Ксилема

Ксилема — один из двух типов транспортной ткани ( сосудистых растений второй — флоэма) . Основная функция ксилемы — транспортировка воды от корней к стеблям и листьям, а также транспортировка питательных веществ . [1] [2] Слово ксилема происходит от древнегреческого слова ξύλον ( ксилон ), что означает «дерево»; Самая известная ткань ксилемы — древесина , хотя она встречается по всему растению. [3] Этот термин был введен Карлом Нэгели в 1858 году. [4] [5]
Структура
[ редактировать ]
Наиболее характерными клетками ксилемы являются длинные трахеарные элементы, переносящие воду. Трахеиды и элементы сосудов отличаются формой; Элементы сосудов короче и соединены между собой в длинные трубки, называемые сосудами . [6]
Ксилема также содержит два других типа клеток: паренхиму и волокна . [7]
Ксилему можно найти:
- в сосудистых пучках , присутствует у недревесных растений и недревесных частей древесных растений
- во вторичной ксилеме, заложенной меристемой, называемой сосудистым камбием у древесных растений
- в составе звездчатого расположения, не разделенного на пучки, как у многих папоротников .
На переходных стадиях растений со вторичным ростом первые две категории не исключают друг друга, хотя обычно проводящий пучок содержит только первичную ксилему .
Характер ветвления ксилемы соответствует закону Мюррея . [8]
Первичная и вторичная ксилема
[ редактировать ]Первичная ксилема образуется при первичном росте из прокамбия . Он включает протоксилему и метаксилему. Метаксилема развивается после протоксилемы, но раньше вторичной ксилемы. Метаксилема имеет более широкие сосуды и трахеиды, чем протоксилема.
Вторичная ксилема образуется при вторичном росте из сосудистого камбия . Хотя вторичная ксилема также встречается у представителей голосеменных групп Gnetophyta и Ginkgophyta и в меньшей степени у представителей Cycadophyta , двумя основными группами, в которых может быть обнаружена вторичная ксилема, являются:
- хвойные ( Coniferae ): известно около 600 видов хвойных. [9] У всех видов имеется вторичная ксилема, структура которой относительно однородна во всей группе. Многие хвойные деревья становятся высокими деревьями: вторичная ксилема таких деревьев используется и продается как хвойная древесина .
- покрытосеменные ( Angiospermae ): насчитывается около 250 000. [9] известные виды покрытосеменных растений. встречается редко Внутри этой группы вторичная ксилема у однодольных . [10] Многие неоднодольные покрытосеменные растения становятся деревьями, а их вторичная ксилема используется и продается как лиственная древесина .
Основная функция – восходящий водный транспорт.
[ редактировать ]Ксилема, сосуды и трахеиды корней, стеблей и листьев соединены между собой, образуя непрерывную систему водопроводящих каналов, идущих ко всем частям растения. Система транспортирует воду и растворимые минеральные питательные вещества от корней по всему растению. Он также используется для замены воды, теряемой при транспирации и фотосинтезе. ксилемы Сок состоит в основном из воды и неорганических ионов, хотя он также может содержать ряд органических химических веществ. Транспорт пассивен, не питается за счет энергии, затрачиваемой самими элементами трахеи , мертвыми к моменту созревания и уже не имеющими живого содержимого. Транспортировка сока вверх становится более трудной по мере увеличения высоты растения, и считается, что перенос воды вверх по ксилеме ограничивает максимальную высоту деревьев. [11] Три явления вызывают выделение сока ксилемы:
- Гипотеза потока под давлением : сахара, образующиеся в листьях и других зеленых тканях, удерживаются в системе флоэмы, создавая перепад давления растворенных веществ по сравнению с системой ксилемы, несущей гораздо меньшую нагрузку растворенных веществ - воды и минералов. Давление во флоэме может достигать нескольких МПа. [12] намного выше атмосферного давления. Избирательная взаимосвязь между этими системами позволяет высокой концентрации растворенных веществ во флоэме вытягивать ксилемную жидкость вверх под действием отрицательного давления.
- Транспирационная тяга . Точно так же испарение воды в атмосферу также с поверхности клеток мезофилла создает отрицательное давление в верхней части растения. Это приводит к образованию миллионов менисков в клеточной стенке мезофилла. Возникающее в результате поверхностное натяжение вызывает отрицательное давление или напряжение в ксилеме, которая вытягивает воду из корней и почвы.
- Корневое давление : если водный потенциал клеток корня более отрицательный, чем потенциал почвы , обычно из-за высоких концентраций растворенных веществ , вода может перемещаться путем осмоса в корень из почвы . Это вызывает положительное давление, которое заставляет ксилему подтягиваться к листьям. В некоторых случаях сок будет вытесняться из листа через гидатод в результате явления, известного как гуттация . Корневое давление является самым высоким утром перед открытием устьиц и позволяет начать транспирацию. Разные виды растений могут иметь разное корневое давление даже в одинаковой среде; примеры включают до 145 кПа у Vitis riparia и около нуля у Celastrus orbiculatus . [13]
Основной силой, создающей капиллярное движение воды вверх в растениях, является сцепление воды с поверхностью ксилемных каналов. [14] [15] Капиллярное действие обеспечивает силу, которая устанавливает равновесную конфигурацию, уравновешивая гравитацию. Когда транспирация удаляет воду наверху, поток необходим для возвращения к равновесию.
Транспирационная тяга возникает в результате испарения воды с клеток листьев поверхности . Это испарение заставляет поверхность воды погружаться в поры клеточной стенки . Под действием капиллярных сил вода образует вогнутые мениски внутри пор. Высокое поверхностное натяжение воды вытягивает вогнутость наружу, создавая достаточную силу , чтобы поднять воду на высоту до ста метров от уровня земли до самых высоких ветвей дерева .
