Чешуйчатоногий брюхоногий моллюск
| Чешуйчатоногий / Морской панголин | |
|---|---|
 | |
| Chrysomallon sqamiferum из Лунци. Масштабная линейка составляет 1 см. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Моллюска |
| Сорт: | Брюхоногие моллюски |
| Подкласс: | Неомфалионы |
| Заказ: | Неомфалида |
| Семья: | Пелтоспириды |
| Род: | Хризомаллон Чен, Линс , Копли и Роджерс, 2015 г. |
| Разновидность: | C. sqamiferum |
| Биномиальное имя | |
| Хризомаллон чешуйчатый Чен, Линс, Копли и Роджерс, 2015 г. [2] | |
| Синонимы [2] | |
Crysomallon sqamiferum (орт. ошибка) | |

Chrysomallon sqamiferum , широко известный как чешуйчатый брюхоногий моллюск , чешуйчатая улитка , морской панголин или улитка-вулкан . [3] [4] — вид глубоководной гидротермальной улитки , морского брюхоногого моллюска семейства Peltospiridae . [2] Этот эндемичный для жерл брюхоногий моллюск известен только из глубоководных гидротермальных источников в Индийском океане , где он был обнаружен на глубинах около 2400–2900 м (1,5–1,8 мили). C. sqamiferum сильно отличается от других глубоководных брюхоногих моллюсков, даже от близкородственных неомфалин. [5] В 2019 году он был объявлен находящимся под угрозой исчезновения и занесен в Красный список МСОП. [6] первый вид, внесенный в список таковых из-за рисков, связанных с глубоководной разработкой его жерловой среды обитания. [7]
Оболочка имеет уникальную трехслойную конструкцию; внешний слой состоит из сульфидов железа , средний слой эквивалентен органическому периостракуму, обнаруженному у других брюхоногих моллюсков, а самый внутренний слой состоит из арагонита . Стопа также необычна: по бокам она покрыта минерализованными железом склеритами .
улитки В пищеводной железе обитают симбиотические гаммапротеобактерии, от которых улитка, по-видимому, получает питание. Этот вид считается одним из наиболее своеобразных глубоководных брюхоногих моллюсков гидротермальных источников и единственным известным из ныне живущих животных, у которого в скелете (как в склеритах, так и в панцире в качестве экзоскелета) присутствует сульфид железа. [2] Его сердце, пропорционально говоря, необычно велико для любого животного: сердце составляет примерно 4% объема его тела. [5]
Таксономия
[ редактировать ]Этот вид был впервые обнаружен в апреле 2001 года, а с 2001 года его называют «чешуйчатоногим» брюхоногим моллюском. [8] С 2003 года его называют Chrysomallon sqamiferum , но он не был официально описан в смысле Международного кодекса зоологической номенклатуры до тех пор, пока Chen et al. назвал его в 2015 году. [2] [9] Типовые экземпляры хранятся в Музее естественной истории в Лондоне . [2] Во времена, когда название еще не было официально оформлено, неправильным вариантом написания было « Crysomallon sqamiferum ». [2]
Chrysomallon sqamiferum — типовой вид и единственный вид в роде Chrysomallon . [2] Родовое название Chrysomallon происходит от древнегреческого языка и означает «золотоволосый», поскольку пирит (соединение, встречающееся в его раковине) имеет золотистый цвет. [2] Видовое название sqamiferum происходит из латинского языка и означает «чешуйчатый» из-за его склеритов. [2] Сначала было неизвестно, к какому семейству принадлежит этот вид. [8] Варен и др. в 2003 году отнес этот вид к семейству Peltospiridae, входящему в состав Neomphalina . [10] Молекулярный анализ, основанный на последовательностях генов цитохром-с-оксидазы I (COI), подтвердил отнесение этого вида к Peltospiridae . [2] [11] Морфотипы из двух местонахождений темные; морфотип из третьего местонахождения белый (пояснение местонахождений см. в следующем разделе). [2] [12] [13] Согласно результатам генетического анализа, эти разноцветные улитки кажутся просто «разновидностями» одного и того же вида. [2]
Распределение
[ редактировать ]
Чешуйчатоногий брюхоногий моллюск — это эндемичный брюхоногий моллюск, известный только из глубоководных гидротермальных источников Индийского океана, глубина которых составляет около 2780 метров (1,73 мили). [2] Вид был открыт в 2001 году, обитая на базах черных курильщиков в Кайрейском гидротермальном поле, 25 ° 19,239' ю.ш., 70 ° 02,429' в.д. / 25,320650 ° ю.ш., 70,040483 ° в.д. , на Центрально-Индийском хребте, к северу от Тройной точки Родригес . [8] Впоследствии этот вид также был обнаружен на поле Пасьянс. 19 ° 33,413' ю.ш., 65 ° 50,888' в.д. / 19,556883 ° ю.ш., 65,848133 ° в.д. , Центрально-Индийский хребет , в исключительной экономической зоне Маврикия . [14] [15] и Лунци (в переводе с китайского означает «Флаг Дракона») [16] поле, 37 ° 47,027' ю.ш., 49 ° 38,963' в.д. / 37,783783 ° ю.ш., 49,649383 ° в.д. , Юго-Западный Индийский хребет . [17] [18] Типовым месторождением было обозначено месторождение Лунци; все типы материалов произошли из этого жерлового поля. [2] Расстояние между Кайрей и Солитер составляет около 700 км (430 миль). Расстояние между Пасьянсом и Лунци составляет около 2500 км (1600 миль). [2] Эти три участка принадлежат биогеографической провинции гидротермальных жерловых систем Индийского океана в смысле Роджерса и др. (2012). [19] Расстояние между участками велико, но общая площадь распространения очень мала, менее 0,02 квадратных километров (0,0077 квадратных миль). [20]

Известно, что улитки Peltospiridae в основном обитают в жерловых полях восточной части Тихого океана. Накамура и др. выдвинули гипотезу, что появление чешуйчатоногого брюхоногого моллюска в Индийском океане предполагает родство фаун гидротермальных жерл между этими двумя районами. [14]

Исследовательские экспедиции включали:
- 2000 г. - экспедиция Японского агентства по морским наукам и технологиям о Земле с использованием корабля RV Kairei и ROV Kaikō обнаружила жерловое поле Кайрей, но чешуйчатоногих брюхоногих моллюсков тогда обнаружено не было. [21] Это было первое жерловое поле, обнаруженное в Индийском океане. [21]
- 2001 г. - экспедиция американского исследовательского судна RV Knorr с ROV Jason обнаружила чешуйчатоногих брюхоногих моллюсков в жерлом Кайрейском поле. [8]
- 2007 г. – экспедиция теплохода Да Ян И Хао открыла жерловое поле Лунци. [2]
- 2009 г. - экспедиция НИС «Йокосука» с DSV Shinkai 6500 обнаружила месторождение «Солитер» и отобрала там образцы чешуйчатоногих брюхоногих моллюсков. [14]
