Однодоменное антитело

Расширенный цикл CDR3 окрашен в оранжевый цвет.
Однодоменное антитело ( sdAb ), также известное как нанотело , представляет собой фрагмент антитела, состоящий из одного мономерного вариабельного домена антитела . Как и целое антитело, оно способно избирательно связываться со специфическим антигеном . При молекулярной массе всего 12–15 кДа однодоменные антитела намного меньше обычных антител (150–160 кДа), которые состоят из двух тяжелых белковых цепей и двух легких цепей , и даже меньше, чем Fab-фрагменты (~ 50 кДа). , одна легкая цепь и половина тяжелой цепи) и одноцепочечные вариабельные фрагменты (~25 кДа, два вариабельных домена, один из легкой и один из тяжелой цепи). [1]
Первые однодоменные антитела были созданы на основе антител с тяжелой цепью, обнаруженных у верблюдовых ; называются VHH фрагментами они . У хрящевых рыб однодоменные антитела, называемые V NAR . фрагментами также есть антитела с тяжелой цепью (IgNAR, «рецептор нового антигена иммуноглобулина»), из которых можно получить [2] Альтернативный подход заключается в разделении димерных вариабельных доменов общего иммуноглобулина G (IgG) человека или мышей на мономеры. Хотя большинство исследований однодоменных антител в настоящее время основано на вариабельных доменах тяжелых цепей, также было показано, что нанотела, полученные из легких цепей, специфически связываются с целевыми эпитопами . [3]
Было показано, что нанотела верблюдовых столь же специфичны, как и антитела, а в некоторых случаях более надежны. Их легко выделить с помощью той же процедуры фагового пэннинга, что и для антител, что позволяет культивировать их in vitro в больших концентрациях. Меньший размер и один домен облегчают трансформацию этих антител в бактериальные клетки для массового производства, что делает их идеальными для исследовательских целей. [4]
Однодоменные антитела исследуются для различных фармацевтических применений и имеют потенциал для использования в лечении острого коронарного синдрома , рака , болезни Альцгеймера , [5] [6] и Ковид-19 . [7] [8] [9]
Характеристики
[ редактировать ]Однодоменное антитело представляет собой пептидную цепь длиной около 110 аминокислот , содержащую один вариабельный домен (VH ) антитела с тяжелой цепью или обычного IgG. Эти пептиды имеют такое же сродство к антигенам, как и цельные антитела, но более термостойки и стабильны к детергентам и высоким концентрациям мочевины . Антитела, полученные из верблюжьих и рыбных антител, менее липофильны и более растворимы в воде благодаря их определяющей комплементарность области 3 (CDR3), которая образует расширенную петлю (оранжевого цвета на ленточной диаграмме выше), покрывающую липофильный участок, который обычно связывается с легкая цепь. [10] [11] В отличие от обычных антител, два из шести однодоменных антител выдержали температуру 90 °C (194 °F), не потеряв своей способности связывать антигены в исследовании 1999 года. [12] Стабильность по отношению к желудочной кислоте и протеазам зависит от аминокислотной последовательности. Было показано, что некоторые виды активны в кишечнике после перорального применения. [13] [14] но их низкая абсорбция из кишечника препятствует развитию системно активных однодоменных антител, вводимых перорально.