Транспирационная тяга требует, чтобы сосуды, транспортирующие воду, были очень маленького диаметра; в противном случае кавитация разрушит толщу воды. По мере того, как вода испаряется из листьев, через растение всасывается еще больше воды, чтобы заменить ее. Когда давление воды внутри ксилемы достигает экстремальных значений из-за малого поступления воды из корней (если, например, почва сухая), то газы выходят из раствора и образуют пузырь – образуется эмболия , которая быстро распространяется. к другим соседним клеткам, если только нет окаймленных ямок (они имеют пробкообразную структуру, называемую тором, которая закрывает отверстие между соседними клетками и останавливает распространение эмболии). Даже после того, как произошла эмболия, растения способны пополнить ксилему и восстановить ее функциональность. [16]
Теория сцепления-напряжения
[ редактировать ]Теория сцепления-напряжения — это теория межмолекулярного притяжения , объясняющая процесс движения воды вверх (против силы тяжести ) через ксилему растений . Его предложили в 1894 году Джон Джоли и Генри Горацио Диксон . [17] [18] Несмотря на многочисленные возражения, [19] [20] это наиболее широко распространенная теория транспортировки воды через сосудистую систему растения, основанная на классических исследованиях Диксона-Жоли (1894 г.), Ойгена Аскенаси (1845–1903) (1895 г.), [21] [22] и Диксон (1914,1924). [23] [24]
Вода – полярная молекула . Когда две молекулы воды приближаются друг к другу, слегка отрицательно заряженный атом кислорода одной образует водородную связь со слегка положительно заряженным атомом водорода другой. Эта сила притяжения, наряду с другими межмолекулярными силами , является одним из основных факторов, ответственных за возникновение поверхностного натяжения в жидкой воде. Это также позволяет растениям переносить воду от корня через ксилему к листу.
Вода постоянно теряется за счет транспирации листа. Когда одна молекула воды теряется, другая притягивается за собой в результате процессов сцепления и напряжения. Транспирационная тяга, использующая капиллярное действие и присущее воде поверхностное натяжение, является основным механизмом движения воды в растениях. Однако это не единственный задействованный механизм. Любое использование воды в листьях заставляет воду проникать в них.
Транспирация в листьях создает напряжение (перепад давления) в клеточных стенках клеток мезофилла . Из-за этого напряжения вода вытягивается из корней в листья, чему способствует сцепление (притяжение между отдельными молекулами воды из-за водородных связей ) и адгезия (липкость между молекулами воды и гидрофильными клеточными стенками растений). Этот механизм потока воды работает благодаря водному потенциалу (вода течет от высокого потенциала к низкому) и правилам простой диффузии . [25]
За последнее столетие было проведено множество исследований механизма транспорта сока ксилемы; сегодня большинство ученых-растениеведов продолжают соглашаться с тем, что теория сцепления-напряжения лучше всего объясняет этот процесс, но были предложены теории мультисил, которые выдвигают гипотезу о нескольких альтернативных механизмах, включая продольные осмотического давления градиенты в клетках и ксилеме , осевые градиенты потенциала в сосудах и гелеобразование. и межфазные градиенты, поддерживаемые газовыми пузырьками. [26] [27]
Измерение давления
[ редактировать ]
До недавнего времени дифференциальное давление (всасывание) транспирационного притяжения можно было измерить только косвенно, применяя внешнее давление с помощью бомбы давления для противодействия ему. [28] Когда была разработана технология проведения прямых измерений с помощью зонда давления, поначалу существовали некоторые сомнения в правильности классической теории, поскольку некоторые рабочие не могли продемонстрировать отрицательное давление. Более поздние измерения по большей части подтверждают классическую теорию. Транспорт ксилемы осуществляется за счет комбинации [29] транспирационного притяжения сверху и корневого давления снизу, что усложняет интерпретацию измерений.
Эволюция
[ редактировать ]Ксилема появилась в начале истории наземной растительной жизни. Ископаемые растения с анатомически сохранившейся ксилемой известны с силура (более 400 млн лет назад), а следовые окаменелости, напоминающие отдельные клетки ксилемы, могут быть обнаружены в более ранних ордовикских породах. [ нужна ссылка ] Самая ранняя истинная и узнаваемая ксилема состоит из трахеид со спирально-кольцевым армирующим слоем, добавленным к клеточной стенке . Это единственный тип ксилемы, обнаруженный у самых ранних сосудистых растений, и этот тип клеток продолжает обнаруживаться в протоксилеме (первообразовавшейся ксилеме) всех живых групп сосудистых растений. Несколько групп растений позже независимо развили ямчатые клетки трахеиды в результате конвергентной эволюции . У живых растений ямчатые трахеиды не появляются в развитии до созревания метаксилемы ( вслед за протоксилемой ).
У большинства растений ямчатые трахеиды функционируют как первичные транспортные клетки. Другой тип сосудистых элементов, обнаруженный у покрытосеменных, — это сосудистый элемент . Элементы сосудов соединяются встык, образуя сосуды, в которых вода течет беспрепятственно, как в трубе. Наличие сосудов ксилемы (также называемой трахеей [30] ) считается одной из ключевых инноваций, приведших к успеху покрытосеменных растений . [31] Однако появление сосудистых элементов не ограничивается покрытосеменными, и они отсутствуют в некоторых архаичных или «базальных» линиях покрытосеменных: (например, Amborellaceae , Tetracentraceae , Trochodendraceae и Winteraceae ), а их вторичная ксилема описана Артуром. Кронквист как «примитивно бессосудистый». Кронквист считал, что сосуды Gnetum сходятся с сосудами покрытосеменных. [32] Является ли отсутствие сосудов у базальных покрытосеменных примитивным состоянием, оспаривается. Альтернативная гипотеза утверждает, что элементы сосудов возникли у предшественников покрытосеменных и впоследствии были утеряны.

