Клональный отбор
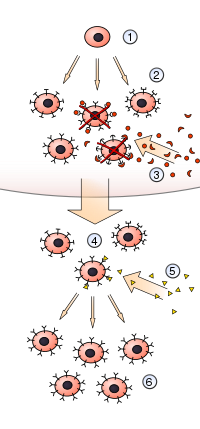
1) Гемопоэтическая стволовая клетка подвергается дифференцировке и генетической перестройке, в результате чего образуется
2) незрелые лимфоциты со множеством различных антигенных рецепторов. Те, которые привязываются к
3) антигены из собственных тканей организма разрушаются, а остальные созревают в
4) неактивные лимфоциты. Большинство из них никогда не встречают совпадающих
5) чужеродный антиген, но те, что есть, активируются и продуцируют
6) множество клонов самих себя.
В иммунологии ) в ответ на вторжение в теория клональной селекции объясняет функции клеток иммунной системы ( лимфоцитов специфических антигенов организм . Эта концепция была введена австралийским врачом Фрэнком Макфарлейном Бёрнетом в 1957 году в попытке объяснить большое разнообразие антител, образующихся во время инициации иммунного ответа . [1] [2] Эта теория стала широко принятой моделью того, как иммунная система человека реагирует на инфекцию и как определенные типы В- и Т-лимфоцитов отбираются для разрушения специфических антигенов . [3]
Теория утверждает, что в уже существующей группе лимфоцитов (как В-, так и Т-клеток) специфический антиген активирует (т.е. отбирает) только свою контрспецифическую клетку, которая затем побуждает эту конкретную клетку размножаться, производя идентичные клоны для производства антител. . Эта активация происходит во вторичных лимфоидных органах, таких как селезенка и лимфатические узлы . [4] Короче говоря, теория представляет собой объяснение механизма возникновения разнообразия специфичности антител. [5] Первые экспериментальные данные были получены в 1958 году, когда Густав Носсаль и Джошуа Ледерберг показали, что одна В-клетка всегда производит только одно антитело. [6] Эта идея оказалась основой молекулярной иммунологии, особенно адаптивного иммунитета . [7]
Постулаты
[ редактировать ]Теорию клонального отбора можно резюмировать следующими четырьмя принципами:
- Каждый лимфоцит несет один тип рецептора с уникальной специфичностью (генерируемый рекомбинацией V(D)J ).
- Занятие рецептора необходимо для активации клеток.
- Дифференцированные эффекторные клетки, происходящие из активированных лимфоцитов, несут рецепторы идентичной специфичности, что и родительская клетка.
- Лимфоциты, несущие рецепторы для собственных молекул (т.е. эндогенных антигенов, вырабатываемых в организме), разрушаются на ранней стадии.
Ранние работы
[ редактировать ]В 1900 году Пауль Эрлих предложил так называемую «теорию боковой цепи» производства антител. Согласно ему, определенные клетки имеют на своей поверхности разные « боковые цепи » (т.е. мембраносвязанные антитела), способные реагировать с разными антигенами. Когда антиген присутствует, он связывается с соответствующей боковой цепью. Затем клетка перестает производить все остальные боковые цепи и начинает интенсивный синтез и секрецию антигенсвязывающей боковой цепи в виде растворимого антитела. Хотя идея Эрлиха отличалась от клонального отбора, она представляла собой теорию отбора, гораздо более точную, чем поучительные теории, которые доминировали в иммунологии в последующие десятилетия.
В 1955 году датский иммунолог Нильс Йерне выдвинул гипотезу о том, что в сыворотке до любого заражения уже имеется огромное количество растворимых антител. Попадание антигена в организм приводит к выделению только одного типа антител, соответствующего ему. Предположительно, это происходит за счет того, что определенные клетки фагоцитируют иммунные комплексы и каким-то образом копируют структуру антитела, чтобы производить его в большем количестве. [8]
В 1957 году Дэвид В. Талмейдж выдвинул гипотезу, что антигены связываются с антителами на поверхности клеток, продуцирующих антитела, и «для размножения отбираются только те клетки, синтезированный продукт которых имеет сродство к антигену». Ключевое отличие от теории Эрлиха заключалось в том, что предполагалось, что каждая клетка синтезирует только один тип антител. После связывания антигена клетка размножается, образуя клоны с идентичными антителами.
Теория клонального отбора Бёрнета
[ редактировать ]Позже, в 1957 году, австралийский иммунолог Фрэнк Макфарлейн Бёрнет статью под названием «Модификация теории Джерна о производстве антител с использованием концепции клональной селекции» опубликовал в довольно малоизвестном Австралийском научном журнале . В ней Бернет развил идеи Талмейджа и назвал получившуюся теорию «теорией клонального отбора». Далее он формализовал эту теорию в своей книге 1959 года «Теория приобретенного иммунитета с помощью клонального отбора» . Он объяснил иммунологическую память клонированием двух типов лимфоцитов. Один клон действует немедленно, борясь с инфекцией, тогда как другой действует дольше, оставаясь в иммунной системе в течение длительного времени и вызывая иммунитет к этому антигену. Согласно гипотезе Бернета, среди антител есть молекулы, которые, вероятно, могут с той или иной степенью точности соответствовать всем или практически всем антигенным детерминантам , встречающимся в биологическом материале, отличным от тех, которые характерны для самого организма. Каждый тип паттерна представляет собой специфический продукт клона лимфоцитов, и суть гипотезы состоит в том, что каждая клетка автоматически имеет на своей поверхности репрезентативные реактивные сайты, эквивалентные таковым у лимфоцитов. глобулин, который они производят. Когда антиген попадает в кровь или тканевые жидкости, предполагается, что он прикрепляется к поверхности любого лимфоцита, несущего реактивные участки, соответствующие одной из его антигенных детерминант. Затем клетка активируется и подвергается пролиферации, производя множество потомков. Таким образом, инициируется преимущественная пролиферация всех тех клонов, чьи реактивные сайты соответствуют антигенным детерминантам антигенов, присутствующих в организме. Потомки способны к активному высвобождению растворимых антител и лимфоцитов и выполняют те же функции, что и родительские формы. [5] [9]
В 1958 году Густав Носсаль и Джошуа Ледерберг показали, что одна В-клетка всегда производит только одно антитело, что стало первым прямым доказательством, подтверждающим теорию клонального отбора. [6]
Теории, поддерживаемые клональным отбором
[ редактировать ]Бёрнет и Питер Медавар вместе работали над пониманием иммунологической толерантности — феномена, который также объясняется клональной селекцией. Это способность организма переносить введение клеток до развития иммунного ответа, если это происходит на ранних стадиях развития организма. В иммунной системе встречается огромное количество лимфоцитов: от клеток, которые толерантны к собственным тканям, до клеток, которые этого не делают. Однако только клетки, толерантные к собственной ткани, выживают на эмбриональной стадии. Если вводится чужая ткань, развиваются лимфоциты, которые включают чужие ткани в качестве собственной ткани.
