Подковообразная летучая мышь
| Подковообразные биты | |
|---|---|

| |
| Малая подковоносная летучая мышь ( Rhinolophus hipposideros ) с синей металлической идентификационной полосой на левом крыле. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Рукокрылые |
| Семья: | Ринолофиды Грей , 1825 г. |
| Подсемейство: | Ринолофины Грей , 1825 г. |
| Род: | Ринолоф Ласепед , 1799 г. |
| Типовой вид | |
| Железная бита Шребер , 1774 г.
| |
| Разновидность | |
Подковоносы — летучие мыши семейства Rhinolophidae . Помимо единственного живого рода , Rhinolophus насчитывающего около 106 видов, вымерший род Palaeonycteris признан . Подковоносы тесно связаны с листоносыми летучими мышами Старого Света семейства Hipposideridae, которых иногда включали в состав Rhinolophidae. Подковоносы делятся на шесть подродов и множество видовых групп. Самый последний общий предок всех подковоносов жил 34–40 миллионов лет назад, хотя неясно, где находятся географические корни этого семейства, и попытки определить его биогеографию оказались нерешительными. Их таксономия сложна, поскольку генетические данные показывают вероятное существование многих загадочных видов , а также видов, признанных отдельными, которые могут иметь небольшое генетическое отличие от ранее признанных таксонов. Они встречаются в Старом Свете , в основном в тропических и субтропических регионах, включая Африку, Азию, Европу и Океанию.
Подковоносы считаются летучими мышами малого или среднего размера , весом 4–28 г (0,14–0,99 унции), с длиной предплечий 30–75 мм (1,2–3,0 дюйма) и общей длиной головы и тела 35–110 мм ( 1,4–4,3 дюйма). Мех, длинный и гладкий у большинства видов, может быть красновато-коричневым, черноватым или ярко-оранжево-красным. Свое общее название они получили из-за больших носовых листьев , имеющих форму подковы. Носовые листки помогают в эхолокации ; Подковоносы обладают очень сложной эхолокацией: они используют сигналы постоянной частоты в интенсивных циклах для обнаружения добычи в районах с высоким уровнем помех в окружающей среде. Они охотятся на насекомых и пауков, налетая на добычу с насеста или собирая ее с листвы. Об их системах спаривания мало что известно, но по крайней мере один вид моногамен, а другой — полигинен . Беременность длится примерно семь недель, и за один раз рождается один потомок. Типичная продолжительность жизни составляет шесть или семь лет, но одна большая подковоносная летучая мышь прожила более тридцати лет.
Подковоносы актуальны для человека в некоторых регионах как источник болезней, как пища и для традиционной медицины . Несколько видов являются естественными резервуарами различных коронавирусов, связанных с атипичной пневмонией , и данные убедительно свидетельствуют о том, что они являются резервуаром SARS-CoV , хотя люди могут столкнуться с большим риском заражения со стороны промежуточных хозяев, таких как маскированные пальмовые циветты . [ 1 ]
На них охотятся ради еды в нескольких регионах, особенно в Африке к югу от Сахары , а также в Юго-Восточной Азии . Некоторые виды или их гуано используются в традиционной медицине Непала, Индии, Вьетнама и Сенегала.
Таксономия и эволюция
[ редактировать ]Таксономическая история
[ редактировать ]Rhinolophus был впервые описан как род в 1799 году французским натуралистом Бернаром Жерменом де Ласепедом . Первоначально все дошедшие до нас подковоносы относились к Rhinolophus , а также к нынешнему виду Hipposideros (круглолистные летучие мыши). [ 2 ] : хii Первоначально Rhinolophus принадлежал к семейству Vespertilionidae . В 1825 году британский зоолог Джон Эдвард Грей разделил Vespertilionidae на подсемейства, включая так называемых Rhinolophina. [ 3 ] Английский зоолог Томас Белл считается первым, кто выделил подковоносов в отдельное семейство, используя Rhinolophidae в 1836 году. [ 4 ] Хотя Белла иногда признают авторитетом в области ринолофид, [ 5 ] авторитет чаще указывается как Грей, 1825 г. [ 4 ] [ 6 ] Подковоносы относятся к надсемейству Rhinolophoidea , наряду с Craseonycteridae , Hipposideridae Megadermatidae , Rhinonycteridae и Rhinopomatidae . [ 7 ] [ 8 ]
Были предприняты попытки разделить Rhinolophus на другие роды. В 1816 году английский зоолог Уильям Элфорд Лич предложил название рода Phyllorhina ; Грей предложил Аквиаса в 1847 году и Филлотиду в 1866 году; и немецкий натуралист Вильгельм Петерс предложил Coelophyllus в 1867 году. В 1876 году ирландский зоолог Джордж Эдвард Добсон вернул всех азиатских подковоносов к Rhinolophus , дополнительно предложив подсемейства Phyllorhininae (для гиппосидерид) и Rhinolophinae. Американский зоолог Геррит Смит Миллер-младший в 1907 году отделил гиппозидеридов от подковоносов, признав Hipposideridae отдельным семейством. [ 2 ] : хii Некоторые авторы считали Hipposideros и связанные с ним роды частью Rhinolophidae еще в начале 2000-х годов. [ 9 ] хотя сейчас их чаще всего признают отдельной семьей. [ 10 ] [ 11 ] После разделения на Rhinolophidae и Hipposideridae для Rhinolophus были предложены дальнейшие подразделения : Rhinolphyllotis в 1934 году и Rhinomegalophus в 1951 году, хотя оба дополнительных рода были возвращены Rhinolophus . [ 2 ] : хii