Для фотосинтеза растения должны поглощать CO2 из атмосферы. Однако за это приходится платить: пока устьица открыты для проникновения CO 2 , вода может испаряться. [33] Вода теряется гораздо быстрее, чем поглощается CO2 , поэтому растениям необходимо ее восполнять, и у них разработаны системы транспортировки воды из влажной почвы к месту фотосинтеза. [33] Ранние растения всасывали воду между стенками своих клеток, а затем развили способность контролировать потерю воды (и поглощение CO 2 ) с помощью устьиц. Специализированные ткани, транспортирующие воду, вскоре развились в форме гидроидов, трахеид, затем вторичной ксилемы, затем эндодермы и, наконец, сосудов. [33]
Высокие уровни CO2 в силурийско-девонском периоде, когда растения впервые колонизировали сушу, означали, что потребность в воде была относительно низкой. Поскольку CO 2 извлекался из атмосферы растениями, при его улавливании терялось больше воды, и появились более элегантные механизмы транспортировки. [33] По мере развития механизмов переноса воды и водонепроницаемой кутикулы растения смогли выжить, не будучи постоянно покрыты пленкой воды. Этот переход от пойкилогидрии к гомойогидрии открыл новый потенциал для колонизации. [33] Тогда растениям требовалась прочная внутренняя структура, которая содержала бы длинные узкие каналы для транспортировки воды из почвы ко всем различным частям надпочвенного растения, особенно к тем частям, где происходил фотосинтез.
В силурийском периоде CO 2 был легко доступен, поэтому для его приобретения требовалось мало воды. К концу каменноугольного периода, когда уровень CO 2 снизился до уровня, приближающегося к сегодняшнему, на единицу поглощения CO 2 было потеряно примерно в 17 раз больше воды . [33] Однако даже в те «легкие» первые дни вода была в дефиците, и ее приходилось доставлять к частям растения из влажной почвы, чтобы избежать высыхания . Этот ранний водный транспорт воспользовался преимуществами механизма сцепления-напряжения, присущего воде. Вода имеет тенденцию диффундировать в более сухие области, и этот процесс ускоряется, когда вода может проходить по ткани с небольшим пространством. В небольших проходах, например, между стенками растительных клеток (или в трахеидах), столб воды ведет себя как резина — когда молекулы испаряются с одного конца, они тянут молекулы за собой по каналам. Таким образом, только транспирация была движущей силой водного транспорта у ранних растений. [33] Однако без специальных транспортных сосудов механизм сцепления-напряжения не может транспортировать воду более чем на 2 см, что серьезно ограничивает размер самых ранних растений. [33] Этот процесс требует постоянной подачи воды с одного конца для поддержания цепей; Чтобы избежать его истощения, у растений появилась водонепроницаемая кутикула . Ранняя кутикула могла не иметь пор, но не покрывала всю поверхность растения, поэтому газообмен мог продолжаться. [33] Однако порой обезвоживание было неизбежным; Ранние растения справляются с этим, сохраняя большое количество воды между стенками своих клеток, а когда дело доходит до этого, они переживают трудные времена, «приостанавливая» жизнь до тех пор, пока не будет получено больше воды. [33]

Чтобы освободиться от ограничений небольшого размера и постоянной влажности, которые накладывала паренхиматозная транспортная система, растениям требовалась более эффективная система водного транспорта. В раннем силуре у них появились специализированные клетки, которые были одревесневшими (или содержали аналогичные химические соединения). [33] избежать взрыва; этот процесс совпал со смертью клеток, позволив опорожнить их внутренности и пройти через них воду. [33] Эти более широкие, мертвые, пустые клетки имели в миллион раз большую проводимость, чем межклеточный метод, что давало возможность переноса на большие расстояния и более высокую CO 2 скорость диффузии .
Самыми ранними макрофоссилиями, имеющими трубки для транспортировки воды, являются силурийские растения, относящиеся к роду Cooksonia . [34] Раннедевонские претрахеофиты Aglaophyton и Horneophyton имеют строение, очень похожее на гидроиды современных мхов.Растения продолжали изобретать новые способы снижения сопротивления потоку внутри своих клеток, тем самым повышая эффективность транспорта воды. Полосы на стенках трубок, наблюдаемые фактически начиная с раннего силура, [35] представляют собой раннюю импровизацию, способствующую легкому течению воды. [36] Ленточные трубки, а также трубки с ямчатым орнаментом на стенках одревеснели. [37] и, когда они образуют одноклеточные кондуиты, считаются трахеидами . Это «следующее поколение» конструкции транспортных ячеек, имеющее более жесткую структуру, чем гидроиды, что позволяет им выдерживать более высокие уровни давления воды. [33] Трахеиды могут иметь единое эволюционное происхождение, возможно, внутри роголистников. [38] объединяющий все трахеофиты (но они могли эволюционировать более одного раза). [33]
Водный транспорт требует регулирования, а динамический контроль обеспечивают устьица . [39] Регулируя объем газообмена, они могут ограничить количество воды, теряемой в результате транспирации. Это важная роль, когда водоснабжение непостоянно, и действительно, устьица, по-видимому, развились раньше трахеид и присутствуют у несосудистых роголистников. [33]
Эндодерма , вероятно, возникла в силуу-девоне, но первые ископаемые свидетельства такой структуры относятся к каменноугольному периоду. [33] Эта структура в корнях покрывает ткань, транспортирующую воду, и регулирует ионный обмен (и предотвращает попадание нежелательных патогенов и т. д. в систему транспорта воды). Энтодерма также может оказывать давление вверх, вытесняя воду из корней, когда транспирация недостаточна.