- 2009 г. - экспедиция теплохода Да Ян И Хао визуально наблюдала чешуйчатоногих брюхоногих моллюсков на жерлом Лунци. [2] [17]
- 2011 г. - экспедиция британского королевского исследовательского корабля RRS James Cook с ROV Kiel 6000 взяла пробы жерлового поля Лунци. [2] [22]
Описание
[ редактировать ]Склериты
[ редактировать ]
У этого вида стороны стопы улитки чрезвычайно необычны и покрыты сотнями минерализованных железом склеритов ; они состоят из сульфидов железа [10] грейгит и пирит . [23] Каждый склерит имеет ядро из мягкой эпителиальной ткани, конхиолиновую оболочку и самый верхний слой, содержащий пирит и грейгит. [2] До открытия чешуйчатоногих брюхоногих моллюсков считалось, что единственные современные моллюски, обладающие чешуйчатыми структурами, относятся к классам Caudofoveata , Solenogastres и Polyplacophora . [18] Склериты не гомологичны покрышке брюхоногих моллюсков . Склериты чешуйчатоногих брюхоногих моллюсков также не гомологичны склеритам хитонов (Polyplacophora). [18] Было высказано предположение, что склериты кембрийских халваксиид , таких как Halkieria, потенциально могут быть более аналогичны склеритам этой улитки, чем склериты хитонов или аплакофоран. [18] Еще в 2015 году подробный морфологический анализ для проверки этой гипотезы не проводился. [18]
Склериты C. sqamiferum преимущественно белковые (конхиолин — сложный белок); напротив, склериты хитонов преимущественно известковые. [18] На поперечных срезах склеритов видимых линий роста конхиолина нет. [18] Никакие другие современные или вымершие брюхоногие моллюски не имеют кожных склеритов. [18] и ни одно другое современное животное не использует сульфиды железа таким образом ни в своем скелете , ни в скелете . [2] или экзоскелет .
Размер каждого склерита у взрослых составляет около 1 × 5 мм. [2] У молодых особей чешуя расположена в несколько рядов, а у взрослых чешуя плотная и асимметричная. [24] Популяция улиток Solitaire имеет белые склериты вместо черных; это связано с недостатком железа в склеритах. [18] Склериты черепитчатые (накладываются друг на друга, напоминающие черепицу ). [5] Предполагается, что целью склеритов является защита или детоксикация . [25] Склериты могут помочь защитить брюхоногих моллюсков от вентиляционной жидкости, чтобы их бактерии могли жить рядом с источником доноров электронов для хемосинтеза. [5] Или, альтернативно, склериты могут возникнуть в результате отложения токсичных сульфидных отходов эндосимбионтов и, следовательно, представляют собой новое решение для детоксикации. [5] Но истинная функция склеритов пока неизвестна. [14] Склериты Кайрейской популяции, имеющие слой сульфида железа, ферримагнитны . [2] Нежелезосульфидно-минерализованный склерит морфотипа Solitaire показал большую механическую прочность всей конструкции при трехточечном изгибе (12,06 МПа), чем склерит морфотипа Kairei (6,54 МПа). [14]
При жизни на внешней поверхности склеритов обитает разнообразный набор эпибионтов : Campylobacterota (ранее Epsilonproteobacteria) и Thermodesulfobacteriota (ранее часть Deltaproteobacteria). [26] Эти бактерии, вероятно, обеспечивают их минерализацию. [26] Гоффреди и др. (2004) предположили, что улитка выделяет некоторые органические соединения, которые облегчают прикрепление бактерий. [26]
Оболочка
[ редактировать ]
Кайрей, Лунци, Пасьянс (слева направо)
Раковина этих видов имеет три оборота . [2] Форма корпуса шаровидная, шпиль сжатый. [2] Скульптура ракушки состоит из ребер и тонких линий роста. [2] Форма отверстия эллиптическая . [2] Вершина панциря хрупкая, у взрослых особей подвергается коррозии. [2]
Это очень крупный пельтоспирид по сравнению с большинством других видов, которые обычно ниже 15 миллиметров ( 3 ⁄ дюйма ). Длина корпуса [2] Ширина раковины 9,80–40,02 мм (0,39–1,58 дюйма); [2] максимальная ширина корпуса достигает 45,5 миллиметров (1,79 дюйма). [2] Средняя ширина раковины взрослых улиток составляет 32 мм. [2] Средняя ширина раковины в популяции Solitaire была несколько меньше, чем в популяции Kairei. [15] Высота раковины 7,65–30,87 мм (0,30–1,22 дюйма). [2] Ширина апертуры составляет 7,26–32,52 мм (0,29–1,28 дюйма). [2] Высота апертуры составляет 6,38–27,29 мм (0,25–1,07 дюйма). [2]
Структура оболочки состоит из трех слоев. Внешний слой толщиной около 30 мкм, черного цвета, выполнен из сульфидов железа, содержит грейгит Fe 3 S 4 . [27] Этот вид — единственное известное из ныне живущих животных, в скелете которого присутствует этот материал. [2] Средний слой (около 150 мкм) эквивалентен органическому периостракуму, который встречается и у других брюхоногих моллюсков. [27] Периостракум толстый, коричневый. [2] Самый внутренний слой состоит из арагонита (толщиной около 250 мкм) — формы карбоната кальция , которая обычно встречается как в раковинах моллюсков, так и в различных кораллах. [27] Цвет слоя арагонита молочно-белый. [2]
Каждый слой раковины по-разному способствует эффективности защиты улитки. Средний органический слой, по-видимому, поглощает механическое напряжение и энергию, возникающую при сдавливании (например, клешнями краба), что делает панцирь намного более прочным. Органический слой также рассеивает тепло. [28] Особенности этого композиционного материала находятся в центре внимания исследователей для возможного использования в гражданских и военных защитных целях. [27]
 Две разновидности чешуйчатоногих брюхоногих моллюсков
Две разновидности чешуйчатоногих брюхоногих моллюсков C. sqamiferum из жерлового поля Кайрей
C. sqamiferum из жерлового поля Кайрей C. sqamiferum из жерлового поля Солитер
C. sqamiferum из жерлового поля Солитер



Оперкулум
[ редактировать ]У этого вида форма жаберной крышки меняется в процессе роста: от округлой у молоди до изогнутой у взрослых. [14] Относительный размер жаберной крышки уменьшается по мере роста особи. [5] Около половины всех взрослых улиток этого вида имеют жаберную крышку среди склеритов в задней части животного. [14] Кажется вероятным, что склериты постепенно разрастаются и полностью покрывают всю стопу для защиты, а жаберная крышка теряет свою защитную функцию по мере роста животного. [14]
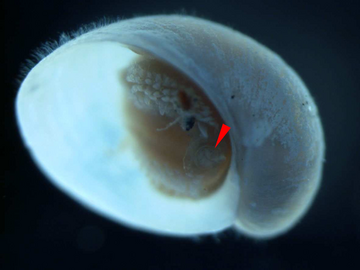 Молодь с жаберной крышкой обозначена красным указателем. Длина раковины около 2 мм.