Датский маммолог Кнуд Андерсен был первым, кто предложил группы видов для Rhinolophus , сделав это в 1905 году. Группы видов — это способ группировки видов для отражения эволюционных взаимоотношений. Он выделил шесть групп видов: R. simplex (ныне R. megaphyllus ), R. lepidus , R. Midas (ныне R. hipposideros ), R. philippinensis , R. macrotis и R. arcuatus . Виды часто перераспределялись между группами по мере добавления новых групп, описания новых видов и пересмотра взаимоотношений между видами. [ 2 ] : xiii Пятнадцать видовых групп были выделены Чорбой и его коллегами в 2003 году. [ 2 ] [ 12 ] различные подроды Также были предложены , шесть из которых перечислены Csorba et al . в 2003 году: Aquias , Phyllorhina , Rhinolophus , Indorhinolophus , Coelophyllus и Rhinophyllotis . [ 2 ] : xvi Неофициально ринолофидов можно разделить на две основные клады : преимущественно африканскую кладу и преимущественно восточную кладу. [ 9 ]
Эволюционная история
[ редактировать ]
Самый последний общий предок ринолофов жил примерно 34–40 миллионов лет назад. [ 13 ] отделение от линии гиппосидерид в эоцене . [ 9 ] Ископаемые подковоносы известны из Европы (ранний-средний миоцен , ранний олигоцен ), Австралии (миоцен) и Африки (миоцен и поздний плиоцен ). [ 14 ] Биогеография подковоносов изучена слабо. Различные исследования предполагают, что семья возникла в Европе, Азии или Африке. Исследование 2010 года подтвердило азиатское или восточное происхождение семейства с быстрым эволюционным распространением африканских и восточных клад в олигоцене. [ 9 ] Исследование 2019 года показало, что R. xinanzhongguoensis и R. nippon , оба евразийских вида, более тесно связаны с африканскими видами, чем с другими евразийскими видами, что позволяет предположить, что ринолофиды могут иметь сложные биогеографические связи с Азией и афротропиками . [ 13 ]
Исследование 2016 года с использованием митохондриальной и ядерной ДНК поместило подковоносов в состав Yinpterochiroptera как сестер Hipposideridae. [ 8 ]
| Рукокрылые |
| ||||||||||||||||||





Ринолофиды представлены одним современным родом — Rhinolophus . И семейство, и род подтверждены как монофилетические (содержащие всех потомков общего предка). было описано 106 видов По состоянию на 2019 год у Rhinolophus , что делает его вторым по разнообразию родом летучих мышей после Myotis . Выборка ринолофов может существовать до двенадцати загадочных видов в афротропической зоне может быть недостаточной: по оценкам одного генетического исследования, в этом регионе . Кроме того, было обнаружено, что некоторые таксоны, признанные полноценными видами, имеют небольшую генетическую дивергенцию . Rhinolophus kahuzi может быть синонимом подковоноса Рувензори ( R. ruwenzorii ), а R. gorongosae или R. rhodesiae могут быть синонимами подковоноса Бушвельда ( R. Simulator ). Кроме того, подковоносы Смитерса ( R. smithersi ), подковоносы Коэна ( R. cohenae ) и подковоносы Маунт-Мабу ( R. mabuensis ) имеют небольшое генетическое отличие от подковоносов Хильдебрандта ( R. hildebrandtii ). Признание первых трех полноценными видами оставляет подковообразную летучую мышь Хильдебрандта. парафилетический . [ 13 ]
Второй род Rhinolophidae — вымерший Palaeonycteris с типовым видом Palaeonycterisrobustus . [ 15 ] Palaeonycterisrobustus жил в нижнем миоцене, и его окаменелые останки были найдены в Сен-Жеран-ле-Пюи , Франция. [ 16 ] [ 17 ]
Описание
[ редактировать ]Появление
[ редактировать ]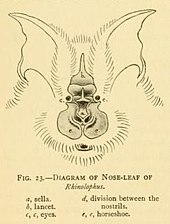
Подковообразные летучие мыши считаются маленькими или средними летучими мышами . [ 10 ] Особи имеют длину головы и тела в пределах 35–110 мм (1,4–4,3 дюйма), а длину предплечий 30–75 мм (1,2–3,0 дюйма). Один из более мелких видов, малый подковонос ( R. hipposideros ), весит 4–10 г (0,14–0,35 унции), а один из более крупных видов, большой подковонос ( R. Ferrumequinum ), весит 16,5–28 г. (0,58–0,99 унции). Цвет меха у разных видов сильно различается: от черноватого до красновато-коричневого и до ярко-оранжево-красного. [ 18 ] [ 14 ] Нижняя часть тела светлее шерсти на спине. [ 18 ] У большинства видов длинный и мягкий мех, но шерстистые и малошерстистые подковоносы ( R. luctus и R. beddomei ) необычны своим очень длинным шерстистым мехом. [ 14 ]
Как и у большинства летучих мышей, у подковоносов есть две молочные железы на груди . У взрослых самок на животе дополнительно есть два выступа, похожих на соски, называемые лобковыми сосками или ложными сосками, которые не связаны с молочными железами. Лишь у нескольких других семейств летучих мышей есть лобковые соски, включая Hipposideridae, Craseonycteridae, Megadermatidae и Rhinopomatidae; они служат точками крепления для своего потомства. [ 19 ] У некоторых видов подковоносов у самцов в каждой подмышке имеется ложный сосок. [ 10 ]
Голова и зубы
[ редактировать ]
У всех подковоносов на носу имеются крупные листовидные выступы, которые называются носовыми листками . [ 10 ] Носовые листья важны для идентификации видов и состоят из нескольких частей. [ 20 ] Передняя часть носового листа напоминает подкову и называется ею , за что они получили общее название «подковообразные летучие мыши». [ 10 ] Подкова находится над верхней губой, тонкая и плоская. Ланцет треугольной формы, заостренный, с карманом и направлен вверх между глазами летучих мышей. [ 20 ] Седло представляет собой плоскую, напоминающую гребень структуру в центре носа. Он поднимается из-за ноздрей и направлен перпендикулярно голове. [ 20 ] Уши у них большие, листовидные, почти такие же широкие, как и длинные, без траги . Антитраги . ушей заметны Их глаза очень маленькие. [ 10 ] Череп всегда имеет ростральное вздутие или костный выступ на морде. Типичная зубная формула подковообразной летучей мыши — 1.1.2.3 2.1.3.3 , но средние нижние премоляры часто отсутствуют, а также передние верхние премоляры (премоляры ближе к передней части рта). [ 2 ] : xi Молодые теряют молочные зубы еще в утробе матери, [ 18 ] зубы резорбировались в тело. [ 21 ] Они рождаются с прорезавшимися четырьмя постоянными клыками, что позволяет им цепляться за свою мать. [ 21 ] Это нетипично для семей летучих мышей, поскольку у большинства новорожденных при рождении есть хотя бы несколько молочных зубов, которые быстро заменяются постоянными. [ 22 ]
