Внешняя морфология чешуекрылых






Внешняя морфология чешуекрылых — физиологическое строение тела насекомых отряда чешуекрылых , известных также как бабочки и мотыльки . Чешуекрылые отличаются от других отрядов наличием чешуи на внешних частях тела и придатках, особенно крыльях. Бабочки и мотыльки различаются по размеру: от микрочешуекрылых длиной всего несколько миллиметров до размаха крыльев во много дюймов, как у мотылька Атласа . Чешуекрылые, насчитывающие более 160 000 описанных видов, обладают вариациями основной структуры тела, которые эволюционировали, чтобы получить преимущества в адаптации и распространении. [1]
Чешуекрылые подвергаются полной метаморфозе , проходя четыре стадии жизненного цикла : яйцо , личинка или гусеница , куколка или куколка и имаго (множественное число: воображает ) /взрослая особь. Личинки — гусеницы — имеют утолщённую ( склеротизированную ) головную капсулу, жевательный ротовой аппарат , мягкое тело, которое может иметь волосовидные или другие выступы, три пары настоящих ног и до пяти пар ложноножек . Большинство гусениц — травоядные , но некоторые — плотоядные (некоторые едят муравьев, тлю или других гусениц) или детритофаги . [2] Личинки находятся на стадиях питания и роста и периодически подвергаются гормонально-индуцированному шелушению , развиваясь дальше с каждым возрастом , пока не претерпят финальную личиночно-куколочную линьку. Личинки многих видов чешуекрылых либо образуют шелковую оболочку, называемую коконом , и окукливаются внутри нее, либо окукливаются в ячейке под землей. У многих бабочек куколка подвешена к кремастеру и называется куколкой .
Тело взрослого человека имеет твердый экзоскелет , за исключением брюшка, который менее склеротизирован. Голова по форме напоминает капсулу с отходящими от нее придатками. Ротовой аппарат взрослых особей включает выступающий хоботок , образованный верхнечелюстными галеями и приспособленный для всасывания нектара. Некоторые виды не питаются во взрослом возрасте и могут иметь уменьшенный ротовой аппарат, в то время как у других они модифицированы для прокалывания и всасывания крови или фруктовых соков. [3] Мандибулы отсутствуют у всех, кроме Micropterigidae , у которых есть жевательный ротовой аппарат . [4] Взрослые чешуекрылые имеют два неподвижных многогранных сложных глаза и только два простых глаза или глазка, которые могут быть редуцированы. [5] Три сегмента грудной клетки слиты вместе. Усики выдающиеся и, помимо обоняния, также помогают ориентироваться, ориентироваться и сохранять равновесие во время полета. [6] У бабочек самцы часто имеют более перистые усики, чем самки, что позволяет им обнаруживать феромоны самок на расстоянии. Имеются две пары перепончатых крыльев , отходящих от среднегрудного (среднего) и заднегрудного (третьего) сегментов; они обычно полностью покрыты минутной чешуей . Два крыла с каждой стороны действуют как одно благодаря механизмам блокировки крыльев . В некоторых группах самки нелетают и имеют редуцированные крылья. Брюшко имеет десять сегментов, соединенных подвижными межсегментарными перепонками. Последние сегменты брюшка образуют наружные половые органы . Гениталии сложны и служат основой для идентификации семьи и дискриминации видов. [7]
Крылья, головные части грудной клетки и брюшко чешуекрылых покрыты мелкими чешуйками, от которых отряд чешуекрылых получил свое название, слово lepidos на древнегреческом языке означает «чешуя». Большинство чешуек пластинчатые (лезвийные) и прикреплены к ножке, в то время как другие формы могут напоминать волосы или специализироваться как вторичные половые признаки. Просвет, или поверхность ламели, имеет сложное строение. Он придает цвет либо за счет содержащихся в нем пигментов , либо за счет своей трехмерной структуры. [8] Чешуя выполняет ряд функций, в том числе изоляцию , терморегуляцию и помощь в полете, наиболее важной из которых является большое разнообразие ярких или нечетких рисунков, которые они обеспечивают, которые помогают организму защитить себя путем маскировки , мимикрии и поиска. приятели.