Молодь с жаберной крышкой обозначена красным указателем. Длина раковины около 2 мм. Крышка молодой улитки. Масштабная линейка составляет 1 мм.
Крышка молодой улитки. Масштабная линейка составляет 1 мм. Крышка взрослой улитки. Масштабная линейка составляет 1 мм.
Крышка взрослой улитки. Масштабная линейка составляет 1 мм. Взрослые улитки с крышечкой, обозначенной красными стрелками. Масштабная линейка составляет 5 мм.
Взрослые улитки с крышечкой, обозначенной красными стрелками. Масштабная линейка составляет 5 мм.




Внешняя анатомия
[ редактировать ]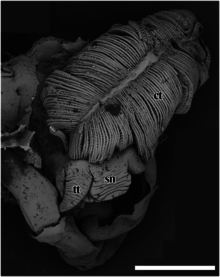
sn – морда,
тт – головное щупальце.
Масштабная линейка составляет 2 мм.
Чешуйчатоногий брюхоногий моллюск имеет толстую морду, которая сужается дистально к тупому концу. Рот представляет собой круглое кольцо мышц в сжатом и закрытом состоянии. [5] Два гладких головных щупальца толстые у основания и постепенно сужаются к тонким кончикам на дистальных концах. [5] У этой улитки нет глаз. [5] Специализированного копулятивного придатка нет. [5] Ножка красная и большая, и улитка не может полностью убрать ее в раковину. [2] отсутствует . Педальная железа в передней части стопы [5] Эпиподиальных щупалец также нет. [5]
Внутренняя анатомия
[ редактировать ]У C. sqamiferum мягкие части животного занимают примерно два оборота внутренней части раковины. [5] Панцирная мышца подковообразной формы, крупная, разделена на две части слева и справа и соединена более узким прикреплением. [5] Край мантии толстый, простой, без каких-либо отличительных черт. [5] Мантийная полость глубокая, доходит до заднего края раковины. [5] В медиальной левой части полости преобладает очень большой двугребешковый ктенидий . [5] Вентральнее висцеральной массы полость тела занята огромной пищеводной железой, которая простирается и заполняет вентральное дно мантийной полости. [5] [26]
проста Пищеварительная система , ее объем уменьшен до менее чем 10% от объема, типичного для брюхоногих моллюсков. [5] [26] Радула . «слабая», типа рифидоглоссана, с одной парой радулярных хрящей [5] [26] Формула радулы : ~50 + 4 + 1 + 4 + ~50. [2] Лента радулы длиной 4 мм, шириной 0,5 мм; [2] соотношение ширины и длины составляет примерно 1:10. [5] . Челюсти и слюнных желез нет [5] Часть переднего отдела пищевода быстро разрастается в огромную гипертрофированную пищеводную железу со слепыми концами, занимающую большую часть вентральной поверхности мантийной полости (около 9,3% объема тела). [5] Пищеводная железа растет изометрически вместе с улиткой, что соответствует улитке, зависящей от ее эндосимбионтных микробов на протяжении всей ее оседлой жизни. [24] Пищеводная железа имеет однородную структуру, хорошо васкуляризирована и имеет мелкие кровеносные сосуды. [5] Желудок пищеварительной имеет как минимум три протока в передней правой части, соединяющихся с железой . [5] Консолидированные гранулы имеются как в желудке, так и в задней кишке. [5] Эти гранулы, вероятно, представляют собой гранулы серы, производимые эндосимбионтом для детоксикации сероводорода. [5] Кишка редуцирована и имеет только одну петлю. [5] Обширная и рыхлая пищеварительная железа простирается назад, заполняя вершину раковины . [5] Прямая кишка не проникает в сердце, а проходит вентрально от него. [5] Анус . расположен на правой стороне улитки, над половым отверстием [5]
В выделительной системе нефридий . занимает центральное место, тяготеет к правой стороне тела, представляет собой тонкий темный слой железистой ткани [5] Нефридий находится в передней и вентральной части пищеварительной железы и соприкасается с дорсальной стороной передней кишки. [5]
Дыхательная система и система кровообращения состоят из единственного левого двугребешкового ктенидия (жаберного), очень большого размера (15,5% объема тела), поддерживаемого множеством крупных и подвижных кровеносных синусов, заполненных гемоцелем . [5] [24] При вскрытии заметным признаком всей полости тела являются кровеносные синусы и комки материала гемоцеля. [5] Хотя система кровообращения у Chrysomallon в основном закрыта (это означает, что гемоцель в основном не покидает кровеносные синусы), выступающие кровеносные синусы кажутся временными и возникают в разных областях тела у разных людей. [24] По обе стороны от ктенидия имеются тонкие жаберные нити. [5] Двояковыпуклый ктенидий простирается далеко позади сердца до верхних оборотов раковины; он значительно крупнее, чем у Peltospira . Хотя этот вид имеет форму раковины и общую форму, сходную с другими пельтоспиридами, ктенидий пропорционален размеру Hirtopelta , у которого самые большие жабры среди родов пельтоспирид, которые были исследованы анатомически до сих пор. [5]
Ктенидий обеспечивает улитку кислородом, но ее кровеносная система расширена и выходит за рамки возможностей других подобных жерловых брюхоногих моллюсков. [5] Эндосимбионтов в жабрах C. sqamiferum и на них нет . [5] Увеличение жабр, вероятно, призвано облегчить извлечение кислорода в условиях низкого содержания кислорода, типичных для гидротермальных жерловых экосистем. [5]
В задней части ктенидия находится удивительно большое и хорошо развитое сердце. [5] Сердце необычайно велико для любого пропорционального животного. [5] В зависимости от объема отдельного предсердия и желудочка сердечный комплекс составляет примерно 4% объема тела (например, сердце человека составляет 1,3% объема тела). [5] Длина желудочка у молодых особей составляет 0,64 мм, длина оболочки 2,2 мм, у взрослых вырастает до 8 мм. [24] Это пропорционально гигантское сердце в основном всасывает кровь через ктенидий и снабжает хорошо васкуляризированную пищеводную железу. [5] У C. sqamiferum эндосимбионты обитают в пищеводной железе, где они изолируются от вентиляционной жидкости. [5] Таким образом, хозяин, вероятно, будет играть важную роль в снабжении эндосимбионтов необходимыми химическими веществами, что приводит к увеличению респираторных потребностей. [5] Детальное исследование гемоцеля C. sqamiferum позволит получить дополнительную информацию о его дыхательных пигментах. [5]

ct – ктенидий,
пм – педальная мышца,
сбн – весы,
если - кровь синусовая,
ты – яичко.