Как только растения развили этот уровень контролируемого транспорта воды, они стали по-настоящему гомойогидридными, способными извлекать воду из окружающей среды через корнеподобные органы, а не полагаться на пленку поверхностной влаги, что позволило им вырасти до гораздо больших размеров. [33] В результате своей независимости от окружающей среды они утратили способность пережить высыхание, а сохранение этой черты обходится дорого. [33]
В девоне максимальный диаметр ксилемы со временем увеличивался, а минимальный оставался практически постоянным. [36] К среднему девону диаметр трахеид некоторых линий растений ( зостерофиллофитов ) выровнялся. [36] Более широкие трахеиды позволяют воде транспортироваться быстрее, но общая скорость транспортировки зависит также от общей площади поперечного сечения самого пучка ксилемы. [36] Увеличение толщины сосудистых пучков, по-видимому, также коррелирует с шириной осей растения и высотой растения; это также тесно связано с внешним видом листьев [36] и повышенная плотность устьиц, что приведет к увеличению потребности в воде. [33]
Хотя более широкие трахеиды с прочными стенками позволяют добиться более высокого напряжения транспортировки воды, это увеличивает вероятность кавитации. [33] Кавитация возникает, когда внутри сосуда образуется пузырь воздуха, разрывающий связи между цепочками молекул воды и не позволяющий им вытягивать больше воды вверх за счет своего когезионного напряжения. Трахеиду, однажды попавшую в полость, невозможно удалить из-за эмболии и вернуть в строй (за исключением некоторых продвинутых покрытосеменных растений). [40] [41] которые разработали соответствующий механизм). Поэтому растениям стоит избегать возникновения кавитации. По этой причине ямки в стенках трахеиды имеют очень маленький диаметр, чтобы предотвратить попадание воздуха и образование пузырьков. Циклы замораживания-оттаивания являются основной причиной кавитации. Повреждение стенки трахеиды почти неизбежно приводит к утечке воздуха и кавитации, отсюда важность параллельной работы многих трахеид. [33]
После возникновения кавитации у растений появляется ряд механизмов, позволяющих сдержать повреждение. [33] Небольшие ямки соединяют соседние каналы, позволяя жидкости течь между ними, но не воздуху, хотя эти ямки, предотвращающие распространение эмболии, также являются ее основной причиной. [33] Эти изрытые поверхности еще больше уменьшают поток воды через ксилему на целых 30%. [33] Разнообразие форм нитей ксилемы с топологиями трахеидной сети, все более устойчивыми к распространению эмболии, вероятно, способствовало увеличению размеров растений и колонизации более засушливых местообитаний во время девонской радиации . [42] У хвойных к юре развиты окаймленные ямки, имевшие клапанообразные структуры, изолирующие кавитационные элементы. Эти структуры тора-марго имеют непроницаемый диск (тор), подвешенный на проницаемой мембране (марго) между двумя соседними порами. Когда трахеида на одной стороне разгерметизируется, диск всасывается в пору на этой стороне и блокирует дальнейший поток. [33] Другие растения просто переносят кавитацию. Например, в начале каждой весны на дубах вырастает кольцо широких сосудов, ни один из которых не выдерживает зимних морозов. [ нужна ссылка ] Каждую весну клены используют корневое давление, чтобы вытеснить сок вверх из корней, выдавливая пузырьки воздуха. [ нужна ссылка ]
Для роста в высоту использовалась еще одна особенность трахеид — поддержка, обеспечиваемая их одревесневшими стенками. Несуществующие трахеиды сохранились и образовали прочный древесный стебель, в большинстве случаев образованный вторичной ксилемой. Однако у ранних растений трахеиды были слишком механически уязвимы и сохраняли центральное положение со слоем жесткой склеренхимы на внешнем крае стеблей. [33] Даже когда трахеиды играют структурную роль, они поддерживаются склеренхиматической тканью.
Трахеиды заканчиваются стенками, которые оказывают большое сопротивление потоку; [36] Члены резервуара имеют перфорированные торцевые стенки и расположены последовательно, чтобы работать так, как если бы они были одним непрерывным резервуаром. [36] Функция торцевых стенок, которые были обычным состоянием в девоне, вероятно, заключалась в предотвращении эмболий . Эмболия – это образование воздушного пузыря в трахеиде. Это может произойти в результате замерзания или растворения газов из раствора. Если образовалась эмболия, ее обычно невозможно удалить (см. ниже); пораженная клетка не может вытягивать воду и становится бесполезной.
Если исключить торцевые стенки, трахеиды предсосудистых растений могли работать при той же гидравлической проводимости, что и трахеиды первого сосудистого растения Cooksonia . [36]
Размер трахеид ограничен, поскольку они состоят из одной клетки; это ограничивает их длину, что, в свою очередь, ограничивает их максимальный полезный диаметр до 80 мкм. [33] Проводимость растет с увеличением диаметра в четвертой степени, поэтому увеличение диаметра приносит огромную пользу; элементы сосудов , состоящие из ряда ячеек, соединенных на концах, преодолели этот предел и позволили образоваться более крупным трубкам, достигающим диаметра до 500 мкм и длины до 10 м. [33]
Сосуды впервые появились в засушливые периоды с низким содержанием CO 2 в поздней перми независимо у хвощей, папоротников и Selaginellales , а позднее появились в середине мела у покрытосеменных и гнетофитов. [33] Суда позволяют одной и той же площади поперечного сечения древесины переносить примерно в сто раз больше воды, чем трахеиды! [33] Это позволило растениям заполнить большую часть своих стеблей структурными волокнами, а также открыло новую нишу для виноградных лоз , которые могли переносить воду, не будучи такими толстыми, как дерево, на котором они росли. [33] Несмотря на эти преимущества, древесина на основе трахеид намного легче и, следовательно, дешевле в производстве, поскольку сосуды необходимо гораздо более армировать, чтобы избежать кавитации. [33]
Разработка
[ редактировать ]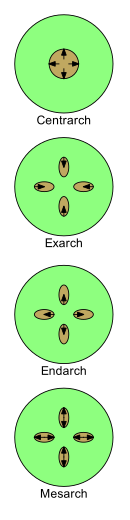
Развитие ксилемы можно описать четырьмя терминами: центрарх, экзарх, эндарх и мезарх . По мере развития у молодых растений его природа меняется от протоксилемы к метаксилеме (т. е. от первой ксилемы к послексилеме ). Порядок расположения протоксилемы и метаксилемы важен при изучении морфологии растений.
Протоксилема и метаксилема
[ редактировать ]По мере роста молодого сосудистого растения в его стеблях и корнях образуются одна или несколько нитей первичной ксилемы. Первая развивающаяся ксилема называется «протоксилемой». По внешнему виду протоксилема обычно отличается более узкими сосудами, образованными более мелкими клетками. Некоторые из этих клеток имеют стенки, содержащие утолщения в виде колец или спиралей. Функционально протоксилема может расширяться: клетки способны увеличиваться в размерах и развиваться, пока стебель или корень удлиняются. Позже в нитях ксилемы развивается «метаксилема». Сосуды и клетки метаксилемы обычно крупнее; клетки имеют утолщения, которые обычно имеют форму лестничных поперечных полосок (лестничные) или сплошных листов, за исключением отверстий или ямок (ямчатые). Функционально метаксилема завершает свое развитие после прекращения удлинения, когда клеткам больше не нужно увеличиваться в размерах. [43] [44]
Образцы протоксилемы и метаксилемы
[ редактировать ]Существует четыре основных закономерности расположения протоксилемы и метаксилемы в стеблях и корнях.