Внешняя морфология
[ редактировать ]
Как и другие представители надотряда Holometabola , Lepidoptera подвергаются полной метаморфозе , проходя четырехстадийный жизненный цикл: яйцо, личинка/гусеница, куколка/куколка и имаго (множественное число: воображает )/взрослая особь. [9]
Размер чешуекрылых варьируется от нескольких миллиметров в длину, как, например, у микрочешуекрылых, до размаха крыльев во много дюймов, как у бабочки Атласа и птичьего крыла крупнейшей в мире бабочки королевы Александры . [10] : 246
Общий план кузова
[ редактировать ]Тело взрослой бабочки или мотылька (имаго) имеет три отдельных отдела, называемых тагматами , соединенных перетяжками; эти тагматы — это голова, грудная клетка и брюшко. Взрослые чешуекрылые имеют четыре крыла — переднее и заднее на левой и правой стороне грудной клетки — и, как и все насекомые, три пары ног. [11]
Морфологические характеристики , отличающие отряд Lepidoptera от других отрядов насекомых: [10] : 246
- Голова : На голове большие сложные глаза и, если имеются ротовые части, они почти всегда представляют собой хоботок, похожий на соломинку для питья.
- Чешуя : Чешуя покрывает внешнюю поверхность тела и придатки.
- Грудная клетка : Переднегруди обычно уменьшена.
- Крылья : Две пары крыльев присутствуют почти у всех таксонов. На крыльях очень мало поперечных жилок .
- Брюшко : Задние брюшные сегменты сильно модифицированы для размножения. Церки отсутствуют.
- Личинка : Личинки чешуекрылых известны как гусеницы и имеют хорошо развитую голову и челюсти. У них может быть от нуля до пяти пар ложноножек , обычно четыре.
- Куколка : Куколки у большинства видов адектические (без функциональных челюстей в состоянии куколки) и обтектные (с придатками, сросшимися или приклеенными к телу), в то время как у других видов есть дектические (с функциональными мандибулами, присутствующими в состоянии куколки) и экзоратные (имеющие усики, ноги и крылья свободны).
Отличительные таксономические признаки
[ редактировать ]Основными характеристиками, используемыми для классификации видов, родов и семейств чешуекрылых, являются: [12]
- ротовой аппарат
- форма и жилкование крыльев
- являются ли крылья однородными (жилкование передних и задних крыльев одинаковое) или разноневральными (передние и задние крылья разные)
- являются ли крылья шиповатыми (более или менее покрытыми специальными щетинками, называемыми микрощетинками) или нет
- тип смыкания крыла (югата или френат)
- анатомия репродуктивных органов
- строение личинки и положение первичных щетинок
- является ли куколка экзаратом или объектом
Морфологические характеристики гусениц и куколок, использованные для классификации, совершенно отличаются от таковых у взрослых особей; [13] : 637 [14] различные схемы классификации иногда предоставляются отдельно для классификации взрослых особей, личинок и куколок. [14] [15] : 28–40 Характеристики неполовозрелых стадий все чаще используются в таксономических целях, поскольку они дают представление о систематике и филогении чешуекрылых, которые не очевидны при обследовании взрослых особей. [15] : 28
Голова
[ редактировать ]
Как и все головы животных, голова бабочки или мотылька содержит органы питания и основные органы чувств. Голова обычно состоит из двух усиков, двух сложных глаз, двух щупиков и хоботка. [11] У чешуекрылых есть глазки , которые могут быть видимыми, а могут и не быть видимыми. У них также есть сенсорные структуры, называемые хетосемами , функции которых в значительной степени неизвестны. Голова в основном заполнена мозгом, сосущим насосом и связанными с ним мышечными пучками. [16] В отличие от взрослых личинок имеют одночлениковые жвалы. [16]
Головная капсула хорошо склеротизирована и имеет ряд склеритов или пластинок, разделенных швами . Склериты трудно отличить от борозд (единственное число – sulcus ), которые представляют собой вторичные утолщения. Области головы разделены на ряд областей, которые служат топографическим ориентиром для описания лепидоптерологами, но не могут быть различены с точки зрения их развития. [16] Голова покрыта волосообразными или пластинчатыми чешуйками и встречается либо в виде пучков на лбу или макушке (так называемая грубая чешуя ), либо прижата близко к голове (так называемая гладкая чешуя ).
Органы чувств и структуры головы отличаются большим разнообразием, а форма и форма этих структур, а также их наличие или отсутствие являются важными таксономическими показателями для отнесения таксонов к семействам. [13]
 Голова бабочки семейства Gracillariidae с чешуей на голове.
