Аппарат для глазных пятен

Глазно- пятнистый аппарат (или стигма ) представляет собой фоторецепторную органеллу, обнаруженную в жгутиковых или (подвижных) клетках зеленых водорослей и других одноклеточных фотосинтезирующих организмов, таких как эвглениды . Это позволяет клеткам чувствовать направление и интенсивность света и реагировать на него, побуждая организм либо плыть к свету (положительный фототаксис ), либо от него (отрицательный фототаксис). Похожая реакция («фотошок» или фотофобная реакция) возникает, когда клетки кратковременно подвергаются воздействию света высокой интенсивности, заставляя клетку останавливаться, ненадолго плыть назад, а затем менять направление плавания. Восприятие света, опосредованное глазными пятнами, помогает клеткам найти среду с оптимальными световыми условиями для фотосинтеза. Глазные пятна — это самые простые и наиболее распространенные «глаза», встречающиеся в природе, состоящие из фоторецепторов и участков ярких оранжево-красных пигментных гранул. [1] Сигналы, передаваемые от фоторецепторов глазных пятен, приводят к изменению характера биения жгутиков, вызывая фототаксическую реакцию. [2]
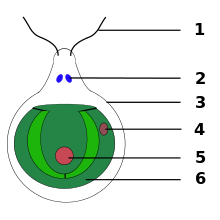
Микроскопическая структура
[ редактировать ]Под световым микроскопом глазные пятна выглядят как темные оранжево-красноватые пятна или стигматы . Свой цвет они получают благодаря каротиноидным пигментам, содержащимся в телах, называемых пигментными гранулами. Фоторецепторы находятся в плазматической мембране, покрывающей пигментированные тельца.
Глазно-пятнистый аппарат эвглены состоит из парафлагеллярного тела, соединяющего глазное пятно со жгутиком . В электронной микроскопии аппарат глазных пятен выглядит как высокоупорядоченная пластинчатая структура, образованная перепончатыми стержнями, расположенными по спирали. [3]
У Chlamydomonas глазное пятно является частью хлоропласта и приобретает вид перепончатой сэндвич-структуры. Он состоит из мембран хлоропластов (внешней, внутренней и тилакоидной мембраны) и заполненных каротиноидами гранул, покрытых плазматической мембраной . Стопки гранул действуют как четвертьволновая пластина , отражая входящие фотоны обратно к вышележащим фоторецепторам, одновременно защищая фоторецепторы от света, идущего с других направлений. Он разбирается во время клеточного деления и преобразуется в дочерних клетках асимметричным образом по отношению к цитоскелету . Такое асимметричное расположение глазного пятна в клетке важно для правильного фототаксиса. [4]
Белки глазных пятен
[ редактировать ]Наиболее важными белками глазных пятен являются белки фоторецепторов , которые воспринимают свет. Фоторецепторы, обнаруженные у одноклеточных организмов, делятся на две основные группы: флавопротеины и ретинилиденовые белки (родопсины). Для флавопротеинов характерно наличие флавина молекул в качестве хромофоров , тогда как ретинилиденовые белки содержат ретиналь . Белок фоторецептора эвглены, вероятно, является флавопротеином. [3] Напротив, фототаксис Chlamydomonas опосредуется родопсинами архейного типа. [5]
Помимо белков фоторецепторов, глазные пятна содержат большое количество структурных, метаболических и сигнальных белков. глазных пятен Протеом клеток Chlamydomonas состоит примерно из 200 различных белков. [6]
Фоторецепция и передача сигнала
[ редактировать ]Фоторецептор эвглены , активируемая синим светом был идентифицирован как аденилатциклаза . [7] Возбуждение этого белка-рецептора приводит к образованию циклического аденозинмонофосфата (цАМФ) в качестве вторичного мессенджера . Химическая передача сигнала в конечном итоге вызывает изменения в характере биения жгутиков и движении клеток.
Родопсины архейного типа Chlamydomonas содержат полностью транс- ретинилиденовый хроматофор, который подвергается фотоизомеризации до 13- цис -изомера. Это активирует канал фоторецептора, что приводит к изменению мембранного потенциала и концентрации ионов кальция в клетках. [5] Передача фотоэлектрического сигнала в конечном итоге вызывает изменения в движениях жгутиков и, следовательно, в движении клеток. [2]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Краймер, Г. (2009). «Аппарат глазных пятен зеленых водорослей: первичная зрительная система и многое другое?». Современная генетика . 55 (1): 19–43. дои : 10.1007/s00294-008-0224-8 . ПМИД 19107486 . S2CID 8011518 .
- ^ Jump up to: а б Хегеманн П. (1997). «Зрение в микроводорослях». Планта . 203 (3): 265–74. Бибкод : 1997Plant.203..265H . дои : 10.1007/s004250050191 . ПМИД 9431675 . S2CID 11933925 .
- ^ Jump up to: а б Волкен Дж (1977). «Эвглена: фоторецепторная система фототаксиса». Дж Протозоол . 24 (4): 518–22. дои : 10.1111/j.1550-7408.1977.tb01004.x . ПМИД 413913 .
- ^ Дикманн С (2003). «Размещение и сборка глазных пятен у зеленой водоросли Chlamydomonas». Биоэссе . 25 (4): 410–6. дои : 10.1002/bies.10259 . ПМИД 12655648 .
- ^ Jump up to: а б Сузуки Т., Ямасаки К., Фудзита С., Ода К., Исеки М., Ёсида К., Ватанабэ М., Дайясу Х., Тох Х., Асамидзу Е., Табата С., Миура К., Фукудзава Х., Накамура С., Такахаши Т. (2003). «Родопсины архейного типа у Chlamydomonas: модельная структура и внутриклеточная локализация». Биохимия Биофиз Рес Коммьюнити . 301 (3): 711–7. дои : 10.1016/S0006-291X(02)03079-6 . ПМИД 12565839 .
- ^ Шмидт М, Гесснер Г, Лафф М, Хейланд И, Вагнер В, Камински М, Геймер С, Эйцингер Н, Райссенвебер Т, Войцех О, Фидлер М, Миттаг М, Креймер Г (2006). «Протеомный анализ глазного пятна Chlamydomonas Reinhardtii дает новое представление о его компонентах и тактических движениях» . Растительная клетка . 18 (8): 1908–30. дои : 10.1105/tpc.106.041749 . ПМЦ 1533972 . ПМИД 16798888 .
- ^ Исеки М, Мацунага С, Мураками А, Оно К, Сига К, Ёсида К, Сугай М, Такахаси Т, Хори Т, Ватанабэ М (2002). «Аденилатциклаза, активируемая синим светом, опосредует избегание фотосъемки у Euglena gracilis». Природа . 415 (6875): 1047–51. Бибкод : 2002Natur.415.1047I . дои : 10.1038/4151047a . ПМИД 11875575 . S2CID 4420996 .
| Базы данных органов управления : Национальные |
|---|