Посткрания
[ редактировать ]Несколько костей грудной клетки срослись — предгрудинная кость, первое ребро , частично второе ребро, седьмой шейный позвонок , первый грудной позвонок — образуя сплошное кольцо. [ 2 ] : xi Это слияние связано со способностью эхолокации в неподвижном состоянии. [ 23 ] За исключением первой цифры , имеющей две фаланги , [ 18 ] все пальцы ног имеют три фаланги. [ 2 ] : xi Это отличает их от гиппосидерид, у которых на всех пальцах ног имеется по две фаланги. [ 10 ] Хвост полностью заключен в уропатагиум (хвостовую перепонку). [ 2 ] : xi а задний край уропатагиума имеет калькары ( хрящевые отростки). [ 10 ]
Биология и экология
[ редактировать ]Эхолокация и слух
[ редактировать ]
У подковоносов очень маленькие глаза, а поле зрения ограничено большими носовыми листьями; таким образом, зрение вряд ли будет очень важным чувством. Вместо этого они используют эхолокацию для навигации. [ 14 ] используя самую сложную эхолокацию среди всех групп летучих мышей. [ 24 ] Для эхолокации они производят звук через ноздри. В то время как некоторые летучие мыши используют эхолокацию с частотной модуляцией , подковоносы используют эхолокацию с постоянной частотой (также известную как одночастотная эхолокация). [ 25 ] У них высокий рабочий цикл , а это означает, что когда люди звонят, они издают звук более 30% времени. Использование высокопроизводительной эхолокации с постоянной частотой помогает различать объекты добычи по размеру. Эти характеристики эхолокации типичны для летучих мышей, которые ищут движущуюся добычу в захламленной среде, полной листвы. [ 24 ] Они осуществляют эхолокацию на особенно высоких для летучих мышей частотах, хотя и не таких высоких, как у гиппосидридов, относительно размеров их тела, и большинство из них концентрируют большую часть энергии эхолокации во второй гармонике. Королевский подковонос ( R. rex ) и большеухий подковонос ( R. philippensis ) являются примерами особенных видов, которые концентрируют энергию в первой гармонике, а не во второй. [ 26 ] Их сильно бороздчатые носовые листья, вероятно, помогают фокусировать излучение звука, уменьшая эффект беспорядка в окружающей среде. [ 25 ] Носовое полотно в целом действует как параболический отражатель , направляя производимый звук и одновременно защищая ухо от его части. [ 10 ]
Подковоносы обладают развитым слухом благодаря хорошо развитой улитке . [ 10 ] и способны обнаруживать эхо с доплеровским сдвигом . Это позволяет им одновременно производить и получать звуки. [ 2 ] : xi У подковоносов существует отрицательная взаимосвязь между длиной ушей и частотой эхолокации: виды с более высокими частотами эхолокации, как правило, имеют более короткую длину ушей. [ 26 ] Во время эхолокации уши могут двигаться независимо друг от друга характерным для семейства «мерцающим» движением, а голова одновременно движется вверх-вниз или из стороны в сторону. [ 10 ]
Диета и добыча пищи
[ редактировать ]
Подковоносы насекомоядны, хотя поедают и других членистоногих, например пауков . [ 18 ] и использовать две основные стратегии поиска пищи. Первая стратегия — летать медленно и низко над землей, охотясь среди деревьев и кустов. Некоторые виды, использующие эту стратегию, способны парить над добычей и подбирать ее из субстрата . Другая стратегия известна как кормление окуня: особи устраиваются на насестах и ждут, пока добыча пролетит мимо, а затем вылетают, чтобы поймать ее. [ 2 ] : xi Добыча пищи обычно происходит на высоте 5,0–5,9 м (16,5–19,5 футов) над землей. [ 14 ] В то время как вечерние летучие мыши могут ловить добычу своей уропатагией и переносить ее в рот, подковоносы не используют свою уропатагию для ловли добычи. По крайней мере, один вид, большая подковоносная летучая мышь, был зарегистрирован, ловя добычу кончиком своего крыла, сгибая вокруг нее фаланги, а затем перенося ее в рот. [ 10 ] [ 28 ] В то время как большинство подковоносов ведут ночной образ жизни и охотятся ночью, подковонос Блита ( R. lepidus известно, что ) добывает корм в дневное время на острове Тиоман . Предполагается, что это является ответом на отсутствие дневных птиц- хищников . на острове [ 29 ]
У них особенно маленькие и закругленные законцовки крыльев, низкая нагрузка на крыло (то есть у них большие крылья по отношению к массе тела) и высокий развал . Эти факторы придают им повышенную маневренность, и они способны совершать быстрые и крутые повороты на медленной скорости. [ 27 ] : 361 Относительно всех летучих мышей размах крыльев подковообразных типичен для размеров их тела, а соотношения сторон , которые соотносят размах крыльев с площадью крыльев, средние или ниже среднего. Некоторые виды, такие как подковоносица Рюппеля ( R. fumigatus ), подковоноска Хильдебрандта, подковоносица Лендера ( R.landeri ) и подковоносица Свинни ( R. swinny ), имеют особенно большую общую площадь крыльев, хотя большинство видов подковоносов имеют среднюю площадь крыла. [ 27 ] : 387
Размножение и жизненный цикл
