Хартебист
| Хартебист | |
|---|---|

| |
| Hartebeest от Coke в Национальном парке Серенгети , Танзания. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Парнокопытные |
| Семья: | Bovidae |
| Подсемейство: | Алселафин |
| Род: | Алцелаф Бленвиль , 1816 г. |
| Разновидность: | А. буселафус
|
| Биномиальное имя | |
| Алцелафус буселафус ( «Паллада» , 1766 г.)
| |
| Подвиды [ 2 ] | |
|
Список | |
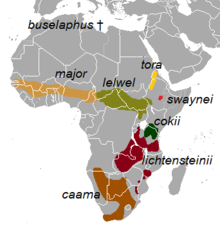
| |
| Распространение подвида | |
| Синонимы [ 2 ] | |
| |

Хартебист ( / ˈ h ɑːr t ə ˌ b iː s t / ; [ 3 ] Alcelaphus buselaphus ), также известный как конгони или каама , — африканская антилопа . Это единственный представитель рода Alcelaphus . восемь подвидов Описано , в том числе два, которые иногда считаются самостоятельными видами. Крупная антилопа, хартбест, имеет высоту чуть более 1 м (3 фута 3 дюйма) в холке, а типичная длина головы и тела составляет от 200 до 250 см (от 79 до 98 дюймов). Вес колеблется от 100 до 200 кг (от 220 до 440 фунтов). странной формы У него особенно удлиненный лоб и рога , короткая шея и заостренные уши. Его ноги, часто имеющие черные отметины, необычайно длинные. Шерсть обычно короткая и блестящая. Цвет шерсти варьируется в зависимости от подвида: от песочно-коричневого у западного бута-хартеба до шоколадно-коричневого у буста Суэйна . Рога есть у обоих полов всех подвидов, причем у самок они более тонкие. Рога могут достигать длины 45–70 см (18–28 дюймов). Помимо длинного лица, большая грудь и резко покатая спина отличают хартебеста от других антилоп. Заметный горб над плечами обусловлен длинной спинной частью. отростков позвонков в этой области. [ 4 ]
Стайные животные, хартебы образуют стада от 20 до 300 особей. Они очень бдительны и неагрессивны. В основном это травоядные животные , их рацион состоит в основном из трав. Спаривание у хартебеста происходит в течение года с одним или двумя пиками и зависит от подвида и местных факторов. И самцы, и самки достигают половой зрелости в возрасте от одного до двух лет. Беременность длится восемь-девять месяцев, после чего рождается один теленок. Пик рождаемости обычно приходится на засушливый сезон. Продолжительность жизни от 12 до 15 лет.
Обитая в сухих саваннах и лесистых лугах, хартебы часто после дождя перебираются в более засушливые места. Сообщалось о них с высоты на горе Кения до 4000 м (13 000 футов). Раньше хартебы были широко распространены в Африке, но их популяция резко сократилась из-за разрушения среды обитания , охоты, поселений людей и конкуренции с домашним скотом за пищу. Каждый из восьми подвидов хартебеста имеет разный природоохранный статус. бубала . (МСОП) объявил вымершим В 1994 году Международный союз охраны природы В то время как популяция красного зверя растет, популяция вида Тора , уже находящегося под угрозой исчезновения , сокращается. Хартебист вымер в Алжире, Египте, Лесото, Ливии, Марокко, Сомали и Тунисе; но был завезен в Эсватини и Зимбабве . Это популярное охотничье животное из-за его высоко ценимого мяса.
Этимология
[ редактировать ]Народное название «hartebeest», возможно, произошло от устаревшего голландского слова «hertebeest » . [ 5 ] буквально олень-зверь , [ 3 ] основано на сходстве (с ранними голландскими поселенцами ) антилопы с оленями . [ 6 ] Первое использование слова «hartebeest» в южноафриканской литературе было в голландского колониального администратора Яна ван Рибека журнале Daghregister в 1660 году. Он писал: « Meester Pieter ein hart-beest geschooten Hadde (Мастер Питер [ван Меерхофф] застрелил одного хартебист)». [ 7 ] Другое название хартебиста — конгони . [ 8 ] слово суахили . [ 9 ] Конгони часто используется, в частности, для обозначения одного из его подвидов — хартебиста кока-колы . [ 10 ]
Таксономия
[ редактировать ]хартебеста Научное название — Alcelaphus buselaphus . Впервые описанный немецким зоологом Петером Симоном Палласом в 1766 году, он отнесен к роду Alcelaphus и помещен в семейство Bovidae . [ 2 ] В 1979 году палеонтолог Элизабет Врба поддержала Sigmoceros как отдельный род антилопы Лихтенштейна , разновидности антилопы, поскольку она предположила, что он связан с Connochaetes (антола гну). [ 11 ] [ 12 ] Она проанализировала черты черепа ныне живущих и вымерших видов антилоп, чтобы составить кладограмму , и утверждала, что широкий череп связывает антилопу Лихтенштейна с Connochaetes . [ 13 ] Однако это открытие не было повторено Аланом В. Джентри из Музея естественной истории , который классифицировал его как независимый вид Alcelaphus . [ 14 ] Зоологи, такие как Джонатан Кингдон и Теодор Халтенорт, считали его подвидом A. buselaphus . [ 2 ] Врба распустил новый род в 1997 году после пересмотра. [ 15 ] Анализ мтДНК не смог найти никаких доказательств в пользу выделения отдельного рода антилоп Лихтенштейна. Это также показало, что триба Alcelaphini является монофилетической , и обнаружило близкое родство между Alcelaphus и сассабиями (род Damaliscus ) — как генетически, так и морфологически. [ 16 ]
Подвиды
[ редактировать ]
Выявлено восемь подвидов, из них два – A. b. чаама и А.б. lichtensteinii – считаются самостоятельными видами. Однако генетическое исследование 1999 года, проведенное П. Арктандером из Копенгагенского университета и его коллегами, в ходе которого были взяты образцы контрольной области митохондриальной ДНК , показало, что эти двое образуют кладу внутри A. buselaphus , и что признание их как видов приведет к тому, что A. buselaphus парафилетический (неестественная группировка). В том же исследовании было обнаружено, что A.b. major является наиболее расходящимся, поскольку он разветвился до того, как линия разделилась, чтобы дать объединенную линию caama/lichtensteinii и другую, которая дала начало оставшимся существующим подвидам. [ 17 ] И наоборот, филогенетическое исследование 2001 года, основанное на анализе D-петли и цитохрома b, проведенное Эйстейном Флагстадом (из Норвежского института исследований природы, Тронхейм ) и его коллегами, показало, что южная линия A. b. caama и A. lichtensteinii разошлись раньше всех. [ 12 ] Анализ строения черепа позволяет разделить его на три основных отдела: A. b. buselaphus подразделение ( номинальное , включая A. b. major Division), A. b. подразделение tora (также включающее A.b.cokii и A.b.swaynei ) и A.b. левельское подразделение. [ 2 ] Другой анализ данных последовательности цитохрома b и D-петли показывает заметное сродство между A. b. лельвель и А.б. дивизии Тора . [ 18 ]
Восемь подвидов, включая два спорных: [ 1 ] [ 19 ]
- † А.б. buselaphus (Pallas, 1766) : известный как бубал-хартебест или северный хартебест. Раньше встречался по всей Северной Африке, от Марокко до Египта . Истреблен к 1920-м годам. [ 20 ] (МСОП) объявил его вымершим В 1994 году Международный союз охраны природы и природных ресурсов . [ 21 ] [ 22 ]
- А. б. каама ( Сен-Илер , 1803 г.) : известный как красный хартебист или капский хартебист. Ранее наблюдался на юге Анголы ; северные и восточные саванны Намибии ; центральная, южная и юго-западная Ботсвана ; Северный Кейп , Восточный Кейп , Западный Кейп , Фри-Стейт , Северо-Западная и Гаутенг провинции , а также западный Квазулу-Натал в Южной Африке. В настоящее время ликвидирован во всех этих районах, за исключением Северного Кейпа, центральной и юго-западной Ботсваны и Намибии. В этих странах произошла крупная реинтродукция. [ 20 ] Популяция этого зверя растет. [ 23 ]