Голова бабочки семейства Gracillariidae с чешуей на голове. Крупночешуйчатая голова бабочки Monopis icterogastra (семейство Tineidae )
Крупночешуйчатая голова бабочки Monopis icterogastra (семейство Tineidae ) Гладчешуйчатая голова бабочки Glyphipterix simpliciella (семейство Glyphipterigidae )
Гладчешуйчатая голова бабочки Glyphipterix simpliciella (семейство Glyphipterigidae ) Гладчешуйчатая головка бабочки Stegasta variana (семейство Gelechiidae )
Гладчешуйчатая головка бабочки Stegasta variana (семейство Gelechiidae )




Антенны
[ редактировать ]

Усики представляют собой выступающие парные придатки, выступающие вперед между глазами животного и состоящие из нескольких сегментов. У бабочек их длина варьируется от половины длины переднего крыла до трех четвертей длины переднего крыла. Усики бабочек либо тонкие, с бугорками на конце, а у Hesperiidae — крючковидные на конце. У некоторых родов бабочек, таких как Libythea и Taractrothera, шишка снизу имеет полость. [11] Усики бабочек бывают нитевидными (нитевидными), одногребешковыми (гребешковыми), двугребешковыми (перьевыми), крючковатыми, булавовидными или утолщенными. [13] : 636 Bombyx mandarina — пример двугребешковых усиков. [17] У некоторых бабочек, например семейства Castniidae, есть шишковидные усики, похожие на таковые у бабочек . [18]
Усики — основные органы обоняния (обоняния) у чешуекрылых. Поверхность антенны покрыта большим количеством обонятельных чешуек, волосков или ямок; их насчитывается до 1 370 000 на усиках монарха . Антенны чрезвычайно чувствительны; пернатые усики самцов бабочек Saturniidae, Lasiocampidae и многих других семейств настолько чувствительны, что могут обнаруживать феромоны самок бабочек на расстоянии до 2 км (1,2 мили). Усики чешуекрылых могут располагаться под разными углами. Они помогают насекомому обнаружить запах и могут рассматриваться как своего рода «обонятельный радар». [6] У бабочек самцы часто имеют более перистые, чем у самок, усики, позволяющие обнаруживать феромоны самок на расстоянии. [7] Поскольку самкам не нужно обнаруживать самцов, у них более простые усики. [6] Также было обнаружено, что антенны играют роль в ориентации по солнечному компасу с компенсацией по времени у мигрирующих бабочек-монархов . [19]
 Нитевидные усики — Eriocrania cicatricella ( Eriocraniidae )
Нитевидные усики — Eriocrania cicatricella ( Eriocraniidae ) Одногребенчатые усики — Abantiades barcas ( Hepialidae )
Одногребенчатые усики — Abantiades barcas ( Hepialidae ) Двугребенчатые усики — Actias artemis ( Saturniidae )
Двугребенчатые усики — Actias artemis ( Saturniidae ) Крючковатые усики — Epargyreus clarus ( Hesperiidae )
Крючковатые усики — Epargyreus clarus ( Hesperiidae ) Булавовидные усики - Vanessa atalanta ( Nymphalidae )
Булавовидные усики - Vanessa atalanta ( Nymphalidae ) Утолщенные усики – Deleiphila elpenor ( Sphingidae )
Утолщенные усики – Deleiphila elpenor ( Sphingidae ) Усики булавовидной бабочки - Athis inca ( Castniidae )
Усики булавовидной бабочки - Athis inca ( Castniidae )








Глаза
[ редактировать ]

У чешуекрылых есть два больших неподвижных сложных глаза, которые состоят из большого количества фасеток или линз, каждая из которых соединена с линзообразным цилиндром, прикрепленным к нерву, ведущему к мозгу . [11] Каждый глаз может иметь до 17 000 отдельных световых рецепторов ( омматидиев ), которые в совокупности обеспечивают широкое мозаичное представление окружающей области. [6] У одного тропического азиатского семейства, Amphitheridae , сложные глаза разделены на два отдельных сегмента. [13] [20] Глаза обычно гладкие, но могут быть покрыты мелкими волосками. Глаза бабочек обычно коричневые, золотисто-коричневые или даже красные, как у некоторых видов шкиперов . [11]
В то время как у большинства насекомых есть три простых глаза, или глазка , у всех видов чешуекрылых, за исключением нескольких мотыльков, присутствуют только два глазка, по одному на каждой стороне головы возле края сложного глаза. У некоторых видов органы чувств, называемые хетосематами . рядом с глазками расположены [11] [21] Глазки не гомологичны простым глазам гусениц, которые по-другому называются стеммами . [5] Глазки чешуекрылых у некоторых семейств редуцированы снаружи; там, где они присутствуют, они не сфокусированы, в отличие от стеблей личинок, которые полностью сфокусированы. Полезность глазков в настоящее время не понятна. [5]