[ редактировать ]Системы спаривания подковоносов изучены недостаточно. Обзор 2000 года показал, что только около 4% видов опубликовали информацию о своих системах спаривания; наряду с летучими мышами со свободным хвостом (Molossidae), они получили наименьшее внимание из всех семейств летучих мышей по сравнению с их видовым разнообразием. По крайней мере, один вид, большая подковоносная летучая мышь, по-видимому, имеет полигинную систему спаривания, при которой самцы пытаются основать и защитить территории, привлекая несколько самок. Rhinolophus sedulus , однако, является одним из немногих видов летучих мышей, которые считаются моногамными (по состоянию на 2000 год только 17 видов летучих мышей признаны таковыми). [ 30 ] У некоторых видов, особенно видов умеренного пояса , ежегодный сезон размножения приходится на осень, тогда как у других видов спаривание происходит весной. [ 18 ] Многие виды подковоносов имеют адаптацию к отсроченному оплодотворению за счет хранения спермы самок . Это особенно характерно для видов умеренного пояса. У спящих видов сроки хранения спермы совпадают со временем спячки. [ 2 ] : xi У других видов, таких как подковонос Лендера, наблюдается эмбриональная диапауза , а это означает, что, хотя оплодотворение происходит непосредственно после совокупления, зигота не имплантируется в стенку матки в течение длительного периода времени. [ 10 ] У большого подковоноса имеется адаптация задержки эмбрионального развития, то есть рост эмбриона условно задерживается, если самка впадает в спячку . Это приводит к тому, что интервал между оплодотворением и родами варьируется от двух до трех месяцев. [ 31 ] Беременность занимает примерно семь недель, прежде чем рождается один потомок, называемый щенком. Особи достигают половой зрелости к двум годам. Хотя продолжительность жизни обычно не превышает шести или семи лет, некоторые люди могут прожить необычайно долго. Большую особь подковообразной летучей мыши однажды окольцевали, а затем заново открыли тридцать лет спустя. [ 18 ]
Поведение и социальные системы
[ редактировать ]различные уровни социальности У подковоносов наблюдаются . Некоторые виды ведут одиночный образ жизни, особи ночуют поодиночке, в то время как другие являются колониальными , образуя скопления из тысяч особей. [ 2 ] : xi Большинство видов умеренно социальны. У некоторых видов полы разделяются ежегодно, когда самки образуют родильные колонии , хотя у других полы остаются вместе в течение всего года. Особи охотятся в одиночку. [ 18 ] Поскольку их задние конечности плохо развиты, они не могут бегать по ровным поверхностям и умело лазать, как другие летучие мыши. [ 14 ] [ 10 ]
Подковоносы впадают в спячку для сохранения энергии. Во время оцепенения температура их тела падает до 16 ° C (61 ° F), а скорость метаболизма замедляется. [ 32 ] Торпор используется подковоносами в умеренных, субтропических и тропических регионах. [ 33 ] Оцепенение имеет короткую продолжительность; когда оцепенение применяется постоянно в течение нескольких дней, недель или месяцев, это называется спячкой. [ 34 ] Спячка используется подковоносами в регионах с умеренным климатом в зимние месяцы. [ 33 ]
Хищники и паразиты
[ редактировать ]В целом у летучих мышей мало естественных хищников. [ 35 ] К хищникам подковоносов относятся птицы отряда Accipitriformes (ястребы, орлы и коршуны), а также соколы и совы . [ 36 ] [ 37 ] Змеи также могут охотиться на некоторые виды, пока они ночуют в пещерах. [ 38 ] и домашние кошки тоже могут охотиться на них. [ 39 ] Исследование, проведенное в 2019 году возле колонии летучих мышей в центральной Италии, показало, что 30% исследованных фекалий кошек содержат останки больших подковоносов. [ 40 ]
Подковоносы имеют множество внутренних и внешних паразитов . К наружным паразитам (эктопаразитам) относятся клещи рода Eyndhovenia , «летучие мыши» семейств Streblidae и Nycteribiidae , [ 41 ] клещи рода Ixodes , [ 42 ] и блохи рода Rhinolophopsylla . [ 43 ] На них также поражаются разнообразные внутренние паразиты (эндопаразиты), в том числе трематоды родов Lecithodendrium , Plagiorchis , Prosthodendrium , [ 44 ] and cestodes of the genus Potorolepsis . [ 45 ]
Ареал и среда обитания
[ редактировать ]Подковоносы имеют в основном палеотропическое распространение, хотя некоторые виды обитают в южной Палеарктике . [ 13 ] Они встречаются в Старом Свете , включая Африку, Австралию, Азию, Европу и Океанию. [ 9 ] Большой подковонос имеет самый большой географический ареал среди всех подковоносов: он встречается в Европе, Северной Африке, Японии, Китае и Южной Азии. Другие виды гораздо более ограничены, например, Андаманская подковоносная летучая мышь ( R. cognatus ), которая встречается только на Андаманских островах . [ 14 ] Они ночуют в самых разных местах, включая здания, пещеры, дупла деревьев и листву. Встречаются как в лесных, так и в безлесных местах обитания. [ 18 ] при этом большинство видов встречается в тропических или субтропических районах. [ 10 ] Для видов, впадающих в спячку, они выбирают пещеры с температурой окружающей среды примерно 11 °C (52 °F). [ 46 ]
Отношение к людям
[ редактировать ]Как резервуары болезней
[ редактировать ]Коронавирусы
[ редактировать ]| Виды летучих мышей | От SARSr-CoVs |
|---|---|
| Китайская рыжая подковоносная летучая мышь | |
| Большая подковообразная летучая мышь | |
| Ушастая подковоносная летучая мышь | |
| Наименьшая подковообразная летучая мышь | |
| Промежуточная подковообразная бита | |
| Подковообразная летучая мышь Блазиуса | |
| Трезубец Столички | |
| Летучая мышь со свободным хвостом и морщинистыми губами |

Подковоносы представляют особый интерес для общественного здравоохранения и зоонозов как источник коронавирусов .