- А. б. cokii Günther , 1884 : известен как хартебист кока-колы или конгони. Родом из и северной Танзании и обитает только на их территории. Кении [ 20 ]
- А. б. Лельвель (Хьюглин, 1877) : известный как хартебист Лелвеля . Ранее встречался на севере и северо-востоке Демократической Республики Конго ; юго-восточный и юго-западный Судан ; и северо-западная оконечность Танзании . [ 20 ] Резкое сокращение численности населения с 1980-х годов привело к тому, что большинство особей оказались на охраняемых территориях внутри и за пределами своего ареала. [ 24 ]
- А. б. lichtensteinii ( Peters , 1849) : известный как хартенштейн Лихтенштейна . Он населяет леса Миомбо в Восточной и Южной Африке. [ 25 ] Он произрастает в Анголе, Демократической Республике Конго, Малави , Мозамбике , Южной Африке, Танзании, Замбии и Зимбабве . [ 26 ]
- А. б. майор ( Блит , 1869) : известный как западный хартебист . Ранее широко встречался в Мали, Нигере , Сенегале , Гамбии , Гвинее-Бисау , Гвинее , Кот-д'Ивуаре , Гане , Нигерии , юго-западном Чаде, Камеруне, западной части Центральноафриканской Республики и Бенине . В настоящее время он встречается в гораздо меньших количествах, главным образом на охраняемых территориях этих стран. Вероятно, он вымер в Гамбии. [ 20 ]
- А. б. swaynei ( Sclater , 1892) : известный как хартебист Суэйна . Ограничено южной частью Рифтовой долины в Эфиопии, подтверждено и внесено в список эндемичных видов. Раньше он встречался по всей Рифтовой долине, а его ареал простирался на восток до северо-запада Сомали . Он исчез из Сомали к 1930 году из-за эпидемии чумы крупного рогатого скота, незаконного браконьерства и чрезмерного выпаса скота. [ 20 ] Подтверждено как вид, находящийся под угрозой исчезновения, его популяция невелика, насчитывает не менее 1500 особей, и находится на грани сокращения. [ 27 ] [ 28 ]
- А. б. Тора Грей , 1873 : Известен как Хартебист. Тора Ранее встречался на северо-западе Эфиопии, а также на западе и Эритреи. юго-западе [ 29 ] Его нынешний статус неясен, хотя местные жители сообщают о небольшом количестве жителей этих районов. [ 20 ]
-
 Бубал Хартебист
Бубал Хартебист -
 Сохранить
Сохранить -
 любимец кока-колы
любимец кока-колы -
 Любимая Лихтенштейна
Любимая Лихтенштейна -
 любимец Суэйна
любимец Суэйна





Генетика и гибриды
[ редактировать ]
А. б. Суэйни , Святилище урожая Суэйна , Эфиопия
В 2000 году в ходе исследования были тщательно изучены две основные популяции антилопы Суэйна из заповедника дикой природы Сенкеле и национального парка Нечисар на предмет митохондриальной (D-петли) и ядерной ( микросателлитной ) изменчивости в попытке оценить уровни генетической изменчивости между популяциях и внутри подвидов. Результаты показали значительную разницу между двумя популяциями; тот, что из заповедника дикой природы Сенкеле, показал большее генетическое разнообразие , чем тот, что из национального парка Нечисар. Еще одним открытием стало то, что переселение особей из заповедника Сенкеле в 1974 году не внесло существенного вклада в генофонд национального парка Нечисар. Кроме того, популяции бульдогов Суэйна сравнили с большой популяцией красных антилоп, и было обнаружено, что оба подвида имеют высокую степень генетической изменчивости. Исследование пропагандировало сохранение in situ зверя Суэйна и новую попытку его перемещения с целью сохранения генетического разнообразия и увеличения его популяции на обеих охраняемых территориях. [ 18 ]
Диплоидное . число хромосом у антилопы составляет 40. Гибриды обычно встречаются из районов, где ареалы двух подвидов перекрываются [ 8 ] Гибриды между зверями Лелвель и Тора были зарегистрированы в восточном Судане и западной Эфиопии, на участке к югу от Голубого Нила примерно до 9° северной широты. [ 30 ] Исследование доказало, что самец гибрида красного була и блесбока ( Damaliscus pygargus ) бесплоден . Стерильность гибрида объяснялась трудностями сегрегации во время мейоза , о чем свидетельствует азооспермия и малое количество половых клеток в семенных канальцах . [ 31 ]

Между подвидами существует три четко выраженных гибрида:
- Alcelaphus lelwel x cokii : Кенийский горный хартебист или Лайкипийский хартебист. Это нечто среднее между Lelwel и Hartebeest от Coke. [ 32 ] Этот гибрид светлее по цвету и крупнее, чем хартебист Кока-Колы. Это светло-желтый цвет с красновато-коричневой верхней частью, а голова длиннее, чем у кока-колы. У обоих полов есть рога, которые тяжелее и длиннее, чем у родителей. Раньше он был распространен по всей западной части Кении, между озером Виктория и горой Кения , но теперь считается, что его распространение ограничено долиной Ламбве (юго-запад Кении) и Лайкипией , а также близлежащими регионами западно-центральной Кении. [ 33 ] [ 34 ]
- У хартебеста Джексона нет четкого таксономического статуса. Как и приведенная выше форма, он считается гибридом хартебеста Лелвеля и кока-колы и имеет аналогичное распространение. В базе данных африканских антилоп (1998) он рассматривается как синоним хартебеста Лельвель. [ 20 ] От озера Баринго до горы Кения хартебест Джексона значительно напоминает хартебест Лелвела, тогда как от озера Виктория до южной части Рифт-Валли он больше похож на хартебеста Кока-Колы. [ 35 ]
- Alcelaphus lelwel x swaynei : также известен как хартебест Неймана, названный в честь путешественника и охотника Артура Генри Неймана . [ 35 ] Это считается помесью хартбеста Лелвела и хартбеста Суэйна. [ 32 ] Морда длиннее, чем у хартебеста Суэйна. Цвет шерсти золотисто-коричневый, к нижней части тела светлее. Подбородок имеет оттенок черного, а хвост заканчивается черным пучком. У обоих полов рога длиннее, чем у буста Суэйна. Рога растут в форме широкой буквы «V», в отличие от широкой скобки у биста Суэйна и узкой буквы «V» у биста Лелвела, загибающихся назад и слегка внутрь. Встречается в Эфиопии, на небольшой территории к востоку от реки Омо и к северу от озера Туркана , простираясь к северо-востоку от озера Чу Бахир до озера Чамо . [ 36 ]
Эволюция
[ редактировать ]Род Alcelaphus возник около 4,4 миллиона лет назад в кладе , членами которой были Damalops , Numidocapra , Rabaticeras , Megalotragus , Oreonagor и Connochaetes . Анализ с использованием филогеографических закономерностей в популяциях хартебе предположил возможное происхождение Alcelaphus в Восточной Африке. [ 37 ] Alcelaphus быстро распространился по африканским саваннам, вытеснив несколько предыдущих форм (например, родственника хиролы ) . Флагстад и его коллеги показали раннее разделение популяций антилоп на две отдельные линии около 0,5 миллиона лет назад — одну к северу, а другую к югу от экватора. Северная линия далее разделилась на восточную и западную линии почти 0,4 миллиона лет назад, скорее всего, в результате расширения пояса тропических лесов Центральной Африки и последующего сокращения среды обитания в саванне в период глобального потепления. Восточная линия дала начало хартебистам Кока-Колы, Суэйна, Торы и Лелвеля; а от западной линии произошли бубалы и западные хартбесты. Южная линия дала начало булу Лихтенштейна и красному хартебусту. Эти два таксона филогенетически близки, разошлись всего 0,2 миллиона лет назад. Исследование пришло к выводу, что эти основные события на протяжении всей эволюции хартебеста тесно связаны с климатическими факторами и что имели место последовательные всплески радиации от более постоянной популяции - рефугиум — в Восточной Африке; это может иметь жизненно важное значение для понимания истории эволюции не только хартебеста, но и других млекопитающих африканской саванны. [ 12 ]