Масштабная линейка равна 1 см.
Чешуйчатоногий брюхоногий моллюск — хемосимбиотический холобионт . [26] Он содержит тиоавтотрофные (окисляющие серу) гаммапротеобактериальные эндосимбионты в значительно увеличенной пищеводной железе и, по-видимому, использует эти симбионты для питания. [26] [29] Ближайшим известным родственником этого эндосимбионта является улитка Alviniconcha . [30] У этого вида размеры пищеводной железы примерно на два порядка превышают обычные размеры. [26] Внутри пищеводной железы имеется значительное разветвление, где артериальное давление, вероятно, снижается почти до нуля. [5] Сложная сердечно-сосудистая система, скорее всего, развилась для насыщения эндосимбионтов кислородом в среде с низким содержанием кислорода и/или для снабжения сероводородом . эндосимбионтов [5] Тиоавтотрофные гаммапротеобактерии обладают полным набором генов, необходимых для аэробного дыхания , и, вероятно, способны переключаться между более эффективным аэробным дыханием и менее эффективным анаэробным дыханием в зависимости от доступности кислорода. [5] В 2014 году эндосимбионт чешуйчатоногого брюхоногого моллюска стал первым эндосимбионтом любого брюхоногого моллюска, полный геном которого был известен. [29] C. sqamiferum является единственным видом Peltospiridae, имеющим увеличенную пищеводную железу. Ранее считалось, что [2] но позже было обнаружено, что оба вида Gigantopelta имеют также увеличенную пищеводную железу. [11] Chrysomallon и Gigantopelta — единственные жерловые животные, за исключением сибоглинидных трубчатых червей, у которых эндосимбионты обитают в замкнутой части тела, не находящейся в прямом контакте с жерловой жидкостью. [24]
Нервная система крупная, а мозг представляет собой сплошную нервную массу без ганглиев. [5] Нервная система усложнена и увеличена в размерах по сравнению с другими таксонами неомфалинов. [5] Как это типично для брюхоногих моллюсков, нервная система состоит из переднего пищеводного нервного кольца и двух пар продольных нервных тяжей: вентральной пары, иннервирующей стопу, и дорсальной пары, образующей скручивание посредством стрептоневрии . [5] Фронтальная часть пищеводного нервного кольца крупная, соединяет два латеральных вздутия. [5] Огромная сросшаяся нервная масса непосредственно прилегает к пищеводной железе, где обитают бактерии, и проходит через нее. [5] В головные щупальца входят большие щупальцевые нервы. [5] Органы чувств чешуйчатоногих брюхоногих включают статоцисты, окруженные пищеводной железой, причем каждый статоцист содержит один статолит . [5] На кончике жаберных лепестков имеются также чувствующие ктенидальные бурсикулы; Известно, что они присутствуют у большинства ветигастропод, а также у некоторых неомфалинов. [5]
Репродуктивная система имеет некоторые необычные особенности. Гонады взрослых улиток не находятся внутри раковины; они находятся в области головы и стоп на правой стороне тела. [5] У молоди с длиной раковины 2,2 мм гонады отсутствуют. [24] находятся У взрослых и семенники , и яичники на разных уровнях развития. [5] Семенник расположен вентрально; яичник располагается дорсально, а нефридий лежит между ними. [5] Рядом с семенником находится «орган упаковки сперматофоров». [5] Гонодукты семенника и яичника первоначально разделены, но, по-видимому, сливаются в один проток и возникают в виде единого генитального отверстия справа от мантийной полости. [5] У животного нет копулятивного органа. [2] [5]
 | 
| 
|
Предполагается, что разработанная стратегия размещения эндосимбиотических микробов в пищеводной железе стала катализатором анатомических инноваций, которые служат в первую очередь улучшению приспособленности бактерий сверх потребностей улитки. [5] Сильное увеличение пищеводной железы, защитные кожные склериты улитки, ее сильно увеличенная дыхательная и кровеносная системы, а также высокая плодовитость — все это считается адаптациями, полезными для ее эндосимбионтных микробов. [5] Эти адаптации, по-видимому, являются результатом специализации по удовлетворению энергетических потребностей в экстремальной хемосинтетической среде. [5]
Экология
[ редактировать ]среда обитания
[ редактировать ]Этот вид населяет гидротермальные жерла Индийского океана. Он обитает рядом как с кислой, так и с восстановительной жидкостью, на стенках дымоходов черных курильщиков или непосредственно в местах диффузного потока. [5]
Глубина месторождения Кайрей варьируется от 2415 до 2460 м (от 7923 до 8071 футов). [8] его размеры составляют примерно 30 на 80 м (98 на 262 фута). [8] Уклон поля составляет от 10° до 30°. [8] Порода - субстрат представляет собой троктолит и истощенный базальт срединно-океанического хребта . [31] Чешуйчатоногие брюхоногие Кайрейские поля обитают в низкотемпературных диффузных жидкостях одного дымохода. [14] Переходная зона, где были обнаружены эти брюхоногие моллюски, имеет ширину около 1–2 м (3–7 футов) и температуру 2–10 °C. [32] Предпочтительная температура воды для этого вида составляет около 5 °C. [33] Эти улитки живут в среде с высокой концентрацией сероводорода и низкой концентрацией кислорода. [33]