- Центрарх относится к случаю, когда первичная ксилема образует один цилиндр в центре стебля и развивается от центра наружу. Таким образом, протоксилема находится в центральном ядре, а метаксилема - в цилиндре вокруг него. [45] Эта закономерность была распространена у ранних наземных растений, таких как риниофиты , но не присутствует ни у каких живых растений. [ нужна ссылка ]
Остальные три термина используются там, где имеется более одной цепи первичной ксилемы.
- Экзарх используется, когда в стебле или корне имеется более одной нити первичной ксилемы и ксилема развивается снаружи внутрь к центру, т. е. центростремительно. Таким образом, метаксилема расположена ближе всего к центру стебля или корня, а протоксилема — ближе всего к периферии. Обычно считается, что корни сосудистых растений имеют экзарховое развитие. [43]
- Эндарч используется, когда в стебле или корне имеется более одной нити первичной ксилемы и ксилема развивается изнутри наружу к периферии, т. е. центробежно. Таким образом, протоксилема находится ближе всего к центру стебля или корня, а метаксилема — ближе всего к периферии. Стебли семенных растений обычно имеют эндархическое развитие. [43]
- Мезарх используется, когда в стебле или корне имеется более одной нити первичной ксилемы и ксилема развивается из середины нити в обоих направлениях. Таким образом, метаксилема находится как на периферической, так и на центральной сторонах цепи, а протоксилема находится между метаксилемой (возможно, окруженной ею). Листья и стебли многих папоротников имеют мезарное развитие. [43]
История
[ редактировать ]В своей книге De plantis libri XVI (О растениях, в 16 книгах) (1583) итальянский врач и ботаник Андреа Чезальпино предположил, что растения вытягивают воду из почвы не за счет магнетизма ( ut magnes Ferrum trahit , поскольку магнитное железо притягивает), ни за счет всасывания. ( вакуум ), а путем впитывания, как это происходит в случае с бельем, губками или порошками. [46] Итальянский биолог Марчелло Мальпиги был первым, кто описал и проиллюстрировал сосуды ксилемы, что он сделал в своей книге Anatome plantarum ... (1675). [47] [примечание 1] Хотя Мальпиги считал, что ксилема содержит только воздух, британский врач и ботаник Неемия Грю , который был современником Мальпиги, считал, что сок поднимается как через кору, так и через ксилему. [48] Однако, по мнению Грю, капиллярное действие в ксилеме поднимет сок всего на несколько дюймов; Чтобы поднять сок на верхушку дерева, Грю предположил, что паренхимные клетки становятся набухшими и тем самым не только выдавливают сок в трахеидах, но и вытесняют некоторое количество сока из паренхимы в трахеиды. [49] В 1727 году английский священник и ботаник Стивен Хейлз показал, что транспирация листьев растения заставляет воду проходить через его ксилему. [50] [примечание 2] К 1891 году польско-немецкий ботаник Эдуард Страсбургер показал, что для транспортировки воды в растениях не требуется, чтобы клетки ксилемы были живыми. [51]
См. также
[ редактировать ]- Флоэма
- Континуум атмосферы почвенных растений
- Звезды
- Всасывание
- Тилоз
- Сосудистый пучок
- Сосудистая ткань
- Ксилемный сок
Пояснительные примечания
[ редактировать ]- ^ Мальпиги впервые описал сосуды ксилемы и назвал клетки трахеиды. Из стр. (Malpighi, 1675) 8 хлопчатобумажные трахеи) распадаются на эту продолговатую и непрерывную полоску. Эти пластинки, при дальнейшем рассмотрении под микроскопом, состоят из мелких частиц, которые обнаруживаются и в трахее этих сосудов, или, так сказать. вернее, из древесных волокон, на которые по длине изделия для большей устойчивости и прочности накладываются N рядов поперечных бутылок, так что оно становится похожим на склад. (... эти [сосуды] трубчатые и несколько круглые, но часто становятся узкими, и они всегда открыты, и ни один из них, [насколько] я мог заметить, не выделяет жидкость: они состоят из серебристых листов L , скрученных в спираль, хотя их легко можно разорвать на эту несколько длинную и связанную полоску (так же, как я это сделал с нитями тутового шелкопряда). частицы, что также наблюдается в трахее насекомых, на этих спиральных сосудах, или, как правильнее сказать, «трахей», стоят древесные нити; M , который вытянут в длину между - для большей прочности и твердости - линиями поперечных ячеек N , так что он имеет форму мата.)
- ^ Хейлз объяснил, что, хотя капиллярное действие может помочь поднять воду внутри ксилемы, транспирация заставляет воду фактически двигаться через ксилему. Из (Хейлз, 1727), с. 100: «И это происходит по тому же [капиллярному] принципу, который мы видим в предыдущих экспериментах, растения так энергично впитывают влагу через свои тонкие капиллярные сосуды; эта влага, поскольку она уносится с потом [т. е. транспирацией] (посредством действие тепла), тем самым дает сосудам с соком свободу почти постоянно привлекать свежие запасы, которые они могли быне делайте этого, если бы они были полностью пропитаны влагой: ибо без пота сок обязательно должен застаиваться, несмотря на то, что сосуды для сока столь любопытно приспособлены из-за своей чрезвычайной тонкости, чтобы поднимать сок на большую высоту, обратно пропорционально их очень маленькому диаметру. ."
Ссылки
[ редактировать ]Цитаты
[ редактировать ]- ^ Перселл, Адам. «Ксилема и флоэма» . Базовая биология . Архивировано из оригинала 4 мая 2016 г.
- ^ Кейт Робертс, изд. (2007). Справочник по растениеводству . Том. 1 (Иллюстрированное изд.). Джон Уайли и сыновья . п. 185. ИСБН 9780470057230 .