Самая ранняя летопись окаменелостей датируется почти 0,7 миллиона лет назад. [ 8 ] Окаменелости красного хартебеста были найдены в Эландсфонтейне, Корнелии (Свободное государство) и Флорисбаде в Южной Африке , а также в Кабве в Замбии . [ 38 ] В Израиле останки антилопы были найдены в северном Негеве , Шефеле , Шаронской равнине и Тель-Лахише . Эта популяция хартебеста первоначально была ограничена открытой местностью самых южных регионов южного Леванта . Вероятно, на него охотились в Египте, что повлияло на численность в Леванте и отдалило его от основной популяции в Африке. [ 39 ]
Описание
[ редактировать ]
Крупная антилопа с особенно удлиненным лбом и рогами странной формы, хартбест имеет высоту чуть более 1 м (3 фута 3 дюйма) в холке, а типичная длина головы и тела составляет от 200 до 250 см (от 79 до 98 дюймов). ). Вес колеблется от 100 до 200 кг (от 220 до 440 фунтов). Хвост длиной от 40 до 60 см (от 16 до 24 дюймов) заканчивается черным пучком. [ 40 ] Другими отличительными особенностями хартебеста являются его длинные ноги (часто с черными отметинами), короткая шея и заостренные уши. [ 41 ] Исследование связало размер видов антилоп с продуктивностью среды обитания и количеством осадков. [ 42 ] Западный хартебист является самым крупным подвидом и имеет характерную белую линию между глазами. [ 43 ] Красный хартбест также крупный, с черным лбом и контрастной светлой полосой между глазами. [ 44 ] У большого зверя Лелвеля на передней части ног темные полосы. [ 30 ] Хартебест Кока-Колы умеренно крупный, с более коротким лбом и более длинным хвостом по сравнению с другими подвидами. [ 45 ] Хартебест Лихтенштейна меньше по размеру, с темными полосами на передней части ног, как и у зверя Лельвеля. [ 46 ] Хартебест Суэйна меньше, чем хартебест Торы, но у обоих более короткий лоб и схожий внешний вид. [ 47 ]
Обычно короткая и блестящая шерсть различается по цвету в зависимости от подвида. [ 48 ] Западный хартбест бледно-песчано-коричневого цвета, но передняя часть ног темнее. [ 43 ] Красный хартебист красновато-коричневый, с темным лицом. Черные отметины можно наблюдать на подбородке, задней части шеи, плечах, бедрах и ногах; они резко контрастируют с широкими белыми пятнами, которые отмечают его бока и нижнюю часть крупа. [ 44 ] [ 49 ] Lelwel Hartebeest имеет красновато-коричневый оттенок. [ 30 ] У кока-колы хартбест верхняя часть тела от красноватой до желтовато-коричневой, но ноги и круп относительно светлее. [ 45 ] Хартебест Лихтенштейна красновато-коричневый, хотя бока более светло-коричневые, а крупа беловатая. [ 46 ] У зверя Тора верхняя часть тела, морда, передние конечности и круп темно-красновато-коричневого цвета, а задние ноги и нижняя часть живота желтовато-белые. [ 29 ] [ 50 ] Хартебист Суэйна имеет насыщенный шоколадно-коричневый цвет с мелкими белыми пятнами, которые на самом деле являются белыми кончиками его волос. Его лицо черное, за исключением шоколадной полосы под глазами. Плечи и верхняя часть ног черные. [ 47 ] Волосы на теле биста имеют тонкую текстуру и имеют длину около 25 мм (1 дюйм). [ 11 ] У антилопы Кока-Колы и Лихтенштейна есть предглазничные железы (железы возле глаз) с центральным протоком, которые выделяют темную липкую жидкость у антилопы Кока-Колы и Лихтенштейна, а также бесцветную жидкость у антилопы Лельвеля. [ 48 ]

Рога есть у обоих полов всех подвидов, причем у самок они более тонкие. Рога могут достигать длины 45–70 см (18–28 дюймов); максимальная длина рупора 74,9 см ( 29 + 1 ⁄ 2 дюйма), зарегистрировано у намибийского красного була. [ 40 ] Рога западного хартбеста толстые, имеют U-образную форму спереди и Z-образную форму с боков, растут сначала назад, а затем вперед, заканчиваясь резким поворотом назад. [ 43 ] Рога красного и лельвельского хартбеста похожи на рога западного хартебеста, но при взгляде спереди кажутся V-образными. [ 30 ] [ 44 ] У була Лихтенштейна толстые рога с параллельными кольцами и плоским основанием. Его рога короче, чем у других подвидов, изгибаются вверх, затем резко вперед, за которым следует поворот внутрь под углом около 45 ° и последний поворот назад. [ 46 ] Рога биста Суэйна тонкие и имеют форму скобок, загибаются вверх, а затем назад. [ 47 ] Рога антилопы Тора особенно тонкие и раскинуты в стороны, расходясь сильнее, чем у любого другого подвида. [ 50 ]
Помимо длинного лица, большая грудь и резко покатая спина отличают хартебеста от других антилоп. [ 5 ] У хартебеста есть несколько общих физических черт с сассаби (род Damaliscus ), такие как удлиненное и узкое лицо, форма рогов, текстура и цвет шерсти , а также конечный пучок хвоста. У антилопы гну более специализированные черты черепа и рогов, чем у антилопы-харте. [ 48 ] У хартебеста наблюдается половой диморфизм , но лишь незначительно, поскольку представители обоих полов имеют рога и одинаковую массу тела. Степень полового диморфизма варьируется в зависимости от подвида. Самцы хартебеста Суэйна и Лихтенштейна тяжелее самок на 8%, а красного хартебеста - на 23%. В одном исследовании самый высокий диморфизм был обнаружен в весе черепа. [ 51 ] Другое исследование пришло к выводу, что продолжительность сезона размножения является хорошим предиктором диморфизма ножки (костных структур, из которых растут рога), высоты и веса черепа, а также лучшим предиктором окружности рогов. [ 52 ]
Экология и поведение
[ редактировать ]Активный в основном в дневное время, хартебест пасется ранним утром и ближе к вечеру, а около полудня отдыхает в тени. Стайный вид, образует стада до 300 особей. Большее количество собирается в местах с обильной травой. В 1963 году на равнине возле Секома-Пан в Ботсване было зарегистрировано скопление из 10 000 животных. [ 48 ] Однако движущиеся стада не столь сплочены и имеют тенденцию часто расходиться. Членов стада можно разделить на четыре группы: территориальные взрослые самцы, нетерриториальные взрослые самцы, молодые самцы и самки с детенышами. Самки образуют группы от пяти до 12 животных, в группе бывает четыре поколения молодняка. Самки борются за господство над стадом. [ 40 ] Спарринги между мужчинами и женщинами являются обычным явлением. [ 8 ] В возрасте трех или четырех лет самцы могут попытаться захватить территорию и ее самок. Местный самец защищает свою территорию и будет сражаться, если его спровоцировать. [ 51 ] Самец отмечает границу своей территории посредством дефекации. [ 40 ]