Численность чешуйчатоногих брюхоногих на поле Кайрей была ниже, чем на поле Лунци. [2] Сообщество Кайрейского гидротермального источника состоит из 35 таксонов. [34] включая морские анемоны Marianactis sp., ракообразные Austinograea Rodriguezensis , Rimicaris kairei , Mirocaris indica , Munidopsis sp., род и виды Neolepadidae , Eochionelasmus sp., двустворчатые моллюски Bathymodiolus marisindicus , брюхоногие моллюски Lepetodrilus sp., Pseudorimula sp., Eulepetopsis sp., с сп ., и Alviniconcha marisindica , [35] Дебрюйерезия марисиндика , [36] Брюсиелла варени , [36] Phymorhynchus sp., Sutilizona sp., блюдечко sp. 1, щелевое блюдечко зр. 2, Ифинопсис бушети , [36] Соленогастрес Хеликорадомения ? sp., кольчатые черви Amphisamytha sp., Archinome jasoni , Capitellidae sp. 1, Ophyotrocha sp., Hesionidae sp. 1, Hesionoidae sp. 2, Branchinotogluma sp., Branchipolynoe sp., Harmothoe ? sp., Левенштейниелла ? sp., Prionospio sp., неопознанная Nemertea и неопознанная Platyhelminthes . [34] Чешуйчатоногие брюхоногие моллюски обитают колониями с улитками Alviniconcha marisindica , а над ними располагаются колонии Rimicaris kairei . [33]
Поле Пасьянс находится на глубине 2606 м (8550 футов), а его размеры составляют примерно 50 на 50 м (160 на 160 футов). [14] Порода-субстрат представляет собой обогащенный базальт срединно-океанического хребта. [14] [31] Чешуйчатоногие брюхоногие моллюски обитают вблизи высокотемпературных диффузных жидкостей дымовых труб жерлового поля. [14] Численность чешуйчатоногих брюхоногих на поле Солитер была ниже, чем на поле Лунци. [2] Сообщество гидротермальных источников Solitaire включает 22 таксона, в том числе: морские анемоны Marianactis sp., ракообразные Austinograea Rodriguezensis , Rimicaris kairei , Mirocaris indica , Munidopsis sp., Neolepadidae gen et sp., Eochionelasmus sp., двустворчатые моллюски Bathymodiolus marisindicus , брюхоногие моллюски Lepetodrilus sp. , Eulepetopsis sp., Shinkailepas sp., Alviniconcha sp. тип 3, Desbruyeresia sp., Phymorhynchus sp., кольчатые черви рода и вида Alvinellidae , Archinome jasoni , Branchinotogluma sp., иглокожие голотурии Apodacea gen et sp., рыбы рода Macrouridae и виды, неопознанные Nemertea и неопознанные Platyhelminthes . [34]

Поле жерл Лунци находится на глубине 2780 м (9120 футов). [2] его размеры составляют примерно 100 на 150 м (330 на 490 футов). [20] C. sqamiferum был густо заселен в районах, непосредственно окружающих диффузное жерло. [5] Сообщество гидротермальных источников Лунци включает 23 [Примечание 1] таксоны макро- и мегафауны: морские анемоны Actinostolidae sp., кольчатые черви Polynoidae n. ген. н. сп. «655», Бранчиполыное с. сп. «Дракон», Пейналеополиное с. сп. «Дракон», Гезиолира ср. bergi , Hesionidae sp. индет., Ophryotrocha n. сп. «F-038/1b», Прионоспий ср. одноламелла , Ampharetidae sp. indet., мидии Bathymodiolus marisindicus , брюхоногие моллюски Gigantopelta aegis , [11] Дракогира подфускус , Лирапекс полированный , [16] Фиморинх н. сп. «SWIR» , Lepetodrilus n. сп. «SWIR», ракообразные Neolepas sp. 1, Rimicaris kairei , Mirocaris indica , Chorocaris sp., Kiwa n. сп. «SWIR»17, Munidopsis sp. и иглокожие голотурии Chiridota sp. [17] [37] Плотность Lepetodrilus n. сп. «SWIR» и чешуйчатоногих брюхоногих моллюсков – более 100 улиток на м2. 2 на близком расстоянии от источников жерловой жидкости на жерловом поле Лунци. [37]
Привычки питания
[ редактировать ]Чешуйчатоногий брюхоногий моллюск является облигатным симбиотрофом на протяжении всей постпоселенческой жизни. [24] На протяжении всей постличиночной жизни чешуйчатоногие брюхоногие получают все свое питание за счет хемоавтотрофии своих эндосимбиотических бактерий. [26] [24] Чешуйчатоногий брюхоногий моллюск не является фильтратором. [5] [24] и не использует другие механизмы для кормления. [5] Радула и хрящ радулы небольшие и составляют соответственно лишь 0,4% и 0,8% объема тела молоди по сравнению с 1,4% и 2,6% у миксотрофной молоди Gigantopelta Chessoia . [24]
Для идентификации трофических взаимодействий в среде обитания, где прямое наблюдение за пищевыми привычками затруднено, можно измерить состав стабильных изотопов углерода и азота. [32] Имеются обедненные значения δ 13 C в пищеводной железе (по отношению к органическому углероду, полученному фотосинтезом). [26] Источником такого углерода считались хемоавтотрофные симбионты. [26] Хемоавтотрофное происхождение стабильного изотопа углерода 13 С было подтверждено экспериментально. [29]
| Салфетка | д 13 С | д 15 Н |
|---|---|---|
| Пищеводная железа | −20.7 ± 0.9 ‰ | 3.3 ± 1.8 ‰ |
| Джилл | −18,3 ± 0,6 ‰, от −17,4 до −18,8 ‰ | 3,9 ± 0,6 ‰, от 3,1 до 4,2 ‰ |
| Мантия | от −17,5 до −18,6 ‰ | от 3,5 до 4,7 ‰ |
| Ступня | −18.2 ± 0.6 ‰ | 3.8 ± 0.5 ‰ |
| Весы | −16.7 ± 0.6 ‰ | 3.8 ± 0.9 ‰ |
Жизненный цикл
[ редактировать ]Этот брюхоногий моллюск одновременно является гермафродитом . [5] Это единственный вид семейства Peltospiridae, который, как известно, одновременно является гермафродитом. [5] Имеет высокую плодовитость . [5] Откладывает яйца, вероятно, лецитотрофного типа. [22] Яйца чешуйчатоногого брюхоногого моллюска обладают отрицательной плавучестью под атмосферным давлением. [15] По состоянию на 2016 год ни личинки, ни протоконх не известны, но считается, что этот вид находится на стадии планктонного расселения. [22] Самые маленькие из когда-либо собранных молодых особей C. sqamiferum имели длину раковины 2,2 мм. [24] Результаты статистического анализа не выявили генетической дифференциации между двумя популяциями в полях Кайрей и Солитер, что позволяет предположить потенциальную связь между двумя жерловыми полями. [15] Популяция Кайрей представляет собой потенциальную исходную популяцию для двух популяций Центрально-Индийского хребта. [15] Этих улиток трудно сохранить в искусственной среде; однако они выживали в аквариумах при атмосферном давлении более трех недель. [33]
Меры по сохранению и угрозы
[ редактировать ]Чешуйчатоногий брюхоногий моллюск не защищен. [1] [20] Его потенциальная среда обитания во всех гидротермальных жерлах Индийского океана оценивается не более чем в 0,27 квадратных километров (67 акров), в то время как три известных участка, в которых он был обнаружен, между которыми происходит лишь незначительная миграция, [38] добавить до 0,0177 квадратных километров (4,4 акра), [1] или менее одной пятой футбольного поля. [20]