Бабочки и мотыльки могут видеть ультрафиолетовый (УФ) свет, а цвет и узор крыльев чешуекрылые наблюдают в основном в этих длинах волн света. [13] Узоры на их крыльях, видимые в ультрафиолетовом свете, значительно отличаются от тех, которые видны при обычном свете. Ультрафиолетовые узоры действуют как визуальные подсказки, которые помогают различать виды для спаривания. Были проведены исследования рисунков крыльев чешуекрылых (в основном бабочек), освещенных УФ-светом. [13]
Пальпация
[ редактировать ]
Обычно губные щупики выпуклые, трехчлениковые, выступают из-под головы и загибаются вверх перед лицом. [7] Морфология губных щупиков у разных семейств чешуекрылых сильно различается; иногда щупики отдельные, иногда сросшиеся и образуют клюв, но они всегда самостоятельно подвижны. В других случаях губные щупики могут быть не вертикальными, а прямыми (выступать вперед горизонтально). [11] [13] Щупики состоят из короткого базального сегмента, сравнительно длинного центрального сегмента и узкой терминальной части. Первые два сегмента покрыты густыми чешуйками и могут быть волосатыми ; терминальный сегмент голый. Конечный сегмент может быть тупым или заостренным; он может выступать прямо или под углом из второго сегмента, внутри которого он может быть скрыт. [11]
Ротовые органы
[ редактировать ]

В то время как жвалы или челюсти (жевательный ротовой аппарат) присутствуют только на стадии гусеницы, ротовой аппарат большинства взрослых чешуекрылых в основном состоит из сосущего типа; эта часть известна как хоботок или хаустеллум . У некоторых видов чешуекрылых ротовой аппарат редуцирован, и во взрослом состоянии они не питаются. Другие, такие как базальное семейство Micropterigidae, имеют жевательный ротовой аппарат. [21]
Хоботок (множественное число — хоботки ) образован из верхнечелюстных косточек и приспособлен для всасывания нектара . [3] Он состоит из двух трубок, скрепленных крючками и отделяемых для очистки. Каждая трубка вогнута внутрь, образуя центральную трубку, через которую всасывается влага. Всасывание осуществляется за счет сокращения и расширения мешочка в голове. [11] Хоботок свернут под головой, когда насекомое отдыхает, и вытягивается только во время кормления. [16] Нижнечелюстные щупики редуцированы и даже рудиментарны. Они заметны; пять сегментированы в некоторые из более базальных семейств и часто складчаты. [7]
Форма и размеры хоботка изменились, чтобы дать различным видам более широкий и, следовательно, более полезный рацион. [3] Существует аллометрическая зависимость между массой тела чешуекрылых и длиной хоботка. [22] интересным адаптивным отклонением от него является необычайно длинноязычная бабочка-сфинкс Xanthopan morganii praedicta . Чарльз Дарвин предсказал существование и длину хоботка этой бабочки до ее открытия, основываясь на своих знаниях о мадагаскарской звездчатой орхидее Angraecum sesquipedale с длинными шпорами . [23]
У чешуекрылых в основном есть две кормовые гильдии : нектароядные, которые получают большую часть своих пищевых потребностей из цветочного нектара, и плодоядные, которые питаются в основном соками гниющих фруктов или ферментирующим древесным соком. В морфологии хоботков обеих кормовых гильдий имеются существенные различия. Бражники (семейство Sphingidae ) имеют удлиненные хоботки, которые позволяют им питаться и опылять цветы с длинными трубчатыми венчиками. Помимо этого, у некоторых таксонов (особенно совок ) развилась иная морфология хоботков. У некоторых видов совок есть колючий ротовой аппарат; хоботок имеет на кончике склеротизованные чешуйки, с помощью которых можно прокалывать и высасывать кровь или фруктовые соки. Хоботки некоторых видов Heliconius развились, чтобы потреблять твердые вещества, такие как пыльца . [24] Некоторые другие бабочки, в основном совки, модифицировали хоботки в соответствии со своим способом питания – лакримофагией (питание слезами спящих птиц). Хоботки часто имеют острые вершины и множество зазубрин и шпор на ножке. [25] [26]
 Сканирующая электронная микрофотография хоботка бабочки семейства Pyralidae.