Hartebeest — чрезвычайно бдительные и осторожные животные с высокоразвитым мозгом . [ 53 ] [ 54 ] Обычно спокойные по своей природе, хартбесты могут стать свирепыми, если их спровоцировать. Во время кормления одна особь следит за опасностью, часто стоя на термитнике, чтобы видеть дальше. В случае опасности все стадо убегает гуськом после того, как особь внезапно тронется с места. [ 40 ] На взрослых зверей охотятся львы, леопарды , гиены и дикие собаки ; гепарды и шакалы охотятся на несовершеннолетних . [ 40 ] Крокодилы также могут охотиться на хартбеста. [ 55 ]
Тонкие длинные ноги хартебеста позволяют быстро сбежать в открытую среду обитания; в случае нападения грозные рога используются, чтобы отогнать хищника. Приподнятое положение глаз позволяет зверюге постоянно осматривать свое окружение, даже когда он пасется. Морда эволюционировала таким образом, чтобы получать максимум питательных веществ даже от скромной диеты. [ 8 ] Рога также используются во время боев между самцами за доминирование в период размножения; [ 52 ] звон рогов настолько громкий, что его можно услышать на расстоянии сотен метров. [ 8 ] Начало боя отмечается серией движений головой и стойками, а также выбрасыванием помета в кучи навоза. Противники падают на колени и, нанеся молотоподобный удар, начинают борьбу, сцепляясь рогами. Один пытается отбросить голову другого в сторону, чтобы нанести удар рогами по шее и плечам. [ 51 ] Драки редко бывают серьезными, но если таковые имеются, они могут оказаться фатальными. [ 48 ]
Как и сассаби, хартбесты издают тихие крякающие и хрюкающие звуки. Молодые особи, как правило, более громкие, чем взрослые, и издают крякающий крик, когда их встревожены или преследуют. [ 40 ] Hartebeest использует дефекацию как обонятельный и визуальный индикатор. [ 48 ] Стада, как правило, ведут оседлый образ жизни и имеют тенденцию мигрировать только при неблагоприятных условиях, таких как стихийные бедствия. [ 56 ] Хартебест является наименее мигрирующим в племени Alcelaphini (в которое также входят антилопы гну и сассаби), а также потребляет наименьшее количество воды и имеет самый низкий уровень метаболизма среди членов племени. [ 48 ]
Паразиты и болезни
[ редактировать ]несколько паразитов . У хартебеста было выделено [ 57 ] [ 58 ] Эти паразиты регулярно чередуются между антилопами и газелями или антилопами гну. [ 59 ] Hartebeest может быть заражен тейлериозом, вызываемым Rhipicephalusevertsi и видами Theileria . [ 60 ] К югу от Сахары распространенными паразитами являются Loewioestrus variolosus , Gedoelstia cristata и G. hassleri . Последние два вида могут вызывать серьезные заболевания, например, энцефалит . [ 61 ] Однако паразиты не всегда вредны — 252 личинки в голове одной замбийской особи обнаружено без какой-либо патогенности . [ 58 ] Нематоды , цестоды , парамфистомы ; и круглый червь Setaria labiatopapillosa также были выделены из антилопы. [ 62 ] [ 63 ] В 1931 году красный антилоп в Гобабисе (юго-западная Африка) был заражен длинными тонкими червями. Они были названы Longistrongylus meyeri в честь их коллекционера Т. Мейера. [ 64 ]

Диета
[ редактировать ]Хартебесты в основном травоядные, и их рацион состоит в основном из трав. [ 65 ] Исследование, проведенное на ранчо Nazinga Game Ranch в Буркина-Фасо, показало, что структура черепа хартебеста облегчает приобретение и пережевывание пищи с высоким содержанием клетчатки. [ 66 ] Hartebeest потребляет гораздо меньше пищи, чем другие представители Alcelaphini. Длинная тонкая морда хартебеста помогает питаться листовыми пластинками короткой травы и отгрызать листовые влагалища со стеблей трав. Кроме того, он может получать питательную пищу даже из высоких старческих трав. Эти приспособления хартебеста позволяют ему хорошо питаться даже в засушливый сезон, который обычно является трудным периодом для травоядных. [ 8 ] Например, по сравнению с чалой антилопой , антилопа лучше добывает и пережевывает скудные отростки многолетних трав в то время, когда корм менее всего доступен. [ 66 ] Эти уникальные способности могли позволить зверю преобладать над другими животными миллионы лет назад, что привело к его успешному распространению по всей Африке. [ 8 ]
Травы обычно составляют не менее 80 процентов рациона хартебеста, но на их долю приходится более 95 процентов их пищи в сезон дождей, с октября по май. Jasminum kerstingii входит в рацион хартебеста в начале сезона дождей. В межсезонье они питаются в основном стеблями трав . [ 66 ] Исследование показало, что антилопа-харте способна переваривать больше пищи, чем топи и антилопа гну. [ 67 ] В районах с нехваткой воды он может выжить за счет дынь, корней и клубней. [ 48 ]
В исследовании избирательности травы среди антилоп гну, зебры и антилопы кока-колы антилопа-харте показала самую высокую избирательность. Все животные предпочитали Themeda triandra Pennisetum mezianum и Digitaria macroblephara . В засушливый сезон съедается больше видов трав, чем в сезон дождей. [ 68 ]
Воспроизведение
[ редактировать ]
Спаривание у хартебеста происходит в течение года, с одним или двумя пиками, на которые может влиять наличие пищи. [ 65 ] И самцы, и самки достигают половой зрелости в возрасте от одного до двух лет. Размножение зависит от подвида и местных факторов. [ 11 ] Спаривание происходит на территориях, охраняемых одним самцом, преимущественно на открытой местности. [ 65 ] Самцы могут яростно бороться за доминирование. [ 51 ] после чего доминирующий самец нюхает гениталии самки и следует за ней, если у нее течка . Иногда самка во время течки слегка вытягивает хвост, сигнализируя о своей восприимчивости. [ 48 ] и самец пытается преградить путь самке. В конце концов она может остановиться и позволить самцу сесть на нее. Копуляция кратковременная и часто повторяется, иногда дважды и более в минуту. [ 48 ] Любой злоумышленник в это время прогоняется. [ 40 ] В больших стадах самки часто спариваются с несколькими самцами. [ 48 ]
Беременность длится восемь-девять месяцев, после чего рождается один теленок весом около 9 кг (20 фунтов). Пик рождаемости обычно приходится на засушливый сезон и происходит в зарослях – в отличие от антилоп гну, которые рожают группами на равнинах. [ 48 ] Хотя телята могут передвигаться самостоятельно вскоре после рождения, обычно они лежат на открытом воздухе в непосредственной близости от своих матерей. [ 32 ] Теленка отнимают от груди в четыре месяца. [ 32 ] но молодые самцы остаются с матерями два с половиной года, дольше, чем у других алцелафинов. [ 48 ] Часто смертность молоди самцов высока, так как им приходится сталкиваться с агрессией территориальных взрослых самцов, а также лишаться ими хорошего корма. [ 40 ] Продолжительность жизни от 12 до 15 лет. [ 65 ]
среда обитания
[ редактировать ]Hartebeest обитает в сухих саваннах, открытых равнинах и лесистых лугах. [ 11 ] часто после дождя переезжают в более засушливые места. Они более терпимы к лесным массивам, чем другие альселафини, и часто встречаются на опушках леса. [ 65 ] Сообщалось о них с высоты на горе Кения до 4000 м (13 000 футов). [ 1 ] Известно, что красный хартбест перемещается по обширным территориям, а самки бродят по домам на расстояние более 1000 км. 2 (390 квадратных миль), с мужскими территориями 200 км 2 (77 квадратных миль) по размеру. [ 69 ] Самки в Национальном парке Найроби (Кения) имеют индивидуальные участки обитания, простирающиеся на 3,7–5,5 км 2 ( 1 + 3 / 8 – 2 + 1 / 8 кв. миль), которые особо не связаны ни с одной женской группой. В среднем женские ареалы достаточно велики, чтобы включать от 20 до 30 мужских территорий. [ 41 ]