После вспышки атипичной пневмонии в 2002–2004 годах несколько видов животных были исследованы как возможные естественные резервуары возбудителя коронавируса SARS-CoV . С 2003 по 2018 год у подковоносов было обнаружено сорок семь коронавирусов, связанных с атипичной пневмонией. [ 47 ] В 2019 году мокрый рынок в Ухане , Китай, был связан со вспышкой SARS-CoV-2 . Генетический анализ SARS-CoV-2 показал, что он очень похож на вирусы, обнаруженные у подковоносов. [ 48 ]
После вспышки атипичной пневмонии наименее подковоносная летучая мышь ( R. pusillus ) оказалась серопозитивной, большая подковоносная летучая мышь получила положительный результат только на вирус, а ушастая подковоносная летучая мышь ( R. macrotis ) и китайская рыжая подковоносная летучая мышь ( R. sinicus ) и подкова Пирсона ( R. pearsoni ) оказались серопозитивными и дали положительный результат на вирус. [ 47 ] [ 49 ] Вирусы летучих мышей были очень похожи на SARS-CoV — сходство 88–92%. [ 1 ] Внутривидовое разнообразие SARS -подобных коронавирусов, по-видимому, возникло у Rhinolophus sinicus в результате гомологичной рекомбинации . [ 50 ] R. sinicus, вероятно, является носителем прямого предка SARS-CoV у людей. Хотя подковоносы оказались естественным резервуаром коронавирусов, связанных с атипичной пневмонией, люди, вероятно, заболели в результате контакта с инфицированными маскированными пальмовыми циветтами , которые были идентифицированы как промежуточные хозяева вируса. [ 1 ]
За период с 2003 по 2018 год у летучих мышей выявлено сорок семь коронавирусов, связанных с атипичной пневмонией, сорок пять — у подковоносов. Тридцать связанных с атипичной пневмонией коронавирусов были получены от китайских рыжих подковоносов, девять – от больших подковоносов, два – от ушастых подковоносов, два – от наименьших подковоносов и по одному – от промежуточной подковоносы ( R. affinis ), подковоноса Блазиуса. ( R. blasii ), трезубец Столички ( Aselliscus stoliczkanus ) и морщинистогубая летучая мышь свободнохвостая летучая мышь ( Chaerephon plicata ). [ 47 ]
На рынке в Ухане, где был обнаружен SARS-CoV-2, 96% имели сходство с вирусом, выделенным из подковоноса . Исследования эволюционного происхождения SARS-CoV-2 [ 51 ] указывает на то, что летучие мыши были естественными резервуарами SARS-CoV-2. Пока неясно, как вирус передался человеку, хотя, возможно, в этом участвовал промежуточный хозяин. Когда-то считалось, что это панголин Сунда . [ 52 ] но публикация в июле 2020 года не обнаружила никаких доказательств передачи вируса от панголинов человеку. [ 51 ]
Другие вирусы
[ редактировать ]Они также связаны с такими вирусами, как ортореовирусы , флавивирусы и хантавирусы . Они дали положительный результат на ортореовирус млекопитающих (MRV), включая MRV типа 1, выделенный от подковоноса, и MRV типа 2, выделенный от подковоноса. Конкретные MRV, обнаруженные у подковоносов, не связаны с заражением человека, хотя люди могут заболеть в результате воздействия других MRV. [ 53 ] Рыжая подковоносная летучая мышь ( R. rouxii ) показала серопозитивный результат на болезнь Кьясанурского леса , которая представляет собой клещевую вирусную геморрагическую лихорадку, известную из южной Индии. Болезнь Кьясанурского леса передается человеку через укусы зараженных клещей, уровень смертности составляет 2–10%. [ 54 ] Вирус Лунцюань , разновидность хантавируса, был обнаружен у подковоноса среднего размера, китайской рыжей подковоноса и маленькой японской подковоноса ( R. cornutus ). [ 55 ]
Как еда и лекарство
[ редактировать ]На микролетучих мышей охотятся не так интенсивно, как на мегалетучих мышей : только 8% насекомоядных видов охотятся ради еды, по сравнению с половиной всех видов летучих мышей в тропиках Старого Света. На подковоносов охотятся ради еды, особенно в странах Африки к югу от Сахары . Виды, на которые охотятся в Африке, включают безмятежного подковоноса ( R. alcyone ), гвинейского подковоноса ( R. guineensis ), подковоноса Хилла ( R. Hilli ), подковоноса Хилла ( R. Hillorum ), подковоноса Маклауда ( R. maclaudi ). ), подковонос Рувензори, подковонос лесной ( R. silvestris ) и подковонос Зиама летучая мышь ( Р. ziama ). В Юго-Восточной Азии подковонос Маршалла ( R. marshalli ) употребляется в пищу в Мьянме, а большой рыжий подковонос ( R. rufus ) – на Филиппинах. [ 56 ]
Сообщается, что народность ао-нага на северо-востоке Индии использует мясо подковоносов для лечения астмы . Экологический антрополог Уилл Туладхар-Дуглас заявил, что неварцы в Непале «почти наверняка» используют подковоносов, среди других видов, для приготовления Cika Lāpa Wasa («масла для летучих мышей»). Мертвых летучих мышей сворачивают и помещают в плотно закрытые банки с горчичным маслом ; масло готово, когда оно издает отчетливый и неприятный запах. Традиционное медицинское применение масла летучих мышей включает удаление «ушных жуков», которые, как сообщается, представляют собой многоножек , которые заползают в уши и грызут мозг, что, возможно, является традиционным объяснением мигрени . Он также используется в качестве предполагаемого лечения облысения и частичного паралича. [ 57 ] В Сенегале имеются отдельные сообщения об использовании подковоносов в зельях для лечения психических заболеваний; Во Вьетнаме фармацевтическая компания сообщила, что использует 50 т (50 000 кг) гуано подковообразной летучей мыши в медицинских целях. ежегодно [ 58 ]
Сохранение
[ редактировать ]По состоянию на 2023 год МСОП оценил 94 вида подковоносов. Они имеют следующие статусы МСОП: [ 59 ]
- Находится под угрозой исчезновения : 1 вид ( подковонос Хилла ).
- Под угрозой исчезновения : 13 видов.
- Уязвимые : 5 видов
- Находящиеся под угрозой исчезновения : 9 видов
- Наименее опасные : 51 вид.
- Недостаточно данных : 15 видов
Как и все летучие мыши, обитающие в пещерах, подковоносы, обитающие в пещерах, уязвимы к нарушению среды их пещерной среды обитания. Беспокойство может включать добычу гуано летучих мышей , добычу известняка и пещерный туризм . [ 46 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с Ван, Линь-Фа; Ши, Чжэнли; Чжан, Шуи; Филд, Хьюм; Дашак, Питер; Итон, Брайан (2006). «Обзор летучих мышей и атипичной пневмонии» . Новые инфекционные заболевания . 12 (12): 1834–1840. дои : 10.3201/eid1212.060401 . ПМК 3291347 . ПМИД 17326933 .
- ^ Jump up to: а б с д и ж г час я дж к л м н Чорба, Г.; Уджели, П.; Томас, П. (2003). Подковоносы мира: (Chiroptera: Rhinolophidae) . Книги Аланы. ISBN 9780953604913 .
- ^ Грей, Дж. Э. (1825). «Попытка разделения семейства Vespertilionidae на группы» . Зоологический журнал . 2 : 242.
- ^ Jump up to: а б Маккенна, MC; Белл, СК (1997). Классификация млекопитающих: выше уровня вида . Издательство Колумбийского университета. п. 305. ИСБН 9780231528535 .
- ^ Тейлор, Питер Дж.; Стоффберг, Саманта; Монадджем, Ара; Шуман, Мартинус Корри; Бэйлисс, Джулиан; Коттерилл, Фентон, полиция (2012). «Четыре новых вида летучих мышей (комплекс Rhinolophus hildebrandtii) отражают плио-плейстоценовое расхождение карликов и гигантов на Афромонтном архипелаге» . ПЛОС ОДИН . 7 (9): е41744. Бибкод : 2012PLoSO...741744T . дои : 10.1371/journal.pone.0041744 . ПМК 3440430 . ПМИД 22984399 .