Сравнительно низкая молекулярная масса приводит к лучшей проницаемости в тканях и к короткому периоду полувыведения из плазмы, поскольку они выводятся почками . [1] В отличие от цельных антител, они не проявляют системой комплемента, запускаемой цитотоксичности, поскольку у них отсутствует Fc-область . sdAb, полученные из верблюдовых и рыб, способны связываться со скрытыми антигенами, недоступными для целых антител, например, с активными центрами ферментов . [15] Было показано, что это свойство является результатом их расширенной петли CDR3, которая способна проникать в такие погребенные места. [11] [16] [15]
Производство
[ редактировать ]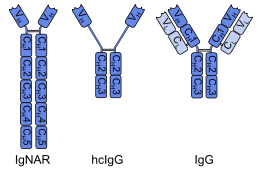
Из антител с тяжелой цепью
[ редактировать ]Однодоменное антитело можно получить путем иммунизации дромадеров , верблюдов , лам , альпак или акул желаемым антигеном и последующего выделения мРНК , кодирующей вариабельную область (V NAR и V H H) антител с тяжелой цепью. Большие фаговые V NAR и V H H были созданы у акул-нянек. однодоменные библиотеки [17] и дромадеры. [18] [19] Методы скрининга, такие как фаговый дисплей и рибосомный дисплей, помогают идентифицировать клоны, связывающие антиген. [20] [17] [21] [18] [22] [8] [19] Однодоменные антитела, включая V NAR , можно гуманизировать для клинических применений. [23]
От обычных антител
[ редактировать ]Альтернативно, однодоменные антитела могут быть получены из мышиных обычных [24] кролик [25] или человеческий IgG [26] с четырьмя цепями. [27] Процесс аналогичен: он включает библиотеки генов от иммунизированных или наивных доноров и методы отображения для идентификации наиболее специфичных антигенов. Проблема этого подхода заключается в том, что область связывания общего IgG состоит из двух доменов (VH и VL ) , которые имеют тенденцию к димеризации или агрегации из-за своей липофильности. Мономеризация обычно осуществляется путем замены липофильных аминокислот гидрофильными, но часто приводит к потере сродства к антигену. [28] Если аффинность может быть сохранена, однодоменные антитела также могут быть получены в E. coli . [25] [26] [29] S. cerevisiae или другие организмы.
Из человеческих однодоменных антител
[ редактировать ]Люди иногда производят однодоменные антитела путем случайного создания стоп-кодона в легкой цепи. Человеческие однодоменные антитела, нацеленные на различные опухолевые антигены, включая мезотелин, [29] ГПК2 [30] и GPC3 [26] [31] были выделены с помощью фагового дисплея. Человеческие однодоменные антитела HN3 были использованы для создания иммунотоксинов. [31] [32] [33] и Т-клетки химерного антигенного рецептора (CAR) [34] для лечения рака печени. Блокирование Wnt-связывающего домена GPC3 человеческим однодоменным антителом HN3 ингибирует активацию Wnt в клетках рака печени. [35]
Возможные применения
[ редактировать ]Однодоменные антитела допускают широкий спектр применения как в биотехнических, так и в терапевтических целях благодаря их небольшому размеру, простоте производства и высокой аффинности. [36] [37] [15]
Биотехнологические и диагностические
[ редактировать ]Слияние флуоресцентного белка с нанотелом приводит к образованию так называемого хромотела . Хромотела можно использовать для распознавания и отслеживания целей в различных отсеках живых клеток. Таким образом, они могут расширить возможности микроскопии живых клеток и позволят провести новые функциональные исследования. [38] Соединение наноантитела анти- GFP с моновалентной матрицей, называемой GFP-наноловушкой, позволяет изолировать слитые с GFP белки и их взаимодействующих партнеров для дальнейшего биохимического анализа. [39] Локализация одной молекулы с помощью методов визуализации сверхвысокого разрешения требует специфической доставки флуорофоров в непосредственной близости от целевого белка. Из-за их большого размера использование антител, связанных с органическими красителями, часто может привести к ошибочному сигналу из-за расстояния между флуорофором и целевым белком. Слияние органических красителей с нанотелами против GFP, нацеленными на белки, меченные GFP, обеспечивает нанометровое пространственное разрешение и минимальную ошибку связывания из-за небольшого размера и высокого сродства. [40] Дивиденд размера нанотел также полезен для исследования корреляционной светоэлектронной микроскопии . Без какого-либо пермеабилизирующего агента цитоплазма химически фиксированных клеток легко доступна для нанотел, меченных флуорофором. Их небольшой размер также позволяет им проникать глубже в объемные образцы, чем обычные антитела. Высокое качество ультраструктуры сохраняется в ткани, которую визуализируют с помощью флуоресцентного, а затем электронного микроскопа. Это особенно полезно для нейробиологических исследований, которые требуют как молекулярной маркировки, так и электронной микроскопии. [41]