Статус и сохранение
[ редактировать ]


Каждый подвид хартебеста внесен в список под своим природоохранным статусом Международного союза охраны природы . Вид в целом классифицируется как вызывающий наименьшее беспокойство . МСОП [ 1 ] Хартебист вымер в Алжире, Египте, Лесото, Ливии, Марокко, Сомали и Тунисе. [ 1 ]
- Бубал -хартебест объявлен вымершим в 1994 году. [ 21 ] Немецкий исследователь Генрих Барт в своих работах 1857 года называет огнестрельное оружие и европейское вторжение среди причин уменьшения его численности. [ 70 ] он вымер в Тунисе . К концу 19 века [ 71 ] Последняя особь была застрелена в Миссуре ( Алжир ) в 1925 году. [ 72 ]
- Hartebeest компании Coke внесен в список вызывающих наименьшее беспокойство. На этот вид сильно повлияло разрушение среды обитания, и сегодня около 42 000 особей кока-колы обитают в Маре , Национальном парке Серенгети , Национальном парке Тарангире в Танзании и Восточном национальном парке Цаво в Кении. Численность населения сокращается, и 70% населения проживает на охраняемых территориях. [ 73 ]
- занесен Хартебест Лелвель в список находящихся под угрозой исчезновения, и его численность значительно сократилась с 1980-х годов, когда его популяция составляла более 285 000 человек. Раньше он был распространен в основном в Центральноафриканской Республике, Эфиопии, северной и северо-восточной Демократической Республике Конго и южном Судане. [ 20 ] Осталось менее 70 000 особей. [ 24 ] Большая часть населения в настоящее время проживает в Чаде, в регионе Саламат и в национальном парке Закума (Чад), население национального парка пользуется улучшенной защитой и наблюдается рост населения с 1980-х годов; Национальный парк Маново-Гунда Сент-Флорис и национальный парк и биосферный заповедник Баминги-Бангоран в Центральноафриканской Республике, популяция которых сокращается; Заповедник Руманьика Оругунду и заповедник Ибанда в Танзании; и национальный парк Мерчисон-Фолс в Уганде . [ 20 ]
- Хартебест Лихтенштейна внесен в список вызывающих наименьшие опасения и встречается на охраняемых территориях, таких как заповедник Селус , а также в дикой природе на юге и западе Танзании и Замбии. [ 26 ]
- Красный хартебист внесен в список видов, вызывающих наименьшее беспокойство. Он является наиболее распространенным, и его численность увеличивается после его реинтродукции на охраняемые и частные территории. он вымер Однако в Лесото с двадцатого века. [ 20 ] Его население оценивается более чем в 130 000 человек (по состоянию на 2008 год). [ 23 ] преимущественно на юге Африки. [ 69 ] В Намибии самая большая популяция встречается в национальном парке Этоша . Реинтродуцированная популяция процветает в заповеднике Малолотья ( Эсватини ), за пределами его ареала. Однако на юго-западе Ботсваны численность резко упала. [ 20 ]
- внесен Хартебест Тора в список находящихся под угрозой исчезновения ; МСОП установил, что по состоянию на 2008 год выжило менее 250 взрослых особей. Возможно, они вымерли в Судане из-за чрезмерной охоты и расширения сельского хозяйства , но могут все еще существовать в меньшем количестве в Эритрее и Эфиопии. [ 74 ] Поступали неподтвержденные сообщения о том, что местные жители видели хартебеста Тора к юго-востоку от национального парка Диндер , откуда он исчез до 1960 года. [ 20 ]
- Хартебист Суэйна занесен в список находящихся под угрозой исчезновения и близок к тому, чтобы находиться под угрозой исчезновения . Общая популяция в 2008 году составляла менее 600 особей, из них половозрелых особей насчитывалось 250. Но в 2021 году популяция медленно увеличилась с 600 до 1528 особей. Он ограничен четырьмя основными охраняемыми территориями: заповедником дикой природы Сенкеле , национальным парком Нечисар , национальным парком Аваш и национальным парком Мейз . [ 75 ] Бисам в Сенкеле приходится конкурировать с домашним скотом народа оромо . [ 27 ] Исследование, проведенное в национальном парке Нечисар в 2009 и 2010 годах, выявило значительное увеличение поголовья оромо (увеличение на 49,9% и 56,5% в 2006 и 2010 годах соответственно), незаконную эксплуатацию ресурсов и утрату среды обитания как основные угрозы для зверей Суэйна. населения там. [ 76 ]
- Западный хартебист внесен в список находящихся под угрозой исчезновения . [ 77 ] Он был уничтожен на большей части своего ареала, включая юго-западные саванны и национальный парк Букль-дю-Бауле в Мали; юго-западный Нигер; южный Сенегал; Гамбия; Кот-д'Ивуар ; Буркина-Фасо . Небольшие популяции сохранились в национальном парке Бафинг и на территории, ограниченной Бамако , Бугуни и Сикассо в Мали; заповедник Таму в Нигере; Национальный парк Ниоколо-Коба в Сенегале; Национальный парк Комоэ в Кот-д'Ивуаре; Лес Дифула и ранчо Назинга в Буркина-Фасо; Национальный парк Пенджари в Бенине; и Буба Нджида , Бенуэ и Фаро в Камеруне. национальные парки [ 20 ]
Отношения с людьми
[ редактировать ]Hartebeest — популярные охотничьи и трофейные животные, поскольку они хорошо заметны и, следовательно, на них легко охотиться. [ 40 ] [ 65 ] Наглядные и эпиграфические свидетельства из Египта позволяют предположить, что в эпоху верхнего палеолита египтяне охотились на зверей и одомашнили их. Хартебист был важным источником мяса. [ 78 ] но его хозяйственное значение было ниже, чем у газелей и других пустынных видов. [ 50 ] Однако с началом эпохи неолита охота стала менее распространенной, и, следовательно, останки хартебеста этого периода в Древнем Египте, где он сейчас вымер, редки. [ 78 ]
В исследовании влияния места и пола на характеристики туши средний вес туши самца красного антилопы составлял 79.3 kg (174+3⁄4 lb) and that of females was 56 kg (123 lb). The meat of the animals from Qua-Qua region had the highest lipid content—1.3 g (20 gr) per 100 г ( 3 + 1 / 2 унции) мяса. Незначительные различия были обнаружены в концентрациях отдельных жирных кислот , аминокислот и минералов . Исследование показало, что мясо булыжника полезно для здоровья, поскольку соотношение полиненасыщенных и насыщенных жирных кислот составляло 0,78, что немного больше рекомендуемого 0,7. [ 79 ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и Группа специалистов МСОП по антилопам SSC (2019 г.) [измененная версия оценки 2016 г.]. « Алцелафус буселафус » . Красный список исчезающих видов МСОП . 2019 : e.T811A143160967. doi : 10.2305/IUCN.UK.2019-1.RLTS.T811A143160967.en . Проверено 18 февраля 2022 г.
- ^ Перейти обратно: а б с д и Уилсон, Делавэр; Ридер, Д.М., ред. (2005). Виды млекопитающих мира: таксономический и географический справочник (3-е изд.). Балтимор, США: Издательство Университета Джонса Хопкинса. п. 674. ИСБН 978-0-8018-8221-0 . OCLC 62265494 . Архивировано из оригинала 28 октября 2020 г. Проверено 5 декабря 2020 г.
- ^ Перейти обратно: а б «Хартебист» . Словарь Merriam-Webster.com . Мерриам-Вебстер . Проверено 24 января 2016 г.
- ^ Кингдон, Джонатан; Хаппольд, Дэвид; Бутынский, Томас; Хоффманн, Майкл; Хаппольд, Мередит; Калина, Ян (23 мая 2013 г.). Млекопитающие Африки . А&С Черный. п. 512. ИСБН 978-1-4081-8996-2 . Архивировано из оригинала 8 марта 2022 г. Проверено 5 февраля 2022 г.