Сканирующая электронная микрофотография хоботка бабочки семейства Pyralidae. Бабочка -нимфалида, сосущая банан
Бабочка -нимфалида, сосущая банан Сара длиннокрылая ( Heliconius sara ), один из многих видов Heliconius, которые, как известно, питаются пыльцой, с пыльцой на хоботке.
Сара длиннокрылая ( Heliconius sara ), один из многих видов Heliconius, которые, как известно, питаются пыльцой, с пыльцой на хоботке. Xanthopan morganii , африканский сфингид , имеет хоботок длиной в фут, приспособленный для питания орхидеей Angraecum sesquipedale.
Xanthopan morganii , африканский сфингид , имеет хоботок длиной в фут, приспособленный для питания орхидеей Angraecum sesquipedale. Слезоядные чешуекрылые, такие как две бабочки Джулия ( Dryas iulia ), пьющие слезы черепах в Эквадоре, имеют крючки и зазубрины на кончике хоботка.
Слезоядные чешуекрылые, такие как две бабочки Джулия ( Dryas iulia ), пьющие слезы черепах в Эквадоре, имеют крючки и зазубрины на кончике хоботка.





грудная клетка
[ редактировать ]

Грудная клетка, развивающаяся из 2, 3 и 4 сегментов личинки, состоит из трех невидимо разделенных сегментов: переднегруди , заднегруди и среднегруди . [11] На грудной клетке расположены органы передвижения насекомых – ноги и крылья. Передние ноги выходят из переднегруди, передние крылья и средняя пара ног несут на среднегруди, а задние крылья и задние ноги возникают из заднегруди. В некоторых случаях крылья рудиментарны. [11] [27]
Верхняя и нижняя части грудной клетки ( терга и грудина соответственно) состоят из сегментарных и внутрисегментарных склеритов , которые у чешуекрылых демонстрируют вторичную склеротизацию и значительную модификацию. Переднегруди — самый простой и маленький из трех сегментов, а среднегруди — наиболее развитый. [27]
Между головой и грудной клеткой находится перепончатая шейка матки . Он состоит из пары латеральных шейных склеритов и состоит как из головных, так и из грудных элементов. [10] : 71 [27] Между головой и грудной клеткой находится чешуйка, называемая переднеспинкой . По обеим сторонам находится щитовидная чешуя, называемая лопаткой . [11] У Noctuoidea заднегруди видоизменена парой барабанных органов . [7]
Нога
[ редактировать ]Передние конечности у Papilionoidea демонстрируют редукцию различных форм: бабочки семейства Nymphalidae или бабочки с кистями, как их обычно называют, имеют только две задние пары полностью функциональных ног, а передняя пара сильно редуцирована и не способна ходить или садиться. У Lycaenidae лапка несегментированная, так как лапки срослись, когти лапок отсутствуют. Аролиарная подушечка (подушечка, выступающая между тарзальными когтями некоторых насекомых) и пульвиллы (единственное число: pulvillus, подушечка или лопасть под каждым тарзальным когтем) редуцированы или отсутствуют у Papilionidae . Предплюсневые когти также отсутствуют у Riodinidae . [28]
У чешуекрылых три пары ног покрыты чешуей. [13] У чешуекрылых также есть органы обоняния на ногах, которые помогают «ощущать вкус» или «нюхать» пищевые растения. [6]
Крылья
[ редактировать ]Взрослые чешуекрылые имеют две пары перепончатых крыльев, обычно полностью покрытых мелкой чешуей. Крыло состоит из верхней и нижней перепонок, которые соединены мельчайшими волокнами и укреплены системой утолщенных полых ребер, которые в народе, но неправильно называются «венами», поскольку они также могут содержать трахеи, нервные волокна и кровеносные сосуды. [11] [29] Перепонки покрыты мелкими чешуйками с зазубренными концами или волосками и прикреплены крючками. Крылья двигаются за счет быстрого мышечного сокращения и расширения грудной клетки. [11]
Крылья возникают из средне- и заднегрудных сегментов и сходны по размерам в базальных группах. В более производных группах среднегрудные крылья крупнее, с более мощной мускулатурой у основания и более жесткими венозными структурами на реберном крае. [7]