В диагностических биосенсорных приложениях нанотела могут быть перспективно использованы в качестве инструмента. Благодаря небольшому размеру их можно более плотно сцеплять на поверхностях биосенсоров. Помимо их преимущества в нацеливании на менее доступные эпитопы, их конформационная стабильность также приводит к более высокой устойчивости к условиям регенерации поверхности. После иммобилизации однодоменных антител на сенсорных поверхностях были протестированы чувствительные к простате-специфическому антигену человека (hPSA). Нанотела превзошли классические антитела в обнаружении клинически значимых концентраций чПСА. [42]
Чтобы увеличить вероятность кристаллизации целевой молекулы, нанотела можно использовать в качестве шаперонов кристаллизации . В качестве вспомогательных белков они могут уменьшать конформационную гетерогенность, связывая и стабилизируя только подмножество конформационных состояний. Они также могут маскировать поверхности, мешающие кристаллизации, одновременно расширяя области, образующие кристаллические контакты. [43] [37]
Терапевтический
[ редактировать ]Однодоменные антитела были протестированы как новый терапевтический инструмент против множества мишеней. У мышей, инфицированных вирусом гриппа А подтипа H5N1 , нанотела, направленные против гемагглютинина, подавляли репликацию вируса H5N1 in vivo и снижали заболеваемость и смертность. [44] Было показано, что нанотела, нацеленные на клеточный рецептор-связывающий домен факторов вирулентности токсина А и токсина B Clostridium difficile, нейтрализуют цитопатические эффекты в фибробластах in vitro . [45] Конъюгаты нанотел, распознающие антигенпрезентирующие клетки, успешно используются для обнаружения опухолей. [46] или целевая доставка антигена для создания сильного иммунного ответа. [47]
перорально доступные однодоменные антитела против диареи, вызванной E. coli, у поросят. Были разработаны и успешно протестированы [14] Другие заболевания желудочно-кишечного тракта , такие как воспалительные заболевания кишечника и рак толстой кишки , также являются возможными мишенями для перорально доступных однодоменных антител. [48]
Устойчивые к моющим средствам виды, нацеленные на поверхностный белок меха Malassezia, были разработаны для использования в шампунях против перхоти . [10]
В качестве подхода к фототермической терапии нанотела, связывающиеся с антигеном HER2 , который сверхэкспрессируется в клетках рака молочной железы и яичников, были конъюгированы с разветвленными наночастицами золота (см. рисунок). Опухолевые клетки уничтожали фототермически с помощью лазера в тестовой среде. [49]
Каплацизумаб , однодоменное антитело, нацеленное на фактор фон Виллебранда, проходит клинические испытания для профилактики тромбозов у пациентов с острым коронарным синдромом. [50] Исследование фазы II по изучению ALX-0081 при чрескожном коронарном вмешательстве высокого риска началось в сентябре 2009 года. [51]
Ablynx ожидает, что их нанотела смогут преодолевать гематоэнцефалический барьер и проникать в крупные солидные опухоли легче, чем цельные антитела, что позволит разработать лекарства против рака мозга . [48]
Недавно были идентифицированы нанотела, которые прочно связываются с доменом RBD спайкового белка бетакоронавирусов (включая SARS-CoV-2 , вызывающий COVID-19 ) и блокируют взаимодействие шипов с клеточным рецептором ACE2. [52] [18]
Сообщалось о применении различных однодоменных антител (нанотел) для профилактики и лечения заражения различными высокопатогенными коронавирусами человека (HPhCoV). Недавно были освещены перспективы, эффективность и проблемы использования нанотел для связывания и нейтрализации SARS-CoV-2 и ему подобных. [53]
одну из наиболее распространенных причин наганы – Trypanosoma brucei brucei ОдАт могут воздействовать на . Стийлеманс и др. В 2004 г. удалось индуцировать эффективные sdAb у кролика и Camelus dromedarius путем демонстрации вариабельного поверхностного гликопротеинового антигена иммунной системе позвоночных с помощью фага. В будущем эти методы лечения превзойдут естественные антитела, достигнув мест, которые в настоящее время недоступны из-за большего размера естественных антител. [54]
Ссылки
[ редактировать ]- ^ Jump up to: а б Хармсен М.М., Де Хаард Х.Дж. (ноябрь 2007 г.). «Свойства, производство и применение фрагментов однодоменных антител верблюдовых» . Прикладная микробиология и биотехнология . 77 (1): 13–22. дои : 10.1007/s00253-007-1142-2 . ПМК 2039825 . ПМИД 17704915 .