- ^ Перейти обратно: а б Марес, Массачусетс (1999). Энциклопедия пустынь . Норман, США: Университет Оклахомы Пресс. п. 265. ИСБН 978-0-8061-3146-7 .
- ^ Ллевеллин, ЕС (1936). «Влияние южноафриканского голландского языка или африкаанс на английский словарный запас» . Влияние нижнеголландского языка на словарный запас английского языка . Лондон, Великобритания: Издательство Оксфордского университета. п. 163. Архивировано из оригинала 29 апреля 2009 г. Проверено 21 января 2008 г.
- ^ Скиннер, доктор медицинских наук; Чимимба, Коннектикут (2005). Млекопитающие Южноафриканского субрегиона (3-е изд.). Кембридж, Великобритания: Издательство Кембриджского университета. п. 649. ИСБН 978-0-521-84418-5 .
- ^ Перейти обратно: а б с д и ж г час Кингдон, Дж. (2013). Млекопитающие Африки . Лондон, Великобритания: Блумсбери. стр. 510–22. ISBN 978-1-4081-2257-0 .
- ^ «Конгони» . Словарь Merriam-Webster.com . Мерриам-Вебстер . Проверено 26 января 2016 г. .
- ^ Суонк, WG (1971). Африканская антилопа . Нью-Йорк, США: Winchester Press. п. 95. ИСБН 978-0-87691-029-0 .
- ^ Перейти обратно: а б с д Новак, Р.М. (1999). Млекопитающие мира Уокера (6-е изд.). Балтимор, США: Издательство Университета Джонса Хопкинса. стр. 1181–3. ISBN 978-0-8018-5789-8 .
- ^ Перейти обратно: а б с Флагстад, Огайо; Сиверстен, ПО; Стенсет, Северная Каролина; Якобсен, КС (2001). «Изменение окружающей среды и темпы эволюции: филогеографическая структура комплекса хартебистов в связи с климатическими изменениями» . Труды Королевского общества B: Биологические науки . 268 (1468): 667–77. дои : 10.1098/rspb.2000.1416 . ПМЦ 1088655 . ПМИД 11321054 .
- ^ Врба, ЕС (1979). «Филогенетический анализ и классификация ископаемых и современных Alcelaphini Mammalia: Bovidae». Биологический журнал Линнеевского общества . 11 (3): 207–28. дои : 10.1111/j.1095-8312.1979.tb00035.x .
- ^ Джентри, AW (2012). «Эволюция и распространение африканских Bovidae». В Бубенике, Джорджия; Бубеник, А.Б. (ред.). Рога, вилороги и рога: эволюция, морфология, физиология и социальное значение . Нью-Йорк, США: Спрингер. п. 216. ИСБН 978-1-4613-8966-8 .
- ^ Гроувс, К.; Грабб, П. (2011). Систематика копытных . Балтимор, США: Издательство Университета Джонса Хопкинса. п. 208. ИСБН 978-1-4214-0093-8 .
- ^ Мэти, Калифорния; Робинсон, Ти Джей (1999). «Филогения цитохрома b семейства Bovidae: разрешение внутри Alcelaphini, Antilopini, Neotragini и Tragelaphini». Молекулярная филогенетика и эволюция . 12 (1): 31–46. дои : 10.1006/mpev.1998.0573 . ПМИД 10222159 .
- ^ Арктандер, П.; Йохансен, К.; Кутельлек-Врето, Массачусетс (1999). «Филогеография трех близкородственных африканских быков (племя Alcelaphini)» . Молекулярная биология и эволюция . 16 (12): 1724–39. doi : 10.1093/oxfordjournals.molbev.a026085 . ПМИД 10605114 .
- ^ Перейти обратно: а б Флагстад, Огайо; Сиверцен, ПО; Стенсет, Северная Каролина; Стейси, Дж. Э.; Олсакер, И.; Рёд, К.Х.; Якобсен, КС (2000). «Генетическая изменчивость хартебеста Суэйна, находящейся под угрозой исчезновения антилопы Эфиопии». Биология сохранения . 14 (1): 254–64. дои : 10.1046/j.1523-1739.2000.98339.x . S2CID 84794781 .
- ^ « Алцелафус буселафус » . Интегрированная таксономическая информационная система . Проверено 7 апреля 2016 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н Восток, Р.; Группа специалистов МСОП/SSC по антилопам (1999). База данных африканских антилоп, 1998 год . Гланд, Швейцария: Комиссия МСОП по выживанию видов. стр. 186–93. ISBN 978-2-8317-0477-7 .
- ^ Перейти обратно: а б Группа специалистов МСОП по антилопам SSC (2017). « Alcelaphus buselaphus ssp. buselaphus » . Красный список исчезающих видов МСОП . 2017 : e.T813A50181474. doi : 10.2305/IUCN.UK.2017-2.RLTS.T813A50181474.en . Проверено 13 ноября 2021 г.
- ^ Мэллон, ДП; Кингсвуд, Южная Каролина (2001). Антилопы: Северная Африка, Ближний Восток и Азия . Гланд, Швейцария: МСОП. п. 25. ISBN 978-2-8317-0594-1 .
- ^ Перейти обратно: а б Группа специалистов МСОП по антилопам SSC (2017). « Alcelaphus buselaphus ssp. caama » . Красный список исчезающих видов МСОП . 2017 : e.T814A50181496. doi : 10.2305/IUCN.UK.2017-2.RLTS.T814A50181496.en . Проверено 13 ноября 2021 г.
- ^ Перейти обратно: а б Группа специалистов МСОП по антилопам SSC (2017). « Alcelaphus buselaphus ssp. lelwel » . Красный список исчезающих видов МСОП . 2017 : e.T816A50181544. doi : 10.2305/IUCN.UK.2017-2.RLTS.T816A50181544.en . Проверено 13 ноября 2021 г.
- ^ Рафферти, JP (2010). Грейзерс (1-е изд.). Нью-Йорк, США: Образовательные публикации Britannica. п. 121. ИСБН 978-1-61530-465-3 .
- ^ Перейти обратно: а б Группа специалистов МСОП по антилопам SSC (2017). « Alcelaphus buselaphus ssp. lichtensteinii » . Красный список исчезающих видов МСОП . 2017 : e.T812A50181339. doi : 10.2305/IUCN.UK.2017-2.RLTS.T812A50181339.en . Проверено 13 ноября 2021 г.
- ^ Перейти обратно: а б Льюис, Дж. Г.; Уилсон, RT (1977). «Тяжелое положение зверя Суэйна» . Орикс . 13 (5): 491–4. дои : 10.1017/S0030605300014551 . S2CID 84704549 .
- ^ Тамрат, М., Атикем, А., Флагстад, Ø, Фишер, М., Роос, К., Евангелиста, П., . . . Зиннер, Д. (2022). Оценка численности населения, генетическая изменчивость и конкуренция с домашним скотом. Орикс, 56(3), 336–344. doi:10.1017/S0030605320000927
- ^ Перейти обратно: а б Хилдьярд, А. (2001). Находящаяся под угрозой исчезновения дикая природа и растения мира . Нью-Йорк, США: Маршалл Кавендиш. стр. 674–5. ISBN 978-0-7614-7199-8 .
- ^ Перейти обратно: а б с д «Лельвел Хартебист» . Рекорды охоты на крупную дичь – Международная онлайн-книга рекордов Safari Club . Сафари Клуб Международный . Архивировано из оригинала 30 января 2016 года . Проверено 26 января 2016 г. .
- ^ Робинсон, Ти Джей; Моррис, диджей; Фэйролл, Н. (1991). «Межвидовая гибридизация Bovidae: стерильность потомства Alcelaphus buselaphus × Damaliscus dorcas F1». Биологическая консервация . 58 (3): 345–56. дои : 10.1016/0006-3207(91)90100-Н .