Население жерлового поля Лунци может вызывать особую озабоченность. Юго-Западный Индийский хребет, в пределах которого он расположен, является одним из срединно-океанических хребтов , распространяющихся медленнее всего , а низкий уровень естественных нарушений связан с экологическими сообществами , которые, вероятно, более чувствительны к разрушениям и медленнее восстанавливаются после них. Медленно распространяющиеся центры могут также создавать более крупные месторождения полезных ископаемых, что делает эти чувствительные районы основными объектами глубоководной добычи полезных ископаемых . Более того, по генетическим показателям популяция Лунци плохо связана с популяциями жерловых полей Кайрей и Солитер, находящихся на расстоянии более 2000 км в пределах Центрально-Индийского хребта. [20]
Месторождение Солитер находится в исключительной экономической зоне Маврикия, тогда как два других участка находятся в пределах зон за пределами национальной юрисдикции (широко известных как открытое море ) под управлением Международного органа по морскому дну , который выдал лицензии на коммерческую разведку месторождений для обоих месторождений. . Лицензия на месторождение Кайрей принадлежит Германии (2015–2030 гг.), а месторождение Лунци – Китаю (2011–2026 гг.). По состоянию на 2017 год никаких мер по сохранению ни для одного из трех объектов не предложено и не применяется. [20]
он внесен в Красный список исчезающих видов МСОП как вид, находящийся под угрозой исчезновения . С 4 июля 2019 года [1]
См. также
[ редактировать ]Примечания
[ редактировать ]- ^ По состоянию на 2016 год в Лунци был известен 21 вид, а в 2017 году были описаны два новых брюхоногих моллюска.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д Сигварт, Дж.; Чен, К.; Томас, Э.А. (2019). « Хризомаллон чешуйчатый » . Красный список исчезающих видов МСОП . 2019 : e.T103636217A103636261. doi : 10.2305/IUCN.UK.2019-2.RLTS.T103636217A103636261.en . Проверено 19 ноября 2021 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а к ап ак с как в Чен, Чонг; Линсе, Катрин; Копли, Джонатан Т.; Роджерс, Алекс Д. (2015). «Чешуйчатоногий брюхоногий моллюск: новый род и вид эндемичных для гидротермальных источников брюхоногих моллюсков (Neomphalina: Peltospiridae) из Индийского океана» . Журнал исследований моллюсков . 81 (3): 322–334. дои : 10.1093/mollus/eyv013 .
- ^ «Познакомьтесь с причудливой морской улиткой, которая строит собственный железный доспех» . 15 ноября 2021 г.
- ^ Сигварт, Джулия Д.; Чен, Чонг; Томас, Элин А.; Олкок, А. Луиза; Бём, Моника; Седдон, Мэри (22 июля 2019 г.). «Красный список может защитить глубоководное биоразнообразие» . Экология и эволюция природы . 3 (8): 1134. Бибкод : 2019NatEE...3.1134S . дои : 10.1038/s41559-019-0930-2 . ISSN 2397-334X . ПМИД 31332328 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а к ап ак с как в В из хорошо топор является тот нет бб до нашей эры др. быть парень бг чб с минет БК с бм млрд быть б.п. БК бр бс БТ этот бв Чен, Чонг; Копли, Джонатан Т.; Линсе, Катрин; Роджерс, Алекс Д.; Сигварт, Джулия Д. (2015). «Сердце дракона: 3D анатомическая реконструкция чешуйчатого брюхоногого моллюска (Mollusca: Gastropoda: Neomphalina) раскрывает его необычную систему кровообращения» . Границы в зоологии . 12:13 . дои : 10.1186/s12983-015-0105-1 . ПМЦ 4470333 . ПМИД 26085836 .
- ^ «Красный список видов, находящихся под угрозой исчезновения» МСОП . Красный список исчезающих видов МСОП . Проверено 28 июля 2019 г.
- ^ Ламберт, Джонатан (22 июля 2019 г.). «Океанская улитка — первое животное, официально находящееся под угрозой исчезновения в результате глубоководной добычи полезных ископаемых» . Природа . 571 (7766): 455–456. Бибкод : 2019Natur.571..455L . дои : 10.1038/d41586-019-02231-1 . ПМИД 31337912 .
- ^ Перейти обратно: а б с д и ж г Довер, Синди Л. Ван; Хамфрис, ЮВ; Форнари, Д.; Кавано, CM; Кольер, Р.; Гоффреди, Шана К.; Хашимото, Дж.; Лилли, доктор медицины; Рейзенбах, Алабама; Шанк, ТМ; Фон Дамм, КЛ; Банта, А.; Галлант, РМ; Гоц, Д.; Грин, Д.; Холл, Дж.; Хармер, ТЛ; Уртадо, Луизиана; Джонсон, П.; МакКинесс, З.П.; Мередит, К.; Олсон, Э.; Пан, Иллинойс; Репка, М.; Вон, Ю.; Янг, CR 3-е место; Вриенгук, Р.К. (2001). «Биогеография и экологическая обстановка гидротермальных источников Индийского океана» . Наука . 294 (5543): 818–23. Бибкод : 2001Sci...294..818V . дои : 10.1126/science.1064574 . ПМИД 11557843 . S2CID 543841 .
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Буше, П. (2014). «Хризомаллон чешуйчатый» . Всемирный реестр морских видов . Проверено 22 апреля 2015 г.
- ^ Перейти обратно: а б Варен, Андерс; Бенгтсон, Стефан; Гоффреди, Шана К.; Дувр, Синди Л. Ван (2003). «Брюхоногий моллюск с горячими жерлами и дермальными склеритами из сульфида железа». Наука . 302 (5647): 1007. doi : 10.1126/science.1087696 . ПМИД 14605361 . S2CID 38386600 .
- ^ Перейти обратно: а б с Чен, Чонг; Линсе, Катрин; Ротерман, Кристофер Н.; Копли, Джонатан Т.; Роджерс, Алекс Д. (2015). «Новый род крупных гидротермальных эндемичных брюхоногих моллюсков (Neomphalina: Peltospiridae)» (PDF) . Зоологический журнал Линнеевского общества (Представлена рукопись). 175 (2): 319–335. дои : 10.1111/zoj.12279 .
- ^ (на японском языке) (2010) «Первое в мире открытие белой чешуйчатой стопы без сульфида железа ~ Результаты новых гидротермальных исследований в Индийском океане ~» Японское агентство морских наук и технологий , Токийский университет , Университет Коти . пресс-релиз). Проверено 16 июля 2016 г.