- ^ Ричард Б. Манке (1977). Обеспечение энергией: отчет Целевой группы Фонда двадцатого века по энергетической политике США (иллюстрированный ред.). Тата МакГроу-Хилл Образование. п. 42 . ISBN 9780070656178 .
- ^ Нэгели, Карл (1858). «Рост стебля и корня у сосудистых растений и расположение сосудистых нитей на стебле » . Вклад в научную ботанику (на немецком языке). 1 :1-156. Из стр. 9: «Я назову те два, которые образованы камбием снаружи и внутри, флоэмой и ксилемой». (Две части постоянной ткани, образуемые камбием снаружи и внутри, я буду называть «флоэмой» и «ксилемой».)
- ^ Буват, Роджер (1989). «Флоэма». Онтогенез, дифференцировка клеток и строение сосудистых растений . стр. 287–368. дои : 10.1007/978-3-642-73635-3_10 . ISBN 978-3-642-73637-7 .
- ^ Рэйвен, Питер А.; Эверт, Рэй Ф.; Эйххорн, Сьюзен Э. (1999). Биология растений . WH Фриман и компания. стр. 576–577. ISBN 978-1-57259-611-5 .
- ^ Xylem. Архивировано 16 сентября 2011 г. в Wayback Machine . Британская энциклопедия
- ^ Маккалох, Кэтрин А.; Джон С. Сперри; Фредерик Р. Адлер (2003). «Перенос воды в растениях подчиняется закону Мюррея». Природа . 421 (6926): 939–942. Бибкод : 2003Natur.421..939M . дои : 10.1038/nature01444 . ПМИД 12607000 . S2CID 4395034 .
- ^ Jump up to: а б Уолтер С. Джадд (2002). Уолтер С. Джадд (ред.). Систематика растений: филогенетический подход (2-е изд.). У. Х. Фриман. ISBN 0-87893-403-0 .
- ^ Дикисон, WC (2000). Интегративная анатомия растений (стр. 196) . Эльзевир Наука. ISBN 9780080508917 . Архивировано из оригинала 06.11.2017.
- ^ Кох, Джордж В.; Силлетт, Стивен С.; Дженнингс, Грегори М.; Дэвис, Стивен Д. (2004). «Пределы высоты дерева». Природа . 428 (6985): 851–854. Бибкод : 2004Natur.428..851K . дои : 10.1038/nature02417 . ПМИД 15103376 . S2CID 11846291 .
- ^ Кноблаух, Майкл; Кноблаух, Ян; Маллендор, Дэниел Л.; Сэвидж, Джессика А.; Бабст, Бенджамин А.; Бичер, Сьерра Д.; Додген, Адам С.; Дженсен, Кааре Х.; Холбрук, Н. Мишель (2 июня 2016 г.). «Проверка гипотезы Мюнха о транспорте флоэмы на большие расстояния у растений» . электронная жизнь . 5 : е15341. doi : 10.7554/eLife.15341 . ISSN 2050-084X . ПМЦ 4946904 . ПМИД 27253062 .
- ^ Тим Дж. Тиббетс; Фрэнк В. Эверс (2000). «Корневое давление и удельная проводимость лиан умеренного пояса: экзотический Celastrus orbiculatus (Celastraceae) и местный Vitis riparia (Vitaceae)» . Американский журнал ботаники . 87 (9): 1272–78. дои : 10.2307/2656720 . JSTOR 2656720 . ПМИД 10991898 .
- ^ Круизиа, Пьер и Рихтер, Ханно. Физиология растений. Архивировано 28 декабря 2008 г. в Wayback Machine . Синауэр Ассошиэйтс.
- ^ Энтони Р. Йео; Тимоти Дж. Флауэрс, ред. (2007). Транспорт растворов растений . Оксфорд, Великобритания: издательство Blackwell Publishing. п. 221 . ISBN 978-1-4051-3995-3 .
- ^ Нардини, Андреа; Ло Гулло, Мария А.; Саллео, Себастьяно (2011). «Заполнение эмболизированных ксилемных проводников: вопрос разгрузки флоэмы?». Наука о растениях . 180 (4): 604–611. doi : 10.1016/j.plantsci.2010.12.011 . ISSN 0168-9452 . ПМИД 21421408 .
- ^ Диксон, Генри Х.; Джоли, Дж. (1894). «На восхождении сока» . Анналы ботаники . 8 : 468–470.
- ^ Диксон, Генри Х.; Джоли, Дж. (1895). «На восхождении сока» . Философские труды Лондонского королевского общества, серия B. 186 : 563–576. дои : 10.1098/rstb.1895.0012 .
- ^ Тайри, Монтана (1997). «Теория сцепления-напряжения подъема сока: текущие споры» . Журнал экспериментальной ботаники . 48 (10): 1753–1765. дои : 10.1093/jxb/48.10.1753 .
- ^ Ван, З.; Чанг, КК; Хонг, С.-Дж.; Шэн, Ю.-Дж.; Цао, Х.-К. (2012). «Капиллярный подъем в микроканале произвольной формы и смачиваемости: петля гистерезиса». Ленгмюр . 28 (49): 16917–16926. дои : 10.1021/la3036242 . ПМИД 23171321 .
- ^ Аскенасы, Э. (1895). «Ueber das Saftsteigen» [О восхождении сока]. Botanisches Centralblatt (на немецком языке). 62 : 237–238.
- ^ Аскенасы, Э. (1895). « На восхождении сока». Труды Медицинского общества естествознания в Гейдельберге . 2-я серия (на немецком языке). 5 : 325-345.
- ^ Диксон, Х (1914). Транспирация и подъем соков у растений . Лондон, Англия, Великобритания: Macmillan and Co.
- ^ Диксон, Х (1924). Транспирационный поток . Лондон: Лондонский университет Press, Ltd., с. 80.
- ^ Кэмпбелл, Нил (2002). Биология . Сан-Франциско, Калифорния: Pearson Education, Inc., стр. 759 . ISBN 978-0-8053-6624-2 .