Помимо выполнения основной функции полета, крылья также выполняют второстепенные функции самозащиты , маскировки и терморегуляции. [3] [30] У некоторых семейств Lepidoptera, таких как Psychidae и Lymantriidae , крылья редуцированы или даже отсутствуют (часто у самок, но не у самцов). [7]
Форма
[ редактировать ]Форма крыльев у чешуекрылых весьма разнообразна. У Papilionoidea ребро может быть прямым или сильно изогнутым. он иногда вогнутый На заднем крыле . Иногда на переднем крыле бывают пильчатые или пилообразные зубцы. Вершина может быть округлой, заостренной или серповидной (выпуклой и вогнутой внизу). Термен имеет тенденцию быть прямым или вогнутым на переднем крыле , тогда как на заднем крыле он обычно более или менее выпуклый. Термен часто кренулированный или зубчатый , т. е. образуется у каждой жилки и вогнутый между ними. Спина обычно прямая, но может быть вогнутой. [11]
Заднее крыло часто хвостатое , т. е. жилки на конце торнуса имеют один или несколько хвостов. Сам торнус часто образуется и часто имеет лопасти. [11] Вдоль термена заднего крыла расположены плотно упакованные чешуи в двойной ряд. Нижняя сторона чешуи выступает и образует правильную узкую бахрому, называемую ресничками . [11]
 У шлейковых бабочек (семейство Pterophoridae ) крылья разделены.
У шлейковых бабочек (семейство Pterophoridae ) крылья разделены. У многоперых бабочек (семейство Alucitidae ) крылья разделены вдоль каждой жилки.
У многоперых бабочек (семейство Alucitidae ) крылья разделены вдоль каждой жилки. Microlepidoptera Gelechioidea , такие как Palumbina guerinii , имеют волосообразную бахрому вдоль задних крыльев.
Microlepidoptera Gelechioidea , такие как Palumbina guerinii , имеют волосообразную бахрому вдоль задних крыльев. Хвостатые задние крылья мадагаскарской закатной бабочки ( Chrysiridia rhipheus семейства Uraniidae )
Хвостатые задние крылья мадагаскарской закатной бабочки ( Chrysiridia rhipheus семейства Uraniidae ) Ликаниды, такие как обезьяна-головоломка ( Rathinda amor ), имеют нитевидные хвосты, которые пытаются объяснить гипотезой «ложной головы». [31]
Ликаниды, такие как обезьяна-головоломка ( Rathinda amor ), имеют нитевидные хвосты, которые пытаются объяснить гипотезой «ложной головы». [31]
 Pachyerannis obliquaria , брачная пара – вверху крылатый самец, внизу маленькая бескрылая самка.
Pachyerannis obliquaria , брачная пара – вверху крылатый самец, внизу маленькая бескрылая самка.







Жилкование
[ редактировать ]

Через двухслойное перепончатое крыло проходят трубчатые жилки. Вены соединены с гемоцелем и теоретически позволяют гемолимфе течь через них. Кроме того, через вены могут проходить нерв и трахея. [29]
Жилкование чешуекрылых простое, перекладин мало. [15] : 88 Жилкование крыльев у Lepidoptera является диагностическим признаком различия таксонов, а также родов и семейств. [13] Терминология основана на системе Комстока-Нидхэма , которая дает морфологическое описание жилкования крыльев насекомых . [32] У базальных чешуекрылых жилкование переднего крыла такое же, как и у заднего; состояние, называемое «гомонным». Жилкование Micropterigidae ( Zeugloptera ) напоминает наиболее примитивных ручейников (Trichoptera). Все остальные чешуекрылые, подавляющее большинство (около 98%), «гетероневральные», жилкование заднего крыла отличается от жилкования переднего крыла, а иногда и редуцировано. Мотыльки семейств Nepticulidae , Opostegidae , Gracillariidae , Tischeriidae и Bucculatricidae , среди прочих, часто имеют сильно уменьшенное жилкование на обоих крыльях. [13] : 635 [32] : 56 Однородные бабочки, как правило, имеют форму соединения крыльев «югум», в отличие от расположения «уздечка-удерживатель» в случае более продвинутых семейств.