- ^ «Новый чешуйчатоногий брюхоногий моллюск обнаружен в Индийском океане; открытие белого чешуйчатого брюхоногого моллюска» . Южная жареная наука. 1 июля 2011 года . Проверено 16 июля 2016 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л Накамура, Кентаро, Хироми, Дзюнъити, Кен; Ногучи, Такуро, Сугуру; Мацудзаки, Такуя, Такадзо; Орихаши, Юджи; Асада, Акира; Кунджул, Мира; Бидесси, Гириш; Тамаки, Шнур, Джоэл М. (2012) . Активность и хемосинтетическая фауна Центрально-Индийского хребта на 18°–20°ю.ш.» . PLOS ONE . 7 (3): e32965. Bibcode : 2012PLoSO...732965N . doi : 10.1371/journal.pone.0032965 . PMC 3303786 . ПМИД 22431990
- ^ Перейти обратно: а б с д и Бидесси, Гириш; Ватанабэ, Хироми; Огура, Томоми; Немото, Сугуру; Яхаги, Такуя; Накагава, Сатоши; Накамура, Кентаро; Такай, Кен; Кунджул, Мира; Мари, Дэниел EP (2013). «Высокая связанность популяций животных в глубоководных гидротермальных полях Центрально-Индийского хребта в связи с его геологическими условиями» . ПЛОС ОДИН . 8 (12): е81570. Бибкод : 2013PLoSO...881570B . дои : 10.1371/journal.pone.0081570 . ПМЦ 3864839 . ПМИД 24358117 .
- ^ Перейти обратно: а б Чен, Чонг; Чжоу, Ядун; Ван, Чуньшэн; Копли, Джонатан Т. (2017). «Две новые улитки-пельтоспириды с горячими источниками (Gastropoda: Neomphalina) из гидротермального поля Лунци, юго-западный Индийский хребет» . Границы морской науки . 4 . дои : 10.3389/fmars.2017.00392 . ISSN 2296-7745 .
- ^ Перейти обратно: а б с Тао, Чуньхуэй; Линь, Цзянь; Го, Шицинь; Чен, Юншун Джон; Ву, Гуанхай; Хан, Сицю; Герман, Кристофер Р.; Йоргер, Дана Р.; Чжоу, Нин; Ли, Хуаймин; Су, Синь; Чжу, Цзянь (2012). «Первые активные гидротермальные источники в центре сверхмедленного расширения: Юго-Западный Индийский хребет». Геология . 40 (1). DY115-19 (этапы 1–2) и DY115-20 (этапы 4–7) Научные вечеринки: 47–50. Бибкод : 2012Geo....40...47T . дои : 10.1130/G32389.1 .
- ^ Перейти обратно: а б с д и ж г час я Чен, Чонг; Копли, Джонатан Т.; Линсе, Катрин; Роджерс, Алекс Д.; Сигварт, Джулия (2015). «Как у моллюска появилась чешуя: конвергентная эволюция склеритома моллюска» . Биологический журнал Линнеевского общества . 114 (4): 949–954. дои : 10.1111/bij.12462 .
- ^ Роджерс, Алекс Д.; Тайлер, Пол А.; Коннелли, Дуглас П.; Копли, Джон Т.; Джеймс, Рэйчел; Лартер, Роберт Д.; Линсе, Катрин; Миллс, Рэйчел А.; Гарабато, Альфредо Навейра; Панкост, Ричард Д.; Пирс, Дэвид А.; Полунин, Николай В.К.; Герман, Кристофер Р.; Шанк, Тимоти; Бурш-Супан, Филипп Х.; Алкер, Белинда Дж.; Аквилина, Альфред; Беннетт, Сара А.; Кларк, Эндрю; Динли, Роберт Джей-Джей; Грэм, Аластер Г.К.; Грин, Дэррил Р.Х.; Хоукс, Джеффри А.; Хепберн, Лора; Иларио, Ана; Ювенн, Верле А.И.; Марш, Ли; Рамирес-Льодра, Ева; Рид, Уильям Д.К.; Ротерман, Кристофер Н.; Свитинг, Кристофер Дж.; Татье, Свен; Цвирглмайер, Катрин (2012). «Открытие новых глубоководных гидротермальных сообществ в Южном океане и последствия для биогеографии» . ПЛОС Биология . 10 (1): –1001234. дои : 10.1371/journal.pbio.1001234 . ISSN 1545-7885 . ПМК 3250512 . ПМИД 22235194 .
- ^ Перейти обратно: а б с д и ж Сигварт, Джулия Д.; Чен, Чонг; Марш, Ли (2017). «Вредна ли разработка морского дна для моллюсков?». Наутилус . 131 (1): 43–49.
- ^ Перейти обратно: а б Хасимото, Джун; Гамо, Тоситака; Ямагути, Цучида, Синдзи; Ватабе, Хадзиме; Яманака, Китазава, Мицуко (2001) . 10.2108 / . : 18 717–721 (5 ) : zsj.18.717 . S2CID 85582014 doi
- ^ Перейти обратно: а б с Чен, Чонг; Копли, Джонатан Т.; Линсе, Катрин; Роджерс, Алекс Д. (2015). «Низкая связь между популяциями чешуйчатоногих брюхоногих моллюсков (Mollusca: Peltospiridae) в гидротермальных жерлах на Юго-Западном Индийском хребте и Центрально-Индийском хребте». Разнообразие и эволюция организмов . 15 (4): 663–670. дои : 10.1007/s13127-015-0224-8 . S2CID 18521717 .
- ^ Пикрелл, Джон (7 ноября 2003 г.). «Бронированная улитка обнаружена в глубоком море» . Национальные географические новости. Архивировано из оригинала 10 ноября 2003 года . Проверено 16 июля 2016 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л Чен, Чонг; Уэмацу, Кацуюки; Линсе, Катрин; Сигварт, Джулия Д. (2017). «Более чем одним способом: быстрая конвергенция в гидротермальных источниках, показанная трехмерной анатомической реконструкцией Gigantopelta (Mollusca: Neomphalina)» . Эволюционная биология BMC . 17 (1): 62. Бибкод : 2017BMCEE..17...62C . дои : 10.1186/s12862-017-0917-z . ISSN 1471-2148 . ПМК 5333402 . ПМИД 28249568 .