- ^ Циммерман, Ульрих (2002). «Каковы движущие силы подъема воды в ксилемном канале?». Физиология Плантарум . 114 (3): 327–335. дои : 10.1034/j.1399-3054.2002.1140301.x . ПМИД 12060254 .
- ^ Тайри, Мелвин Т. (1997). «Теория сцепления-напряжения подъема сока: текущие споры» . Журнал экспериментальной ботаники . 48 (10): 1753–1765. дои : 10.1093/jxb/48.10.1753 .
- ^ Давление водного потенциала ксилемы в стебле вашего растения можно определить с помощью бомбы Шоландера. bio.usyd.edu.au
- ^ Эндрю Дж. МакЭлрон, Брендан Чоат, Грег А. Гамбетта, Крейг Р. Бродерсен (2013). «Поглощение и транспорт воды в сосудистых растениях» . Проект знаний по природному образованию .
{{cite web}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ «Строение растений и грибов | Электронная библиотека» . regi.tankonyvtar.hu (на венгерском языке) . Проверено 2 апреля 2021 г. [ постоянная мертвая ссылка ]
- ^ Карлквист, С.; Э. Л. Шнайдер (2002). «Переход элемента трахеида-сосуд у покрытосеменных включает в себя множество независимых особенностей: кладистические последствия». Американский журнал ботаники . 89 (2): 185–195. дои : 10.3732/ajb.89.2.185 . ПМИД 21669726 .
- ^ Кронквист, А. (август 1988 г.). Эволюция и классификация цветковых растений . Нью-Йорк, Нью-Йорк: Издательство Нью-Йоркского ботанического сада. ISBN 978-0-89327-332-3 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из Сперри, Дж. С. (2003). «Эволюция водного транспорта и структуры ксилемы». Международный журнал наук о растениях . 164 (3): С115–С127. дои : 10.1086/368398 . JSTOR 3691719 . S2CID 15314720 .
- ^ Эдвардс, Д.; Дэвис, КЛ; Акс, Л. (1992). «Сосудистая проводящая нить у раннего наземного растения Cooksonia ». Природа . 357 (6380): 683–685. Бибкод : 1992Natur.357..683E . дои : 10.1038/357683a0 . S2CID 4264332 .
- ^ Никлас, К.Дж.; Смоковит, В. (1983). «Свидетельства наличия проводящей нити у раннесилурийских (лландоверских) растений: значение для эволюции наземных растений». Палеобиология . 9 (2): 126–137. Бибкод : 1983Pbio....9..126N . дои : 10.1017/S009483730000751X . JSTOR 2400461 . S2CID 35550235 .
- ^ Jump up to: а б с д и ж г час Никлас, KJ (1985). «Эволюция диаметра трахеиды у ранних сосудистых растений и ее влияние на гидравлическую проводимость первичной цепи ксилемы». Эволюция . 39 (5): 1110–1122. дои : 10.2307/2408738 . JSTOR 2408738 . ПМИД 28561493 .
- ^ Никлас, К.; Пратт, Л. (1980). «Доказательства наличия лигниноподобных компонентов в окаменелостях растений раннего силура (лландовера)». Наука . 209 (4454): 396–397. Бибкод : 1980Sci...209..396N . дои : 10.1126/science.209.4454.396 . ПМИД 17747811 . S2CID 46073056 .
- ^ Цю, ЮЛ; Ли, Л.; Ван, Б.; Чен, З.; Кноп, В.; Грот-малонек, М.; Домбровская, О.; Ли, Дж.; Кент, Л.; Рест, Дж.; и др. (2006). «Самые глубокие различия у наземных растений, выявленные на основе филогеномных данных» . Труды Национальной академии наук . 103 (42): 15511–6. Бибкод : 2006PNAS..10315511Q . дои : 10.1073/pnas.0603335103 . ПМК 1622854 . ПМИД 17030812 .
- ^ Стюарт, Западная Нью-Йорк; Ротвелл, GW (1993). Палеобиология и эволюция растений . Издательство Кембриджского университета.
- ^ Кораткар, Санджай (24 февраля 2016 г.). «Кавитация и эмболия в сосудистых растениях (со схемой)» . Беседа по биологии .
- ^ Джонсон, Дэниел М.; Маккалох, Кэтрин А.; Вудрафф, Дэвид Р.; Мейнцерк, Фредерик К. (июнь 2012 г.). «Гидравлические запасы прочности и предотвращение эмболии стеблей и листьев: почему хвойные и покрытосеменные растения такие разные?» (PDF) . Наука о растениях . 195 : 48–53. doi : 10.1016/j.plantsci.2012.06.010 . ПМИД 22920998 . Архивировано из оригинала (PDF) 13 марта 2021 г.
- ^ Боуда, Мартин; Хаггетт, Бретт А.; Пратс, Кира А.; Уэйсон, Джей В.; Уилсон, Джонатан П.; Бродерсен, Крейг Р. (11 ноября 2022 г.). «Гидравлический отказ как основной двигатель эволюции сети ксилемы у ранних сосудистых растений» . Наука . 378 (6620): 642–646. Бибкод : 2022Sci...378..642B . дои : 10.1126/science.add2910 . ISSN 0036-8075 . ПМИД 36356120 . S2CID 253458196 .
- ^ Jump up to: а б с д Фостер, А.С.; Гиффорд, Э.М. (1974). Сравнительная морфология сосудистых растений (2-е изд.). Сан-Франциско: WH Freeman. стр. 55–56 . ISBN 978-0-7167-0712-7 .
- ^ Тейлор, Теннесси; Тейлор, Эл.; Крингс, М. (2009). Палеоботаника, биология и эволюция ископаемых растений (2-е изд.). Амстердам; Бостон: Академическая пресса. стр. 207 и след., 212 и далее. ISBN 978-0-12-373972-8 .
- ^ Уайт, А. Тоби; Казлев, М. Алан. «Глоссарий» . palaeos.com . Архивировано из оригинала 20 декабря 2010 года.