- ^ Английский Х, Хонг Дж, Хо М (январь 2020 г.). «Древние виды предлагают современные методы лечения: обновленная информация о акулы V NAR последовательностях однодоменных антител , фаговых библиотеках и потенциальных клинических применениях» . Терапия антителами . 3 (1): 1–9. дои : 10.1093/abt/tbaa001 . ПМК 7034638 . ПМИД 32118195 .
- ^ Мёллер А., Пион Э., Нараян В., Болл К.Л. (декабрь 2010 г.). «Внутриклеточная активация регуляторного фактора интерферона-1 нанотелами многофункционального (Mf1) домена» . Журнал биологической химии . 285 (49): 38348–38361. дои : 10.1074/jbc.M110.149476 . ПМЦ 2992268 . ПМИД 20817723 .
- ^ Ганнам А., Кумари С., Мюлдерманс С., Аббади А.К. (март 2015 г.). «Нанотела верблюжьих с высоким сродством к вирусу крапчатости бобов: возможный многообещающий инструмент для иммуномодулирования устойчивости растений к вирусам». Молекулярная биология растений . 87 (4–5): 355–369. дои : 10.1007/s11103-015-0282-5 . ПМИД 25648551 . S2CID 17578111 .
- ^ «Нанотела знаменуют новую эру в терапии рака» . Медицинские новости. 12 мая 2004 г.
- ^ «Трубопровод» . Аблинкс . Проверено 20 января 2010 г.
- ^ Гилл, Виктория (22 сентября 2021 г.). «Ковид: иммунная терапия с помощью лам обещает многообещающе» . Новости Би-би-си .
- ^ Jump up to: а б Баффингтон Дж., Дуан З., Квон Х.Дж., Хонг Дж., Ли Д., Фэн М. и др. (июнь 2023 г.). «Идентификация V NAR однодоменных антител акулы-няньки, нацеленных на субъединицу шипа S2 SARS-CoV-2» . Журнал ФАСЭБ . 37 (6): e22973. дои : 10.1096/fj.202202099RR . ПМЦ 10715488 . ПМИД 37191949 . S2CID 258717083 .
- ^ Хонг Дж., Квон Х.Дж., Качау Р., Чен Ч.З., Бутай К.Дж., Дуан З. и др. (май 2022 г.). «Нанотела одногорбого верблюда в целом нейтрализуют варианты SARS-CoV-2» . Труды Национальной академии наук Соединенных Штатов Америки . 119 (18): e2201433119. Бибкод : 2022PNAS..11901433H . дои : 10.1073/pnas.2201433119 . ПМК 9170159 . ПМИД 35476528 .
- ^ Jump up to: а б Долк Э., ван дер Ваарт М., Лутье Хулсик Д., Вриенд Г., де Хаард Х., Спинелли С. и др. (январь 2005 г.). «Выделение фрагментов антител ламы для профилактики перхоти путем фагового дисплея в шампуне» . Прикладная и экологическая микробиология . 71 (1): 442–450. Бибкод : 2005ApEnM..71..442D . дои : 10.1128/АЕМ.71.1.442-450.2005 . ПМК 544197 . ПМИД 15640220 .
- ^ Jump up to: а б Стэнфилд Р.Л., Дули Х., Флажник М.Ф., Уилсон И.А. (сентябрь 2004 г.). «Кристаллическая структура V-области однодоменного антитела акулы в комплексе с лизоцимом» . Наука . 305 (5691): 1770–1773. Бибкод : 2004Sci...305.1770S . дои : 10.1126/science.1101148 . ПМИД 15319492 . S2CID 25137728 .
- ^ ван дер Линден Р.Х., Френкен Л.Г., де Геус Б., Хармсен М.М., Руулс Р.К., Сток В. и др. (апрель 1999 г.). «Сравнение физико-химических свойств фрагментов антител VHH ламы и моноклональных антител мыши». Biochimica et Biophysical Acta (BBA) - Структура белка и молекулярная энзимология . 1431 (1): 37–46. дои : 10.1016/S0167-4838(99)00030-8 . ПМИД 10209277 .
- ^ Хармсен М.М., ван Солт CB, Хугендорн А., ван Зейдервельд Ф.Г., Ниволд Т.А., ван дер Мейлен Дж. (ноябрь 2005 г.). «Фрагменты однодоменных антител ламы, специфичные для фимбрии F4 Escherichia coli, эффективно ингибируют бактериальную адгезию in vitro, но плохо защищают от диареи». Ветеринарная микробиология . 111 (1–2): 89–98. дои : 10.1016/j.vetmic.2005.09.005 . ПМИД 16221532 .