В 1959 году Бернет предположил, что при определенных обстоятельствах ткани можно успешно трансплантировать иностранным реципиентам. Эта работа привела к гораздо лучшему пониманию иммунной системы, а также к большим достижениям в трансплантации тканей. Бёрнет и Медавар разделили Нобелевскую премию по физиологии и медицине в 1960 году.
В 1974 году Нильс Кай Джерне предположил, что иммунная система функционирует как сеть, регулируемая посредством взаимодействия между различными частями лимфоцитов и секретируемыми ими молекулами. Теория иммунной сети прочно основана на концепции клонального отбора. Джерн получил Нобелевскую премию по физиологии и медицине в 1984 году, главным образом за вклад в теорию иммунных сетей.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Бернет, FM (1976). «Модификация теории производства антител Джерна с использованием концепции клональной селекции» . CA: Журнал рака для врачей . 26 (2): 119–21. дои : 10.3322/canjclin.26.2.119 . ПМИД 816431 . S2CID 40609269 .
- ^ Кон, Мелвин; Ав Митчисон, Н.; Пол, Уильям Э.; Сильверстайн, Артур М.; Талмейдж, Дэвид В.; Вейгерт, Мартин (2007). «Размышления о теории клонального отбора». Обзоры природы Иммунология . 7 (10): 823–830. дои : 10.1038/nri2177 . ПМИД 17893695 . S2CID 24741671 .
- ^ Раевский, Клаус (1996). «Клональная селекция и обучение в системе антител». Природа . 381 (6585): 751–758. Бибкод : 1996Natur.381..751R . дои : 10.1038/381751a0 . ISSN 0028-0836 . ПМИД 8657279 . S2CID 4279640 .
- ^ Мерфи, Кеннет (2012). Иммунобиология Джейнвей, 8-е издание . Нью-Йорк, штат Нью-Йорк: Garland Science. ISBN 9780815342434 .
- ^ Jump up to: а б Джордан, Маргарет А; Бакстер, Алан Дж. (2007). «Количественный и качественный подходы к БОГУ: первые 10 лет теории клонального отбора». Иммунология и клеточная биология . 86 (1): 72–79. дои : 10.1038/sj.icb.7100140 . ПМИД 18040281 . S2CID 19122290 .
- ^ Jump up to: а б Носсаль, GJV; Ледерберг, Джошуа (1958). «Продуцирование антител отдельными клетками» . Природа . 181 (4620): 1419–1420. Бибкод : 1958Natur.181.1419N . дои : 10.1038/1811419a0 . ПМК 2082245 . ПМИД 13552693 .
- ^ Меджитов Р. (2013). «Теория распознавания образов и появление современного врожденного иммунитета» . Журнал иммунологии . 191 (9): 4473–4474. doi : 10.4049/jimmunol.1302427 . ПМИД 24141853 .
- ^ Бернет, FM (1976). «Модификация теории производства антител Джерна с использованием концепции клонального отбора» . CA: Журнал рака для врачей . 26 (2): 119–121. дои : 10.3322/canjclin.26.2.119 . ISSN 0007-9235 . ПМИД 816431 . S2CID 40609269 .
- ^ Ходжкин, Филип Д.; Хит, Уильям Р.; Бакстер, Алан Дж. (2007). «Теория клонального отбора: 50 лет после революции». Природная иммунология . 8 (10): 1019–1026. дои : 10.1038/ni1007-1019 . ПМИД 17878907 . S2CID 29935594 .
Дальнейшее чтение
[ редактировать ]- Подольский, Альфред И. Таубер; Скотт Х. (1997). Поколение разнообразия: теория клональной селекции и развитие молекулярной иммунологии (1-е изд. в мягкой обложке). Кембридж, Массачусетс: Гарвардский университет. Нажимать. ISBN 0-674-00182-6 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - «Биология в контексте – спектр жизни», авторы Питер Обюссон, Эйлин Кеннеди.
- Форсдайк Д.Р. (1995). «Истоки теории иммунитета с клональной селекцией» . Журнал ФАСЭБ . 9 (2): 164–66. дои : 10.1096/fasebj.9.2.7781918 . ПМИД 7781918 . S2CID 38467403 .
Внешние ссылки
[ редактировать ]- Анимация клонального отбора. Архивировано 6 июля 2011 года в Wayback Machine института Уолтера и Элизы Холл.