- ^ «Семейство Ринолофиды» . Виды млекопитающих мира . Бакнеллский университет . Проверено 23 июля 2020 г.
- ^ Спрингер, М.С.; Тилинг, ЕС; Мэдсен, О.; Стэнхоуп, MJ; Де Йонг, WW (2001). «Комплексные ископаемые и молекулярные данные реконструируют эхолокацию летучих мышей» . Труды Национальной академии наук . 98 (11): 6241–6246. Бибкод : 2001PNAS...98.6241S . дои : 10.1073/pnas.111551998 . ПМК 33452 . ПМИД 11353869 .
- ^ Jump up to: а б Амадор, Л.И.; Аревало, RLM; Алмейда, ФК; Каталано, ЮАР; Джаннини, НП (2018). «Систематика летучих мышей в свете неограниченного анализа комплексной молекулярной суперматрицы». Журнал эволюции млекопитающих . 25 : 37–70. дои : 10.1007/s10914-016-9363-8 . hdl : 11336/55671 . S2CID 3318167 .
- ^ Jump up to: а б с д и Стоффберг, Саманта; Джейкобс, Дэвид С.; Макки, Иэн Дж.; Мэти, Конрад А. (2010). «Молекулярная филогенетика и историческая биогеография летучих мышей Rhinolophus». Молекулярная филогенетика и эволюция . 54 (1): 1–9. дои : 10.1016/j.ympev.2009.09.021 . ПМИД 19766726 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот Хаппольд, М.; Коттерилл, FPD (2013). Кингдон, Дж.; Хаппольд, Д.; Бутынский Т.; Хоффманн, М.; Хаппольд, М.; Калина Дж. (ред.). Млекопитающие Африки . Том. 4. А&С Черный. стр. 300–303. ISBN 9781408189962 .
- ^ Уилсон, Дон Э.; Ридер, ДиЭнн М., ред. (2005). «Семейство Hipposideridae» . Виды млекопитающих мира (3-е изд.). Бакнеллский университет . Проверено 23 июля 2020 г.
- ^ Воллет, Марианна; Лойдл, Йозеф; Майер, Фридер; Ён, Хой-Сен; Мюллер, Стефан; Хеллер, Клаус-Герхард (2015). «Удивительное генетическое разнообразие Rhinolophus luctus (Chiroptera: Rhinolophidae) с полуострова Малайзия: описание нового вида на основе генетических и морфологических признаков» . Acta Chiropterologica . 17 :1–20. дои : 10.3161/15081109ACC2015.17.1.001 . S2CID 86009452 .
- ^ Jump up to: а б с д Демос, Терренс К.; Вебала, Пол В.; Гудман, Стивен М.; Кербис Петеранс, Джулиан К.; Бартонджо, Майкл; Паттерсон, Брюс Д. (2019). «Молекулярная филогенетика африканских подковоносов (Chiroptera: Rhinolophidae): расширенная географическая и таксономическая выборка афротропиков» . Эволюционная биология BMC . 19 (1): 166. Бибкод : 2019BMCEE..19..166D . дои : 10.1186/s12862-019-1485-1 . ПМК 6704657 . ПМИД 31434566 .
- ^ Jump up to: а б с д и ж г Ум, В.; Клейман, Д.Г.; Макдейд, MC (2004). Энциклопедия жизни животных Гржимека Млекопитающие II . Том. 13 (2-е изд.). Гейл . стр. 387–393. ISBN 978-0787657895 .
- ^ Палмер, Т.С. (1904). «Список родов и семейств млекопитающих» . Североамериканская фауна (23): 503.
- ^ Лидеккер, Ричард (1885). Каталог ископаемых млекопитающих в Британском музее (Естественная история): отряды Primates, Chiroptera, Insectivora, Carnivora и Rodentia . Приказ попечителей. п. 13.
- ^ Богданович, В.; Оуэн, Р.Д. (1992). «Филогенетический анализ семейства летучих мышей Rhinolophidae» (PDF) . Журнал зоологической систематики и эволюционных исследований . 30 (2): 152. doi : 10.1111/j.1439-0469.1992.tb00164.x .
Единственный ископаемый род, Palaeonycteris , известен из миоцена Европы (Heller 1936; Sigb and Legendre 1983; Hand 1984; ср. Simpson 1945 и Hall 1989).
- ^ Jump up to: а б с д и ж г час я Новак, Рональд М. (1994). Летучие мыши мира Уокера . Джу Пресс. С. 108–110 . ISBN 978-0-8018-4986-2 .
- ^ Симмонс, Н.Б. (1993). «Морфология, функция и филогенетическое значение лобковых сосков летучих мышей (Mammalia, Chiroptera)» (PDF) . Американский музей послушников (3077 г.).
- ^ Jump up to: а б с Холл, Лесли (1989). «Ринолофиды». В Уолтоне, Д.В.; Ричардсон, Би Джей (ред.). Фауна Австралии (PDF) . АГПС Канберра.
- ^ Jump up to: а б Хермансон, Дж.В.; Вудс, Калифорния; Хауэлл, К.М. (1982). «Онтогенез зубов листоносых летучих мышей Старого Света (Rhinolophidae, Hipposiderinae)». Журнал маммологии . 63 (3): 527–529. дои : 10.2307/1380461 . JSTOR 1380461 .
- ^ Воган, Т. (1970). «Глава 3: Скелетная система». В Вимсатте, В. (ред.). Биология летучих мышей . Академическая пресса . стр. 103–136. ISBN 9780323151191 .
- ^ Стоффберг, Саманта; Джейкобс, Дэвид С.; Макки, Иэн Дж.; Мэти, Конрад А. (2010). «Молекулярная филогенетика и историческая биогеография летучих мышей Rhinolophus». Молекулярная филогенетика и эволюция . 54 (1): 1–9. дои : 10.1016/j.ympev.2009.09.021 . ПМИД 19766726 .
- ^ Jump up to: а б Джонс, Г.; Тилинг, Э. (2006). «Эволюция эхолокации у летучих мышей». Тенденции в экологии и эволюции . 21 (3): 149–156. дои : 10.1016/j.tree.2006.01.001 . ПМИД 16701491 .