- ^ Jump up to: а б Хармсен М.М., ван Солт CB, ван Зидервельд-ван Беммель А.М., Ниволд Т.А., ван Зийдервельд Ф.Г. (сентябрь 2006 г.). «Отбор и оптимизация протеолитически стабильных фрагментов однодоменных антител ламы для пероральной иммунотерапии». Прикладная микробиология и биотехнология . 72 (3): 544–551. дои : 10.1007/s00253-005-0300-7 . ПМИД 16450109 . S2CID 2319210 .
- ^ Jump up to: а б с д Хо М (июнь 2018 г.). «Первая редакционная статья: В поисках волшебных пуль» . Терапия антителами . 1 (1): 1–5. дои : 10.1093/abt/tby001 . ПМК 6086361 . ПМИД 30101214 .
- ^ Демитер А., Трансью Т.Р., Гаруди М.А., Ти М.Х., Портманс Ф., Хамерс Р. и др. (сентябрь 1996 г.). «Кристаллическая структура фрагмента однодоменного верблюжьего антитела VH в комплексе с лизоцимом». Структурная биология природы . 3 (9): 803–811. дои : 10.1038/nsb0996-803 . ПМИД 8784355 . S2CID 10419927 .
- ^ Jump up to: а б Фэн М., Бянь Х., Ву С., Фу Т., Фу Ю., Хун Дж. и др. (январь 2019 г.). «Создание и анализ секвенирования следующего поколения большой V NAR библиотеки однодоменных антител , отображаемой на фагах, от шести наивных акул-нянек» . Терапия антителами . 2 (1): 1–11. дои : 10.1093/abt/tby011 . ПМК 6312525 . ПМИД 30627698 .
- ^ Jump up to: а б с Хонг Дж., Квон Х.Дж., Качау Р., Чен Ч.З., Бутай К.Дж., Дуан З. и др. (май 2022 г.). «Нанотела одногорбого верблюда в целом нейтрализуют варианты SARS-CoV-2» . Труды Национальной академии наук Соединенных Штатов Америки . 119 (18): e2201433119. Бибкод : 2022PNAS..11901433H . дои : 10.1073/pnas.2201433119 . ПМК 9170159 . ПМИД 35476528 .
- ^ Jump up to: а б Ли Н., Цюань А., Ли Д., Пан Дж., Рен Х., Хельцель Г. и др. (апрель 2023 г.). «Шарнир IgG4 с трансмембранным доменом CD28 улучшает CAR T-клетки на основе V H H, нацеленные на мембранно-дистальный эпитоп GPC1 при раке поджелудочной железы» . Природные коммуникации . 14 (1): 1986. Бибкод : 2023NatCo..14.1986L . дои : 10.1038/s41467-023-37616-4 . ПМЦ 10082787 . ПМИД 37031249 .
- ^ Арбаби Гаруди М., Десмитер А., Винс Л., Хамерс Р., Мюлдерманс С. (сентябрь 1997 г.). «Отбор и идентификация фрагментов однодоменных антител из верблюжьих антител с тяжелой цепью» . Письма ФЭБС . 414 (3): 521–526. дои : 10.1016/S0014-5793(97)01062-4 . ПМИД 9323027 .
- ^ Ли Д., Инглиш Х., Хонг Дж., Лян Т., Мерлино Дж., Дэй CP, Хо М. (март 2022 г.). «Новая нацеленная на PD-L1 акулы V NAR, стратегия однодоменных CAR-T-клеток CAR-T, для лечения рака молочной железы и рака печени» . Молекулярная терапия онколитики . 24 : 849–863. дои : 10.1016/j.omto.2022.02.015 . ПМЦ 8917269 . ПМИД 35317524 .
- ^ Ли Д., Ван Р., Лян Т., Рен Х., Пак С., Тай Ч. и др. (сентябрь 2023 г.). «CAR-T-клетки B7-H3 CAR-T на основе верблюжьих нанотел демонстрируют высокую эффективность против крупных солидных опухолей» . Природные коммуникации . 14 (1): 5920. Бибкод : 2023NatCo..14.5920L . дои : 10.1038/s41467-023-41631-w . ПМЦ 10517151 . ПМИД 37739951 .