 Жилкование крыльев насекомых с названиями по системе Комстока-Нидхэма.
Жилкование крыльев насекомых с названиями по системе Комстока-Нидхэма. Гомонное жилкование у Sabatinca lucilia ( Micropterigidae ).
Гомонное жилкование у Sabatinca lucilia ( Micropterigidae ). Гетероневральное жилкование у Gonepteryx rhamni ( Pieridae )
Гетероневральное жилкование у Gonepteryx rhamni ( Pieridae ) Пониженное жилкование у Synanthedon Tipuliformis ( Sesiidae ).
Пониженное жилкование у Synanthedon Tipuliformis ( Sesiidae ).




Муфта крыла
[ редактировать ]
У чешуекрылых в имаго развилось большое разнообразие морфологических механизмов соединения крыльев, которые делают эти таксоны «функционально двукрылыми» (двукрылыми). [33] Все формы, кроме самых базальных, демонстрируют такое соединение крыльев. [34] Существует три различных типа механизмов: скуловой, уздечно-ретинакулярный и плексиформный. [35]
У более примитивных групп есть увеличенная лопастная область возле базального заднего края (т. е. у основания переднего крыла), называемая югумом , которая во время полета складывается под заднее крыло. [7] [34] У других групп есть уздечка на заднем крыле, которая зацепляется под удерживатель на переднем крыле. [7]
У всех бабочек (за исключением самцов Euschemoninae ) и у бабочек Bombycoidea (за исключением Sphingidae ) отсутствует расположение уздечки и удерживателя для соединения крыльев. Вместо этого увеличенная плечевая область заднего крыла широко перекрывается передним крылом. Несмотря на отсутствие специфической механической связи, крылья перекрываются и работают синфазно. Мощный удар переднего крыла синхронно толкает заднее крыло. Этот тип соединения является разновидностью типа уздечки, но при этом уздечка и удерживатель полностью утрачены. [33] [36]
Весы
[ редактировать ]
Крылья чешуекрылых мелкочешуйчатые , что и дало название этому отряду; Название Lepidoptera было придумано в 1735 году Карлом Линнеем для группы «насекомых с четырьмя чешуйчатыми крыльями». Оно происходит от древнегреческого леписа (λεπίς), что означает «(рыбья) чешуя» (и связано с лепеином «очищать») и птерона (πτερόν), означающего «крыло». [37]
Чешуя также покрывает голову, части грудной клетки и живота, а также части гениталий. Морфологию чешуи изучали Дж. К. Дауни и А. С. Аллин (1975). [38] и чешуи были разделены на три группы, а именно волосообразные, или пилообразные , пластинчатые, пластинчатые и другие различные формы. [8]
Примитивные бабочки (non-Glossata и Eriocranidae) имеют «твердые» чешуи, неперфорированные, т. е. у них отсутствует просвет . [8]
Некоторые таксоны Trichoptera ( ручейники), которые являются сестринской группой Lepidoptera, имеют волосообразные чешуи, но всегда на крыльях и никогда на теле или других частях насекомого. [13] У ручейников также имеются хвостовые церки на брюшке, чего нет у чешуекрылых. [7] По словам Скобла (2005), [8] «морфологически чешуя представляет собой макротрихию и, таким образом, гомологична большим волоскам (и чешуйкам), покрывающим крылья Trichoptera (ручейников)».