- ^ Jump up to: а б Вандерельст, Дитер; Йонас, Рейньерс; Герберт, Переманс (2012). «Возвращение к бороздам Rhinolophidae» . Журнал интерфейса Королевского общества . 9 (70): 1100–1103. дои : 10.1098/rsif.2011.0812 . ПМК 3306658 . ПМИД 22279156 .
- ^ Jump up to: а б Хуэйхуа, Чжао; Шуйи, Чжан; Минсюэ, Цзо; Цзян, Чжоу (2003). «Корреляция между частотой криков и длиной ушей у летучих мышей, принадлежащих к семействам Rhinolophidae и Hipposideridae». Журнал зоологии . 259 (2): 189–195. дои : 10.1017/S0952836902003199 .
- ^ Jump up to: а б с Норберг, UM; Рейнер, JMV (1987). «Экологическая морфология и полет летучих мышей (Mammalia; Chiroptera): адаптации крыльев, летные характеристики, стратегия кормления и эхолокация» . Философские труды Лондонского королевского общества B: Биологические науки . 316 (1179): 335–427. Бибкод : 1987РСТБ.316..335Н . дои : 10.1098/rstb.1987.0030 .
- ^ Вебстер, Фредерик А.; Гриффин, Дональд Р. (1962). «Роль летных перепонок в захвате насекомых летучими мышами». Поведение животных . 10 (3–4): 332–340. дои : 10.1016/0003-3472(62)90056-8 .
- ^ Чуа, Маркус А.Х.; Азиз, Шима Абдул (19 декабря 2018 г.). «На свет: нетипичная дневная кормовая деятельность подковообразной летучей мыши Блита, Rhinolophus lepidus (Chiroptera: Rhinolophidae) на острове Тиоман, Малайзия» . Млекопитающие . 83 (1): 78–8 дои : 10.1515/млекопитающее-2017-0128 . ISSN 1864-1547 . S2CID 90531252 .
- ^ Маккракен, Гэри Ф.; Уилкинсон, Джеральд С. (2000). «Системы спаривания летучих мышей». Репродуктивная биология летучих мышей . стр. 321–362. дои : 10.1016/B978-012195670-7/50009-6 . ISBN 9780121956707 .
- ^ Гайслер, Дж. (2013). Кингдон, Дж.; Хаппольд, Д.; Бутынский Т.; Хоффманн, М.; Хаппольд, М.; Калина Дж. (ред.). Млекопитающие Африки . Том. 4. А&С Черный. стр. 327–328. ISBN 9781408189962 .
- ^ Гейзер, Фриц; Кёртнер, Герхард (2010). «Спячка и суточное оцепенение австралийских млекопитающих» . Австралийский зоолог . 35 (2): 204–215. дои : 10.7882/AZ.2010.009 . hdl : 10072/37379 .
- ^ Jump up to: а б Гейзер, Ф.; Ставски, К. (2011). «Спячка и оцепенение тропических и субтропических летучих мышей в связи с энергетикой, вымиранием и эволюцией эндотермии» . Интегративная и сравнительная биология . 51 (3): 337–348. дои : 10.1093/icb/icr042 . ПМИД 21700575 .
- ^ Альтрингем, Джон Д. (2011). Летучие мыши: от эволюции к сохранению . Издательство Оксфордского университета. п. 99. ИСБН 9780191548727 .
- ^ Ниффелер, Мартин; Кнёрншильд, Мирьям (2013). «Хищничество пауков на летучих мышей» . ПЛОС ОДИН . 8 (3): e58120. Бибкод : 2013PLoSO...858120N . дои : 10.1371/journal.pone.0058120 . ПМЦ 3596325 . ПМИД 23516436 .
- ^ Микула, Питер; Морелли, Федерико; Лучан, Радек К.; Джонс, Дэррил Н.; Тряновский, Петр (2016). «Летучие мыши как добыча дневных птиц: глобальная перспектива: хищничество летучих мышей дневных птиц». Обзор млекопитающих . 46 (3): 160–174. дои : 10.1111/мам.12060 .
- ^ Гарсия, AM; Сервера, Ф.; Родригес, А. (2005). «Хищничество ушастых сов на летучих мышей в средиземноморских и умеренных регионах южной Европы» (PDF) . Журнал исследований хищников . 39 (4): 445–453.
- ^ Барти, Левенте; Питер, Аарон; Чос, Иштван; Шандор, Аттила Д. (2019). «Нападение змей на летучих мышей в Европе: новые случаи и региональная оценка» (PDF) . Млекопитающие . 83 (6): 581–585. doi : 10.1515/mammalia-2018-0079 . S2CID 92282216 .
- ^ Анчиллотто, Леонардо; Серанджели, Мария Тициана; Руссо, Данило (2013). «Любопытство убило летучую мышь: домашние кошки как хищники летучих мышей». Биология млекопитающих . 78 (5): 369–373. Бибкод : 2013MamBi..78..369A . дои : 10.1016/j.mambio.2013.01.003 .
- ^ Анчиллотто, Л.; Вентури, Г.; Руссо, Д. (2019). «Присутствие людей и домашних кошек влияет на поведение летучих мышей в городском питомнике больших подковоносов ( Rhinolophusferrumequinum )». Поведенческие процессы . 164 : 4–9. doi : 10.1016/j.beproc.2019.04.003 . ПМИД 30951813 . S2CID 92844287 .
- ^ Шарифи, Мозафар; Тагинежад, Найме; Мосафар, Фатима; Вайси, Некоторые (2013). «Изменение нагрузки эктопаразитов у подковоносов Мехели, Rhinolophus mehelyi (Chiroptera: Rhinolophidae) в детской колонии в западном Иране». Акта Паразитологическая . 58 (2): 180–184. дои : 10.2478/ s11686-013-0122-1 ПМИД 23666653 . S2CID 7173658 .
- ^ Хорнок, Шандор; Гёрфёл, Тамаш; Эсток, Питер; Ту, Выонг Тан; Кончан, Йено (2016). «Описание нового вида клещей Ixodes Colliris n. sp. (Acari: Ixodidae) от летучих мышей (Chiroptera: Hipposideridae, Rhinolophidae) во Вьетнаме» . Паразиты и переносчики . 9 (1): 332. дои : 10.1186/s13071-016-1608-0 . ПМК 4902904 . ПМИД 27286701 .