- ^ Чжан Ю.Ф., Сунь Ю., Хун Дж., Хо М. (январь 2023 г.). «Гуманизация однодоменного антитела Shark V NAR с использованием прививки CDR» . Текущие протоколы . 3 (1): е630. дои : 10.1002/cpz1.630 . ПМЦ 9813873 . ПМИД 36594750 .
- ^ Уорд Э.С., Гюссов Д., Гриффитс А.Д., Джонс П.Т., Уинтер Дж. (октябрь 1989 г.). «Связывающая активность репертуара отдельных вариабельных доменов иммуноглобулина, секретируемых Escherichia coli». Природа . 341 (6242): 544–546. Бибкод : 1989Natur.341..544W . дои : 10.1038/341544a0 . ПМИД 2677748 . S2CID 4329440 .
- ^ Jump up to: а б Фэн Р., Ван Р., Хонг Дж., Дауэр СМ, Круа Б.С., Хо М. (январь 2020 г.). «Выделение кроличьих однодоменных антител к B7-H3 посредством белковой иммунизации и фагового дисплея» . Терапия антителами . 3 (1): 10–17. дои : 10.1093/abt/tbaa002 . ПМЦ 7052794 . ПМИД 32166218 .
- ^ Jump up to: а б с Фэн М., Гао В., Ван Р., Чен В., Ман Ю.Г., Фигг В.Д. и др. (март 2013 г.). «Терапевтическое воздействие на глипикан-3 с помощью конформационно-специфического однодоменного антитела при гепатоцеллюлярной карциноме» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (12): E1083–E1091. Бибкод : 2013PNAS..110E1083F . дои : 10.1073/pnas.1217868110 . ПМК 3607002 . ПМИД 23471984 .
- ^ Холт Л.Дж., Херринг С., Джесперс Л.С., Вулвен Б.П., Томлинсон И.М. (ноябрь 2003 г.). «Доменные антитела: белки для терапии». Тенденции в биотехнологии . 21 (11): 484–490. дои : 10.1016/j.tibtech.2003.08.007 . ПМИД 14573361 .
- ^ Борребек К.А., Олин М. (декабрь 2002 г.). «Эволюция антител за пределами природы». Природная биотехнология . 20 (12): 1189–1190. дои : 10.1038/nbt1202-1189 . ПМИД 12454662 . S2CID 7937915 .
- ^ Jump up to: а б Тан З., Фэн М., Гао В., Фунг Ю., Чен В., Чаудхари А. и др. (апрель 2013 г.). «Человеческое однодоменное антитело проявляет мощную противоопухолевую активность, нацеливаясь на эпитоп мезотелина, расположенный вблизи поверхности раковой клетки» . Молекулярная терапия рака . 12 (4): 416–426. дои : 10.1158/1535-7163.MCT-12-0731 . ПМК 3624043 . ПМИД 23371858 .
- ^ Ли Н, Фу Х, Хьюитт С.М., Димитров Д.С., Хо М (август 2017 г.). «Терапевтическое воздействие на глипикан-2 через химерные антигенные рецепторы и иммунотоксины на основе однодоменных антител при нейробластоме» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (32): E6623–E6631. Бибкод : 2017PNAS..114E6623L . дои : 10.1073/pnas.1706055114 . ПМК 5559039 . ПМИД 28739923 .
- ^ Jump up to: а б Гао В., Тан З., Чжан Ю.Ф., Фэн М., Цянь М., Димитров Д.С., Хо М. (март 2015 г.). «Иммунотоксин, нацеленный на глипикан-3, регрессирует рак печени посредством двойного ингибирования передачи сигналов Wnt и синтеза белка» . Природные коммуникации . 6 : 6536. Бибкод : 2015NatCo...6.6536G . дои : 10.1038/ncomms7536 . ПМЦ 4357278 . ПМИД 25758784 .
- ^ Ван С., Гао В., Фэн М., Пастан И., Хо М. (май 2017 г.). «Создание иммунотоксина HN3-mPE24, нацеленного на глипикан-3, для терапии рака печени» . Онкотаргет . 8 (20): 32450–32460. дои : 10.18632/oncotarget.10592 . ПМК 5464801 . ПМИД 27419635 .