- ^ Перейти обратно: а б с д Кастельо, младший (2016). Крупные рогатые животные мира: антилопы, газели, крупный рогатый скот, козы, овцы и их родственники . Принстон, США: Издательство Принстонского университета. стр. 537–9. ISBN 978-0-691-16717-6 .
- ^ Августин, диджей; Веблен, Кентукки; Гохин, младший; Ригинос, К.; Янг, Т.П. (2011). «Пути позитивного взаимодействия крупного рогатого скота и дикой природы на полузасушливых пастбищах» (PDF) . В Георгиадисе, штат Нью-Джерси (ред.). Сохранение дикой природы в африканских ландшафтах: экосистема Эвасо в Кении . Смитсоновский вклад в зоологию. Том. 632. Вашингтон, округ Колумбия, США: Научная пресса Смитсоновского института. стр. 55–71. Архивировано (PDF) из оригинала 11 сентября 2015 г. Проверено 7 апреля 2016 г.
- ^ «Кенийский Хайленд Хартебист» . Рекорды охоты на крупную дичь – Международная онлайн-книга рекордов Safari Club . Сафари Клуб Международный . Архивировано из оригинала 31 января 2016 года . Проверено 26 января 2016 г. .
- ^ Перейти обратно: а б Ракстон, AE; Шварц, Э. (1929). «О гибридных антилопах и о распространении группы Alcelaphus buselaphus ». Труды Лондонского зоологического общества . 99 (3): 567–83. дои : 10.1111/j.1469-7998.1929.tb07706.x .
- ^ «Нойманн Хартебист» . Рекорды охоты на крупную дичь – Международная онлайн-книга рекордов Safari Club . Сафари Клуб Международный . Архивировано из оригинала 31 января 2016 года . Проверено 26 января 2016 г. .
- ^ Харрис, Дж.; Лики, М. (2001). Лотагам: Рассвет человечества в Восточной Африке . Нью-Йорк, США: Издательство Колумбийского университета. п. 547. ИСБН 978-0-231-11870-5 .
- ^ Бергер, ЛР; Хилтон-Барбер, Б. (2004). Полевой путеводитель по колыбели человечества: Стеркфонтейн, Сварткранс, Кромдраай и окрестности, объекты всемирного наследия (2-е (пересмотренное) изд.). Кейптаун, Южная Африка: Struik Publishers. п. 163. ИСБН 978-1-77007-065-3 .
- ^ Царь, Э.; Ижаки И.; Лев-Ядун, С.; Бар-Оз, Г.; Хансен, DM (2009). «Распространение и вымирание копытных в голоцене южного Леванта» . ПЛОС ОДИН . 4 (4): 5316–28. Бибкод : 2009PLoSO...4.5316T . дои : 10.1371/journal.pone.0005316 . ПМК 2670510 . ПМИД 19401760 .

- ^ Перейти обратно: а б с д и ж г час я дж Кингдон, Дж. (1989). Млекопитающие Восточной Африки: Атлас эволюции Африки . Том. 3, Часть D: Крупные рогатые скоты. Чикаго: Издательство Чикагского университета. ISBN 978-0-226-43725-5 .
- ^ Перейти обратно: а б Макдональд, Д. (1987). Энциклопедия млекопитающих . Нью-Йорк, США: факты в архиве. стр. 564–71 . ISBN 978-0-87196-871-5 .
- ^ Капеллини, И.; Гослинг, LM (2007). «Первичная продукция среды обитания и эволюция размеров тела внутри клады хартебесов» . Биологический журнал Линнеевского общества . 92 (3): 431–40. дои : 10.1111/j.1095-8312.2007.00883.x .
- ^ Перейти обратно: а б с «Вестерн Хартебист» . Рекорды охоты на крупную дичь – Международная онлайн-книга рекордов Safari Club . Сафари Клуб Международный . Архивировано из оригинала 30 января 2016 года . Проверено 26 января 2016 г. .
- ^ Перейти обратно: а б с «Мыс или Красный Хартебист» . Рекорды охоты на крупную дичь – Международная онлайн-книга рекордов Safari Club . Сафари Клуб Международный . Архивировано из оригинала 31 января 2016 года . Проверено 26 января 2016 г. .
- ^ Перейти обратно: а б «Кокс Хартбист» . Рекорды охоты на крупную дичь – Международная онлайн-книга рекордов Safari Club . Сафари Клуб Международный . Архивировано из оригинала 31 января 2016 года . Проверено 26 января 2016 г. .
- ^ Перейти обратно: а б с «Лихтенштейн Хартебист» . Рекорды охоты на крупную дичь – Международная онлайн-книга рекордов Safari Club . Сафари Клуб Международный . Архивировано из оригинала 31 января 2016 года . Проверено 26 января 2016 г. .
- ^ Перейти обратно: а б с «Суэйн Хартбист» . Рекорды охоты на крупную дичь – Международная онлайн-книга рекордов Safari Club . Сафари Клуб Международный . Архивировано из оригинала 30 января 2016 года . Проверено 26 января 2016 г. .
- ^ Перейти обратно: а б с д и ж г час я дж к л м Эстес, Р.Д. (2004). Руководство по поведению африканских млекопитающих: включая копытных, хищников и приматов (4-е изд.). Беркли, США: Издательство Калифорнийского университета. стр. 133–42 . ISBN 978-0-520-08085-0 .
- ^ Файерстоун, М. (2009). Наблюдение за дикой природой: Южная Африка; Южная Африка, Намибия, Ботсвана, Зимбабве, Малави, Замбия (2-е изд.). Футскрей, Австралия: Одинокая планета. стр. 228–9. ISBN 978-1-74104-210-8 .
- ^ Перейти обратно: а б с Хекель, Дж. О. (2007). Современный статус подвида антилопы Тора (PDF) (Отчет). Управление охраны дикой природы Эфиопии. стр. 100-1 1–13. Архивировано из оригинала (PDF) 3 февраля . Получено 26 января.
- ^ Перейти обратно: а б с д Капеллини, И. (2007). «Диморфизм у хартбеста». В Фэйрберне, диджей; Бланкенхорн, Ву; Секели, Т. (ред.). Пол, размер и гендерные роли: эволюционные исследования полового диморфизма размеров . Лондон, Великобритания: Издательство Оксфордского университета. стр. 124–32. doi : 10.1093/acprof:oso/9780199208784.003.0014 . ISBN 978-0-19-954558-2 .
- ^ Перейти обратно: а б Капеллини, И.; Гослинг, LM (2006). «Эволюция боевых структур у хартебеста» (PDF) . Исследования в области эволюционной экологии . 8 : 997–1011. Архивировано (PDF) из оригинала 14 января 2019 г. Проверено 7 апреля 2016 г.
- ^ Обусье, Х. (1970). «Информация об Alcelaphini (Bovidae-Mammalia) с особым упором на мозг и гипофиз. Результаты исследовательских поездок по Африке (1959–1967)». Gegenbaurs Morphologisches Jahrbuch . 114 (3): 393–435. ПМИД 5523305 .
- ^ Шаллер, Великобритания (1976). Лев Серенгети: исследование отношений хищник-жертва . Чикаго, США: Издательство Чикагского университета. стр. 461–5 . ISBN 978-0-226-73640-2 .
- ^ Элтрингем, СК (1979). Экология и охрана крупных африканских млекопитающих (1-е изд.). Лондон, Великобритания: Макмиллан. п. 177. ИСБН 978-0-333-23580-5 .
- ^ Верлинден, А. (1998). «Сезонные модели передвижения некоторых копытных в экосистеме Калахари в Ботсване в период с 1990 по 1995 год» . Африканский журнал экологии . 36 (2): 117–28. дои : 10.1046/j.1365-2028.1998.00112.x . S2CID 85774858 .