- ^ Котти, БК (2018). «Распространение и специфика паразитарных ассоциаций блох (Siphonaptera) на Центральном Кавказе». Энтомологический обзор . 98 (9): 1342–1350. дои : 10.1134/S0013873818090129 . S2CID 85527706 .
- ^ Хорват, Ж.; Чабрило, Б.; Паунович, М.; Карапанджа, Б.; Йованович, Дж.; Будинский И.; Бьелич Чабрило, О. (2017). «Желудочно-кишечные дигенеи (Platyhelminthes: Trematoda) подковообразных и вечерних летучих мышей (Chiroptera: Rhinolophidae и Vespertilionidae) в Сербии» . Гельминтология . 54 : 17–25. дои : 10.1515/helm-2017-0009 . S2CID 90530235 .
- ^ Макарикова, Т. А.; Макариков А.А. (2012). «Первое сообщение о Potorolepis Spassky, 1994 (Eucestoda: Hymenolepididae) из Китая, с описанием нового вида летучих мышей (Chiroptera:: Rhinolophidae)» . Фолиа Паразитологическая . 59 (4): 272–278. дои : 10.14411/fp.2012.038 . ПМИД 23327008 .
- ^ Jump up to: а б Фьюри, Нил М.; Рэйси, Пол А. (2016). «Экология сохранения пещерных летучих мышей» (PDF) . Летучие мыши в антропоцене: сохранение летучих мышей в меняющемся мире . Спрингер, Чам. стр. 463–500. дои : 10.1007/978-3-319-25220-9_15 . ISBN 978-3-319-25218-6 .
- ^ Jump up to: а б с д Люк, Хейс К.Х.; Ли, Синь; Фунг, Джошуа; Лау, Сюзанна КП; Ву, Патрик Сай (2019). «Молекулярная эпидемиология, эволюция и филогения коронавируса SARS» . Инфекция, генетика и эволюция . 71 : 21–30. Бибкод : 2019InfGE..71...21L . дои : 10.1016/j.meegid.2019.03.001 . ПМК 7106202 . ПМИД 30844511 .
- ^ «Отчет о ситуации с новым коронавирусом (2019-nCoV)» (PDF) . Всемирная организация здравоохранения . 11 февраля 2020 г. Проверено 15 февраля 2020 г. .
- ^ Ши, Чжэнли; Ху, Чжихун (2008). «Обзор исследований на животных-резервуарах коронавируса SARS» . Вирусные исследования . 133 (1): 74–87. doi : 10.1016/j.virusres.2007.03.012 . ПМК 7114516 . ПМИД 17451830 .
- ^ Юань Дж., Хон CC, Ли Ю, Ван Д., Сюй Г., Чжан Х., Чжоу П., Пун Л.Л., Лам Т.Т., Люнг Ф.К., Ши З. Внутривидовое разнообразие SARS-подобных коронавирусов у Rhinolophus sinicus и его значение для происхождения Коронавирусов SARS у человека. Джей Ген Вирол. 91 апреля 2010 г. (Часть 4): 1058–62. дои: 10.1099/vir.0.016378-0. Epub 2009, 16 декабря. ПМИД 20016037
- ^ Jump up to: а б Бони, Мацей Ф.; Леми, Филипп; Цзян, Сяовэй; Лам, Томми Цан-Юк; Перри, Блэр В.; Касто, Тодд А.; Рамбо, Эндрю; Робертсон, Дэвид Л. (2020). «Эволюционное происхождение линии сарбековируса SARS-CoV-2, ответственной за пандемию COVID-19» . Природная микробиология . 5 (11): 1408–1417. дои : 10.1038/s41564-020-0771-4 . hdl : 20.500.11820/222bb9b9-2481-4086-bd22-f0b200930bef . ПМИД 32724171 . S2CID 220809302 .
- ^ Маккензи, Джон С.; Смит, Дэвид В. (2020). «COVID-19: новое зоонозное заболевание, вызванное коронавирусом из Китая: что мы знаем, а что нет» . Микробиология Австралии . 41 : 45. дои : 10.1071/MA20013 . ПМК 7086482 . ПМИД 32226946 .
Данные анализа последовательностей четко указывают на то, что резервуарным хозяином вируса была летучая мышь, вероятно, китайская или промежуточная подковоносная летучая мышь, и вполне вероятно, что, как и в случае с SARS-CoV, источником вспышки был промежуточный хозяин.
- ^ Бельц, Лиза А. (2017). Летучие мыши и здоровье человека: Эбола, атипичная пневмония, бешенство и не только . Джон Уайли и сыновья. п. 155. ИСБН 9781119150046 .
- ^ Паттнаик, Приябрата (2006). «Лесная болезнь Кьясанур: эпидемиологический взгляд в Индии». Обзоры по медицинской вирусологии . 16 (3): 151–165. дои : 10.1002/rmv.495 . ПМИД 16710839 . S2CID 32814428 .
- ^ Го, Вэнь-Пин; Ван, Цзюнь-Хуа; Чжан, Хай-Линь; Чжоу, Жунь-Хун; Бо; Ли, Мин-Хуэй; Сюй, Цзянго; Эдвард Чжан , Юн-Чжэнь (2013). Цзянь - Летучие мыши, насекомоядные и грызуны» . PLOS Pathogens . 9 (2): e1003159. : 10.1371 /journal.ppat.1003159 . PMC 3567184. . PMID 23408889 doi
- ^ Милденштейн, Т.; Танши, И.; Рэйси, Пенсильвания (2016). «Эксплуатация летучих мышей для добычи мяса диких животных и лекарств». Летучие мыши в антропоцене: сохранение летучих мышей в меняющемся мире . Спрингер. п. 327. дои : 10.1007/978-3-319-25220-9_12 . ISBN 978-3-319-25218-6 . S2CID 130038936 .
- ^ Туладхар-Дуглас, Уилл (2008). «Использование летучих мышей в качестве лекарства среди неваров». Журнал этнобиологии . 28 : 69–91. doi : 10.2993/0278-0771(2008)28[69:TUOBAM]2.0.CO;2 . ISSN 0278-0771 . S2CID 146308445 .
- ^ Рикуччи, М. (2012). «Летучие мыши как Материя медика: этномедицинский обзор и значение для сохранения» . Веспертилио . 16 (16): 249–270.
- ^ «Таксономия=Ринолофиды» . МСОП . Проверено 28 декабря 2023 г.
| Базы данных органов управления : Национальные |
|---|