- ^ Флеминг Б.Д., Urban DJ, Hall MD, Longerich T, Greten TF, Pastan I, Ho M (май 2020 г.). «Инженерный иммунотоксин против GPC3, HN3-ABD-T20, вызывает регрессию ксенотрансплантатов рака печени мышей за счет длительного удержания сыворотки» . Гепатология . 71 (5): 1696–1711. дои : 10.1002/hep.30949 . ПМК 7069773 . ПМИД 31520528 .
- ^ Коллури А., Ли Д., Ли Н., Дуан З., Робертс Л.Р., Хо М. (февраль 2023 г.). «Т-клетки химерного антигенного рецептора человека на основе VH, нацеленные на глипикан 3, устраняют опухоли в доклинических моделях ГЦК» . Гепатологические сообщения . 7 (2): e0022. дои : 10.1097/HC9.0000000000000022 . ПМЦ 9851680 . ПМИД 36691969 .
- ^ Ли Н, Вэй Л, Лю Х, Бай Х, Йе Ю, Ли Д и др. (октябрь 2019 г.). «Вьющийся, богатый цистеином домен в глипикане-3 опосредует связывание Wnt и регулирует рост опухоли гепатоцеллюлярной карциномы у мышей» . Гепатология . 70 (4): 1231–1245. дои : 10.1002/hep.30646 . ПМК 6783318 . ПМИД 30963603 .
- ^ Мюлдерманс С (2013). «Нанотела: природные однодоменные антитела». Ежегодный обзор биохимии . 82 : 775–797. doi : 10.1146/annurev-biochem-063011-092449 . ПМИД 23495938 .
- ^ Jump up to: а б Гулати С., Джин Х., Масухо И., Орбан Т., Цай Ю., Пардон Э. и др. (май 2018 г.). «Нацеливание на передачу сигналов рецептора, связанного с G-белком, на уровне G-белка с помощью селективного ингибитора нанотел» . Природные коммуникации . 9 (1): 1996. Бибкод : 2018NatCo...9.1996G . дои : 10.1038/s41467-018-04432-0 . ПМК 5959942 . ПМИД 29777099 .
- ^ Ротбауэр У., Золгадр К., Тиллиб С., Новак Д., Шермелле Л., Гал А. и др. (ноябрь 2006 г.). «Нацеливание и отслеживание антигенов в живых клетках с помощью флуоресцентных нанотел». Природные методы . 3 (11): 887–889. дои : 10.1038/nmeth953 . ПМИД 17060912 . S2CID 29985078 .
- ^ Ротбауэр У., Золгадр К., Мюлдерманс С., Шеперс А., Кардозо М.С., Леонхардт Х. (февраль 2008 г.). «Универсальная наноловушка для биохимических и функциональных исследований с флуоресцентными слитыми белками» . Молекулярная и клеточная протеомика . 7 (2): 282–289. дои : 10.1074/mcp.M700342-MCP200 . ПМИД 17951627 .
- ^ Райс Дж., Каплан С., Платонова Е., Эглиди Х., Эверс Х. (июнь 2012 г.). «Простой и универсальный метод микроскопии сверхвысокого разрешения на основе GFP с помощью нанотел». Природные методы . 9 (6): 582–584. doi : 10.1038/nmeth.1991 . ПМИД 22543348 . S2CID 12190938 .
- ^ Фанг Т., Лу Х, Бергер Д., Гмайнер С., Чо Дж., Шалек Р. и др. (декабрь 2018 г.). «Иммуноокрашивание нанотел для коррелированной световой и электронной микроскопии с сохранением ультраструктуры» . Природные методы . 15 (12): 1029–1032. дои : 10.1038/s41592-018-0177-x . ПМК 6405223 . ПМИД 30397326 .
- ^ Саеренс Д., Фредерикс Ф., Рикманс Г., Конрат К., Янс К., Брис Л. и др. (декабрь 2005 г.). «Разработка верблюжьих однодоменных антител и химия иммобилизации для определения специфического антигена простаты человека». Аналитическая химия . 77 (23): 7547–7555. дои : 10.1021/ac051092j . ПМИД 16316161 .
- ^ Койде С. (август 2009 г.). «Инженерия шаперонов рекомбинантной кристаллизации» . Современное мнение в области структурной биологии . 19 (4): 449–457. дои : 10.1016/j.sbi.2009.04.008 . ПМЦ 2736338 . ПМИД 19477632 .