- ^ См.:
- Чезальпино, Андреа (1583). De Plantis libri XVI [ О растениях, в 16 книгах ] (на латыни). Флоренция, Италия: Джорджо Марескотти. п. 4 . Из стр. 4: «Или [как] сухие вещи притягивают [то есть поглощают] в соответствии с природой жидкости? [такие как] белье, губки, порошки: ...)
- Беллорини, Кристина (2016). Мир растений в Тоскане эпохи Возрождения: медицина и ботаника . Абингдон-на-Темзе, Англия: Рутледж. п. 72. ИСБН 9781317011491 .
- Крамер, Пол Дж.; Бойер, Джон С. (1995). Водные отношения растений и почв . Лондон, Англия: Elsevier Science. п. 2. ISBN 9780080924113 .
- ^ См.:
- Мальпиги, Марчелло (1675). Anatome Plantarum… (на латыни). Лондон, Англия, Великобритания: Лондонское королевское общество. п. 8.
- Янсен, Стивен; Шенк, Х. Йохен (2015). «О подъёме сока при наличии пузырьков» . Американский журнал ботаники . 102 (10): 1561–1563. дои : 10.3732/ajb.1500305 . ПМИД 26400778 .
- Лэзенби, Элизабет Мэри (1995) « Всеобщая история растений Джона Рэя: Книга I – перевод и комментарии», докторская диссертация, Университет Ньюкасла-апон-Тайн, Англия, Великобритания, том. 1, с. 160. Доступно по адресу: Университет Ньюкасла-апон-Тайн, Великобритания. Архивировано 14 августа 2018 г. в Wayback Machine.
- ^ Грю, Неемия (1682 г.). Анатомия растений… . Лондон, Англия: У. Роулинз. стр. 124–125. Со стр. 124–125: «Большую часть года он [т. е. сок] поднимается в коре [т. е. коре], т. е. на внутренней окраине , прилегающей к лесу , а весной - в или через сам Лес , и только там».
- ^ См.:
- (Грю, 1682), с. 126. Грю признал пределы действия капилляров (со стр. 126): «… маленькие стеклянные трубки [т. е. капиллярные трубки], погруженные в воду, придадут ей (т. е. воде) подъем на несколько дюймов; однако существует определенный период в зависимости от диаметра трубы , после которого она не поднимется». Грю предложил следующий механизм подъема сока у растений (со стр. 126): «Но пузыри [т. е. паренхиматозные клетки] ДП, окружающие его [т. е. столбик трахеид], будучи опухшими и набухшими от сока, , тем самым надавите на него; и таким образом не только немного сузьте его отверстие, но также перелейте или процедите в него некоторую часть своего сока : в обоих случаях сок будет вынужден подняться в нем выше».
- Арбер, Агнес (1913). «Неемия Рос 1641–1712» . В Оливере, Фрэнсис Уолл (ред.). Создатели британской ботаники: сборник биографий ныне живущих ботаников . Кембридж, Англия: Издательство Кембриджского университета. п. 58.
- ^ Хейлз, Стивен (1727). Овощная статика: Или отчет о некоторых статических экспериментах с соком овощей: … . Лондон, Англия: У. и Дж. Иннис и Т. Вудворд. п. 100 . ISBN 9780356030128 .
- ^ См.:
- Страсбургер, Эдуард (1891). материалы Гистологические . (на немецком языке) Том 3: О строении и функции сосудистых пучков у растений . Йена, Германия: Густав Фишер. С. 607–625: Подъем ядовитых жидкостей на значительную высоту у растений , стр. 645–671: Способность погибших частей растений проводить [воду]].
- (Янсен и Шенк, 2015), с. 1561.
Общие ссылки
[ редактировать ]- К. Вэй; Э. Стедле; МТ Тайри; Премьер-министр Линтильяк (май 2001 г.). «Основы прямого измерения давления ксилемы». Растение, клетка и окружающая среда . 24 (5): 549–555. дои : 10.1046/j.1365-3040.2001.00697.x . S2CID 5039439 . является основным источником, использованным в параграфе о недавних исследованиях.
- Н. Мишель Холбрук; Майкл Дж. Бернс; Кристофер Б. Филд (ноябрь 1995 г.). «Отрицательное давление ксилемы в растениях: испытание метода уравновешивающего давления». Наука . 270 (5239): 1193–4. Бибкод : 1995Sci...270.1193H . дои : 10.1126/science.270.5239.1193 . S2CID 97217181 . Это первый опубликованный независимый тест, показывающий, что бомба Шоландер действительно измеряет напряжение в ксилеме.
- Покман, WT; Дж. С. Сперри; Дж. В. О'Лири (декабрь 1995 г.). «Устойчивое и значительное отрицательное давление воды в ксилеме». Природа . 378 (6558): 715–6. Бибкод : 1995Natur.378..715P . дои : 10.1038/378715a0 . S2CID 31357329 . Это второй опубликованный независимый тест, показывающий, что бомба Шоландер действительно измеряет напряжение в ксилеме.
- Кэмпбелл, Нил А.; Джейн Б. Рис (2002). Биология (6-е изд.). Бенджамин Каммингс. ISBN 978-0-8053-6624-2 .
- Кенрик, Пол; Крейн, Питер Р. (1997). Происхождение и ранняя диверсификация наземных растений: кладистическое исследование . Вашингтон, округ Колумбия: Издательство Смитсоновского института. ISBN 978-1-56098-730-7 .
- Мухаммед, А.Ф.; Р. Саттлер (1982). «Сосудистое строение Gnetum и происхождение покрытосеменных». Американский журнал ботаники . 69 (6): 1004–21. дои : 10.2307/2442898 . JSTOR 2442898 .
- Мелвин Т. Тайри; Мартин Х. Циммерманн (2003). Структура ксилемы и восхождение сока (2-е изд.). Спрингер. ISBN 978-3-540-43354-5 . недавнее обновление классической книги покойного Мартина Циммермана о транспорте ксилемы.
Внешние ссылки
[ редактировать ] СМИ, связанные с Ксилемом, на Викискладе?
СМИ, связанные с Ксилемом, на Викискладе?
| Животные | |
|---|---|
| Растения |
|
| Базы данных органов управления : Национальные |
|---|