- ^ Сузуки, Йохей; Копп, Роберт Э.; Коруге, Тошихиро; Суга, Акинобу; Такай, Кен; Цучида, Синдзи; Одзаки, Нориаки; Эндо, Казуёси; Хашимото, Джун; Като, Ясухиро; Мизота, Титоши; Хирата, Такафуми; Тиба, Хитоши; Нилсон, Кеннет Х.; Хорикоши, Коки; Киршвинк, Джозеф Л. (2006). «Образование склерита в гидротермальных жерлах брюхоногих моллюсков с чешуйчатыми ногами - возможный контроль биоминерализации сульфида железа животным» (PDF) . Письма о Земле и планетологии . 242 (1–2): 39–50. Бибкод : 2006E&PSL.242...39S . дои : 10.1016/j.epsl.2005.11.029 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м Гоффреди, Шана К.; Варен, Андерс; Сирота, Виктория Дж.; Довер, Синди Л. Ван; Вриенгук, Роберт К. (5 мая 2004 г.). «Новые формы структурной интеграции между микробами и брюхоногими моллюсками из гидротермальных источников Индийского океана» . Прикладная и экологическая микробиология . 70 (5): 3082–3090. Бибкод : 2004ApEnM..70.3082G . дои : 10.1128/АЕМ.70.5.3082-3090.2004 . ПМК 404406 . PMID 15128570 .
- ^ Перейти обратно: а б с д Яо, Хаймин; Дао, Мин; Имхольт, Тимоти; Хуанг, Джейми; Уилер, Кевин; Бонилья, Алехандро; Суреш, Субра; Ортис, Кристина (2010). «Защитные механизмы железной брони глубоководного брюхоногого моллюска гидротермальных источников» . ПНАС . 107 (3): 987–992. Бибкод : 2010PNAS..107..987Y . дои : 10.1073/pnas.0912988107 . ПМК 2808221 . ПМИД 20133823 .
- ^ «Железные доспехи улитки на глазах у военных» . Новости Си-Би-Си. 19 января 2010 г. Проверено 16 июля 2016 г.
- ^ Перейти обратно: а б с Накагава, Сигэру, Ёсихиро; Мураками, Сюн-ичи; Фуджиёси, Саяка; Маэда, Макита, Хироко; Нисимура, Шин-Ичиро; Вацуджи, Томо-о; Кен (2014). «Союзник с панцирными улитками: полный геном гаммапротеобактериального эндосимбионта» . Журнал ISME 8 ( 1): 40–51. Бибкод : 2014ISMEJ...8... 40N doi : 10.1038 PMC 3869010. / ismej.2013.131 PMID 23924784 .
- ^ Дистел, Дэниел Л.; Альтамия, Марвин А.; Линь, Чжэньцзянь; Шипвей, Дж. Рубен; Хан, Эндрю; Фортеза, Имельда; Антемано, Ровена; Лимбако, Ма Гвен Дж. Пеньяфлор; Тебо, Элисон Г.; Дешавес, Ранде; Альбано, Джули; Розенберг, Гэри; Консепсьон, Гизела П.; Шмидт, Эрик В.; Хейгуд, Марго Г. (17 апреля 2017 г.). «Открытие хемоавтотрофного симбиоза у гигантского корабельного червя Kuphus polythalamia (Bivalvia: Teredinidae) расширяет теорию деревянных ступенек» . Труды Национальной академии наук . 114 (18): Е3652–Е3658. Бибкод : 2017PNAS..114E3652D . дои : 10.1073/pnas.1620470114 . ISSN 1091-6490 . ПМЦ 5422788 . ПМИД 28416684 .
- ^ Перейти обратно: а б Накамура, Кентаро; Такай, Кен (2015). «Гидротермальные системы Индийского океана: гидротермальная активность морского дна, физические и химические характеристики гидротермальных флюидов и биологические сообщества, связанные с жерлами». В Исибаши Ж.-и.; и др. (ред.). Подводная биосфера, связанная с гидротермальными системами . Спрингер, Токио. стр. 147–161. дои : 10.1007/978-4-431-54865-2_12 . ISBN 9784431548645 .
- ^ Перейти обратно: а б с Довер, Синди Ван (2002). «Трофические взаимоотношения беспозвоночных на гидротермальном жерлом Кайрей (Центрально-Индийский хребет)». Морская биология . 141 (4): 761–772. Бибкод : 2002МарБи.141..761В . дои : 10.1007/s00227-002-0865-y . S2CID 189819863 .
- ^ Перейти обратно: а б с д «Обнаружена обширная популяция «редкого» чешуйчатого брюхоногого моллюска» . Японское агентство по морским наукам и технологиям о Земле , Университет Хоккайдо , Аквариум Эносима . 30 ноября 2009 года . Проверено 16 июля 2016 г.
- ^ Перейти обратно: а б с Ватанабэ, Хироми; Бидесси, Гириш (2015). «Вентфауна Центрально-Индийского хребта». В Исибаши Ж.-и.; и др. (ред.). Подводная биосфера, связанная с гидротермальными системами . Спрингер, Токио. стр. 205–212. дои : 10.1007/978-4-431-54865-2_16 . ISBN 9784431548645 . S2CID 127157740 .
- ^ Джонсон, Шеннон Б.; Варен, Андерс; Танниклифф, Верена; Довер, Синди Ван; Пшеница, К. Джеффри; Шульц, Томас Ф.; Вриенгук, Роберт К. (04 мая 2015 г.). «Молекулярная систематика и наименование пяти загадочных видов улиток Alviniconcha (Gastropoda: Abyssochrysoidea) из гидротермальных источников» . Систематика и биоразнообразие . 13 (3): 278–295. Бибкод : 2015SyBio..13..278J . дои : 10.1080/14772000.2014.970673 . ISSN 1477-2000 . S2CID 85253352 .
- ^ Перейти обратно: а б с Окутани, Такаши; Хасимото, Джун; Сасаки, Такенори (2004). «Новые таксоны брюхоногих моллюсков из гидротермального источника (поле Кайрей) в центральной части Индийского океана» (PDF) . Венера . 63 (1–2): 1–10. Архивировано из оригинала (PDF) 4 октября 2013 г.
- ^ Перейти обратно: а б Копли, Дж. Т.; Марш, Л.; Гловер, АГ; Хюнербах, В.; Най, В.Е.; Рид, WDK; Свитинг, CJ; Уигэм, Б.Д.; Виклунд, Х. (2016). «Экология и биогеография мегафауны и макрофауны первых известных глубоководных гидротермальных источников на ультрамедленно спрединговом Юго-Западном Индийском хребте» . Научные отчеты . 6 : 39158. Бибкод : 2016NatSR...639158C . дои : 10.1038/srep39158 . ISSN 2045-2322 . ПМК 5155287 . ПМИД 27966649 .
- ^ Сигварт, Джулия Д. (зима 2017 г.). «Глубоководная охрана и« чешуйчатоногие брюхоногие моллюски » » (PDF) . Щупальца . 25 : 39–40.
Внешние ссылки
[ редактировать ] СМИ, связанные с Chrysomallon sqamiferum, на Викискладе?
СМИ, связанные с Chrysomallon sqamiferum, на Викискладе?