- ^ Ибаньес Л.И., Де Филетт М., Хультберг А., Веррипс Т., Темпертон Н., Вайс Р.А. и др. (апрель 2011 г.). «Нанотела с нейтрализующей активностью in vitro защищают мышей от заражения вирусом гриппа H5N1» . Журнал инфекционных болезней . 203 (8): 1063–1072. дои : 10.1093/infdis/jiq168 . hdl : 1854/LU-1212323 . ПМИД 21450996 .
- ^ Хассак Дж., Арбаби-Гахруди М., ван Фаассен Х., Сонгер Дж.Г., Нг К.К., Маккензи Р., Танха Дж. (март 2011 г.). «Нейтрализация токсина А Clostridium difficile с помощью однодоменных антител, нацеленных на домен, связывающий клеточный рецептор» . Журнал биологической химии . 286 (11): 8961–8976. дои : 10.1074/jbc.M110.198754 . ПМК 3058971 . ПМИД 21216961 .
- ^ Фанг Т., Дуарте Дж. Н., Линг Дж., Ли З., Гузман Дж. С., Плу Х. Л. (февраль 2016 г.). «Структурно определенные конъюгаты αMHC-II нанотело-лекарственное средство: терапевтическая система и система визуализации для B-клеточной лимфомы» . Ангеванде Хеми . 55 (7): 2416–2420. дои : 10.1002/anie.201509432 . ПМЦ 4820396 . ПМИД 26840214 .
- ^ Фанг Т., Ван Элссен СН, Дуарте Й.Н., Гузман Дж.С., Чахал Дж.С., Линг Дж., Плух Х.Л. (август 2017 г.). «Направленная доставка антигена с помощью антигена MHC VHH класса II вызывает направленный иммунитет к αMUC1 (Tn)» . Химическая наука . 8 (8): 5591–5597. дои : 10.1039/c7sc00446j . ПМЦ 5618788 . ПМИД 28970938 .
- ^ Jump up to: а б «Нанотела» . Nanobody.org. 2006.
- ^ Ван де Брук Б., Девогдт Н., Д'Холландер А., Гийс Х.Л., Янс К., Лага Л. и др. (июнь 2011 г.). «Специфическое нацеливание на клетки с помощью конъюгированных с нанотелами разветвленных наночастиц золота для фототермической терапии». АСУ Нано . 5 (6): 4319–4328. дои : 10.1021/nn1023363 . ПМИД 21609027 .
- ^ «Ablynx объявляет промежуточные результаты первого исследования фазы I NANOBODY® ALX-0081 (ANTI-VWF)» . Био-Медицина.org. 2 июля 2007 г.
- ^ Номер клинического исследования NCT01020383 «Сравнительное исследование ALX-0081 по сравнению с ингибитором GPIIb/IIIa у пациентов с высоким риском чрескожного коронарного вмешательства (ЧКВ)» на сайте ClinicalTrials.gov.
- ^ Wrapp D, De Vlieger D, Corbett KS, Torres GM, Wang N, Van Breedam W и др. (май 2020 г.). «Структурная основа мощной нейтрализации бетакоронавирусов однодоменными антителами верблюдовых» . Клетка . 181 (5): 1004–1015.e15. дои : 10.1016/j.cell.2020.04.031 . ПМЦ 7199733 . ПМИД 32375025 .
- ^ Конвар Р. (23 июня 2020 г.). «Нанотела: перспективы расширения спектра нейтрализующих антител против нового коронавируса SARS-CoV-2» . Границы в иммунологии . 11 : 1531. дои : 10.3389/fimmu.2020.01531 . ПМЦ 7324746 . PMID 32655584 .
- ^ Мюлдерманс С (2 июня 2013 г.). «Нанотела: природные однодоменные антитела». Ежегодный обзор биохимии . 82 (1). Годовые обзоры : 775–797. doi : 10.1146/annurev-biochem-063011-092449 . ПМИД 23495938 . п. 788:
Более того...
Внешние ссылки
[ редактировать ]Сконструированные моноклональные антитела и миметики антител | ||
|---|---|---|
| Цельное антитело |  | |
| Потрясающий фрагмент | ||
| Переменный фрагмент | ||
| Меньшие единицы | ||
| Внутриклеточный | ||
| Миметики антител | ||