- ^ Бумкер, Дж.; Горак, ИГ; Де Вос, В. (1986). «Гельминты-паразиты различных парнокопытных из некоторых заповедников Южной Африки». Журнал ветеринарных исследований Ондерстепоорта . 53 (2): 93–102. ПМИД 3725333 .
- ^ Перейти обратно: а б Ховард, GW (1977). «Распространенность носовых ботов (Diptera: Oestridiae) у некоторых замбийских зверей» . Журнал болезней дикой природы . 13 (4): 400–4. дои : 10.7589/0090-3558-13.4.400 . ПМИД 24228960 . S2CID 27306683 .
- ^ Пестер, ФРН; Лоуренс, БР (2009). «Паразитная нагрузка некоторых африканских охотничьих животных». Журнал зоологии . 174 (3): 397–406. дои : 10.1111/j.1469-7998.1974.tb03167.x .
- ^ Спитальска, Э.; Ридделл, М.; Хейн, Х.; Спарагано, ОА (2005). «Распространенность тейлериоза у красного зверя ( Alcelaphus buselaphus caama ) в Намибии» Паразитологические исследования . 97 (1): 77–9. дои : 10.1007/s00436-005-1390-y . ISSN 1432-1955 . ПМИД 15986252 . S2CID 23721115 .
- ^ Спинаж, Калифорния (2012). Африканская экология: ориентиры и исторические перспективы . Берлин, Германия: Шпрингер. п. 1176. ИСБН 978-3-642-22872-8 .
- ^ Белем, АМГ; Баконе, Э. У. (2009). «Желудочно-кишечные паразиты антилоп и буйволов ( brachyceros ) с охотничьего ранчо Назинга в Буркина-Фасо» Syncerus caffer . Биотехнология, агрономия, общество и окружающая среда (на французском языке). 13 (4): 493–8. ISSN 1370-6233 . Архивировано из оригинала 18 августа 2016 г. Проверено 7 апреля 2016 г.
- ^ Хоберг, Е.П.; Абрамс, А.; Пиллитт, Пенсильвания (2009). « Robustostrongylus aferensis gen. nov. et sp. nov. (Nematoda: Trichostrongyloidea) в початках ( Kobus cob ) и хартебесте ( Alcelaphus buselaphus jacksoni ) (Парнокопытные) из Африки к югу от Сахары, с дальнейшими размышлениями об Ostertagiinae» . Журнал паразитологии 95 (3): 702–17. дои : 10.1645/ge-1859.1 . ПМИД 19228080 . S2CID 7641994 . Архивировано из оригинала 0 марта 2022 г. Получено 2 апреля 2018 г.
- ^ Ле Ру, Польша (1931). «На Longistrongylus meyeri gen. и sp. nov., трихостронгиле, паразитирующем на Red Hartebeest Bubalis caama ». Журнал гельминтологии . 9 (3): 141. дои : 10.1017/S0022149X00030376 . S2CID 86009616 .
- ^ Перейти обратно: а б с д и ж «Хартебист» . Африканский фонд дикой природы . Архивировано из оригинала 26 января 2013 года . Проверено 20 января 2013 г.
- ^ Перейти обратно: а б с Шютте, младший; Лесли, DM; Лохмиллер, РЛ; Дженкс, Дж. А. (1998). «Диета хартебеста и чалой антилопы в Буркина-Фасо: поддержка давней гипотезы» . Журнал маммологии . 79 (2): 426–36. дои : 10.2307/1382973 . JSTOR 1382973 . S2CID 83671165 .
- ^ Мюррей, МГ (1993). «Сравнительное питание гну, антилопы и топи в Серенгети» Африканский журнал экологии . 31 (2): 172–7. дои : 10.1111/j.1365-2028.1993.tb00530.x .
- ^ Кейсбир, РЛ ; Косс, Г.Г. (1970). «Пищевые привычки антилоп гну, зебр, антилоп и крупного рогатого скота в кенийском Масаиленде» Африканский журнал экологии . 8 (1): 25–3 дои : 10.1111/j.1365-2028.1970.tb00827.x .
- ^ Перейти обратно: а б Миллс, Г.; Хес, Л. (1997). Полная книга млекопитающих Южной Африки . Кейптаун, Южная Африка: Struik Publishers. п. 255. ИСБН 978-0-947430-55-9 .
- ^ Ядав, PR (2004). Исчезающие и исчезающие виды . Нью-Дели, Индия: Издательство Discovery. стр. 139–40. ISBN 978-81-7141-776-6 .
- ^ Мэллон, ДП; Кингсвуд, Южная Каролина (2001). Антилопы: Северная Африка, Ближний Восток и Азия . Гланд, Швейцария: МСОП. ISBN 978-2-8317-0594-1 .
- ^ Харпер, Ф. (1945). Вымершие и исчезающие млекопитающие Старого Света . Нью-Йорк, США: Американский комитет международной защиты дикой природы. стр. 642–8.
- ^ Группа специалистов МСОП по антилопам SSC (2017). « Alcelaphus buselaphus ssp. cokii » . Красный список исчезающих видов МСОП . 2017 : e.T815A50181521. doi : 10.2305/IUCN.UK.2017-2.RLTS.T815A50181521.en . Проверено 13 ноября 2021 г.
- ^ Группа специалистов МСОП по антилопам SSC (2017). « Alcelaphus buselaphus ssp. tora » . Красный список исчезающих видов МСОП . 2017 : e.T810A50180985. doi : 10.2305/IUCN.UK.2017-2.RLTS.T810A50180985.en . Проверено 13 ноября 2021 г.
- ^ Группа специалистов МСОП по антилопам SSC (2017). « Alcelaphus buselaphus ssp. swaynei » . Красный список исчезающих видов МСОП . 2017 : e.T809A3145291. doi : 10.2305/IUCN.UK.2017-2.RLTS.T809A3145291.en . Проверено 13 ноября 2021 г.
- ^ Датико, Д.; Бекеле, А. (2011). «Популяционный статус и воздействие на человека находящегося под угрозой исчезновения зверя суэйна ( Alcelaphus buselaphus swaynei ) на равнинах Нечисар, национальный парк Нечисар, Эфиопия» Африканский журнал экологии . 49 (3): 311–9. дои : 10.1111/j.1365-2028.2011.01266.x .
- ^ Группа специалистов МСОП по антилопам SSC (2017). « Alcelaphus buselaphus ssp. major » . Красный список исчезающих видов МСОП . 2017 : e.T817A50181578. doi : 10.2305/IUCN.UK.2017-2.RLTS.T817A50181578.en . Проверено 13 ноября 2021 г.
- ^ Перейти обратно: а б Ван Нир, В.; Линзеле, В.; Фридман, РФ (2004). «Захоронения животных и подношение еды на элитном кладбище HK6 Иераконполя» . В Хендриксе, С.; Фридман, Р; Циалович, К.; Хлодницкий М. (ред.). Египет у истоков: исследования памяти Барбары Адамс . Ориенталия Лованиенсия Аналекта. Том. 138. Левен, Бельгия: Издательство Peeters. п. 111. ИСБН 978-90-429-1469-8 . Архивировано из оригинала 8 марта 2022 г. Проверено 1 ноября 2020 г.
- ^ Хоффман, Л.К.; Смит, К.; Мюллер, Н. (2010). «Химические характеристики мяса красного антилопы ( Alcelaphus buselaphus caama )» . Южноафриканский журнал зоотехники . 40 (3): 221–8. дои : 10.4314/sajas.v40i3.6 .
Внешние ссылки
[ редактировать ] СМИ, связанные с Alcelaphus buselaphus , на Викискладе?
СМИ, связанные с Alcelaphus buselaphus , на Викискладе?  Данные, относящиеся к Alcelaphus buselaphus , в Wikispecies
Данные, относящиеся к Alcelaphus buselaphus , в Wikispecies
| Базы данных органов управления : Национальные |
|---|