М., Като Ю., Курабаяши А. (апрель 2004 г.). «Секвенирование и анализ внутренних транскрибируемых спейсеров (ITS) и кодирующих областей во фрагменте EcoRI I рибосомальной ДНК японской прудовой лягушки Rana nigromaculata» . Гены и генетические системы . 79 (2): 105–18. дои : 10.1266/ggs.79.105 . ПМИД 15215676 .
- ^ Назар Р.Н., Ситц Т.О., Буш Х. (февраль 1976 г.). «Гомологии последовательностей 5.8S рибосомальной РНК млекопитающих». Биохимия . 15 (3): 505–8. дои : 10.1021/bi00648a008 . ПМИД 1252408 .
- ^ Фэнъи М.Ю., Цзяньнун X, Чжэмин Z (1998). «Различия последовательностей рДНК-ITS2 и видоводиагностического ПЦР-анализа Anopheles sinensis и Anopheles anthropophagus из Китая» (PDF) . J Med Coll PLA . 13 : 123–128.
- ^ Ли, С; Ли, Дж. С.; Гребнер, Дж.Л.; Ким, ХК; Кляйн, Т.А.; О'Гуинн, ML; Вилкерсон, Р.К. (2005). «Недавно признанный вид группы Anopheles hyrcanus и молекулярная идентификация родственных видов из Южной Кореи (Diptera: Culicidae)» (PDF) . Зоотакса . 939 : 1–8. дои : 10.11646/zootaxa.939.1.1 . Архивировано из оригинала 1 октября 2012 года.
- ^ Кейл Р.Л., Редер Г.С. (декабрь 1984 г.). «Цис-действующая, стимулирующая рекомбинацию активность фрагмента рибосомальной ДНК S. cerevisiae» . Клетка . 39 (2, часть 1): 377–86. дои : 10.1016/0092-8674(84)90016-3 . ПМИД 6094015 .
- ^ Серидзава Н., Хориучи Т., Кобаяши Т. (апрель 2004 г.). «Опосредованная транскрипцией гиперрекомбинация в HOT1». Гены в клетки . 9 (4): 305–15. дои : 10.1111/j.1356-9597.2004.00729.x . ПМИД 15066122 . S2CID 23978914 .
- ^ Вармердам, DO, Wolthuis RM (март 2019 г.). «Сохранение рибосомальной ДНК нетронутой: повторяющаяся задача» . Хромосомные исследования . 27 (1–2): 57–72. дои : 10.1007/s10577-018-9594-z . ПМК 6394564 . ПМИД 30556094 .