Структура
[ редактировать ]Несмотря на большое разнообразие масштабов, все они имеют схожую структуру. Чешуя, как и у других макрохет , возникает из особых трихогенных (волосообразующих) клеток и имеет гнездо, заключенное в особую «тормогенную» клетку; [15] : 9 такое расположение обеспечивает стебель или цветоножку , с помощью которой чешуи прикрепляются к субстрату. Чешуя может быть волосистой (волосистой) или уплощенной. Тело или «лезвие» типичной уплощенной чешуи состоит из верхней и нижней пластинок с воздушным пространством между ними. Поверхность по направлению к телу гладкая и известна как нижняя пластинка. Верхняя поверхность, или верхняя пластинка, имеет поперечные и продольные гребни и ребра. Ламели разделены распорками, называемыми трабакулами , и содержат пигменты, придающие цвет. Чешуйки неплотно прилегают к крылу и легко отрываются, не причиняя вреда бабочке. [8] [13] [39]
Цвет
[ редактировать ]Чешуйки на крыльях бабочки пигментированы меланином , который придает ей черный и коричневый цвета. Белый цвет у бабочек семейства Pieridae — производное мочевой кислоты , продукт выведения. [13] [40] : 84 Яркие синие, зеленые, красные цвета и переливы обычно создаются не пигментами, а микроструктурой чешуек. Эта структурная окраска является результатом когерентного рассеяния света фотонно-кристаллической природой чешуек. [41] [42] [43] Специализированные чешуйки, которые обеспечивают структурные цвета отраженного света, в основном создают ультрафиолетовые узоры, которые различимы в той части ультрафиолетового спектра, которую могут видеть глаза чешуекрылых. [13] Видимый структурный цвет часто зависит от угла зрения. Например, у Morpho cypris цвет спереди ярко-синий, но если смотреть под углом, он очень быстро меняется на черный. [44]
радужная структурная окраска крыльев многих видов ликанид и папилионид, таких как Parides sesostris и Teinopalpus Imperialis , а также ликанид, таких как Callophrys Rubi , Cyanophrys remus и Mitoura gryneus . Была изучена [45] Они представляют собой наиболее сложную из известных архитектур фотонного масштаба – регулярные трехмерные периодические решетки, которые встречаются в просвете некоторых масштабов. [46] В случае с Кайзер-и-Хиндом ( Teinopalpus Imperialis ) трехмерная фотонная структура была исследована с помощью трансмиссионной электронной томографии и компьютерного моделирования, чтобы выявить встречающиеся в природе «хиральные тетраэдрические повторяющиеся единицы, упакованные в триклинную решетку». [47] [48] причина радужности.
 Структурный синий цвет у morpho cypris , нимфалиды.
Структурный синий цвет у morpho cypris , нимфалиды. тот же экземпляр Morpho cypris с конца, синий цвет становится черным. Если увидеть
тот же экземпляр Morpho cypris с конца, синий цвет становится черным. Если увидеть Белый цвет у околоцветников, таких как Delias eucharis, является производным мочевой кислоты, продуктом выведения.
Белый цвет у околоцветников, таких как Delias eucharis, является производным мочевой кислоты, продуктом выведения. Зеленое переливание парусника Кайзер-и-Хинд ( Teinopalpus Imperialis ) привело к открытию трехмерной фотонно-кристаллической структуры.
Зеленое переливание парусника Кайзер-и-Хинд ( Teinopalpus Imperialis ) привело к открытию трехмерной фотонно-кристаллической структуры. Окраска крыльев у некоторых чешуекрылых позволяет маскироваться, как это можно увидеть на примере бабочки-геометриды Colostygia aqueata .
Окраска крыльев у некоторых чешуекрылых позволяет маскироваться, как это можно увидеть на примере бабочки-геометриды Colostygia aqueata .





Функция
[ редактировать ]

Чешуя играет важную роль в естественной истории чешуекрылых. Чешуя позволяет развивать яркие или нечеткие узоры , которые помогают организму защитить себя путем маскировки , мимикрии и предупреждения. Помимо обеспечения изоляции , темные узоры на крыльях позволяют поглощать солнечный свет и, вероятно, участвуют в терморегуляции . Яркие и характерные цветовые узоры бабочек, которые неприятны хищникам, помогают сообщить об их токсичности или несъедобности, тем самым предотвращая нападение хищников. В бейтсовской мимикрии рисунок цвета крыльев помогает съедобным чешуекрылым имитировать несъедобные модели, в то время как в мюллеровской мимикрии несъедобные бабочки напоминают друг друга, чтобы уменьшить количество особей, отбираемых неопытными хищниками. [8]
Возможно, чешуя изначально возникла для обеспечения изоляции. Чешуя на грудной клетке и других частях тела может способствовать поддержанию высокой температуры тела, необходимой во время полета. Однако «твердые» чешуи базальных бабочек не так эффективны, как у их более продвинутых родственников, поскольку наличие просвета добавляет воздушные слои и увеличивает изоляционную ценность. [8] Весы также помогают увеличить соотношение подъемной силы и лобового сопротивления в полете. [8]
Для недавно появившихся взрослых особей большинства мирмекофильных Lycaenidae лиственные восковые чешуйки обеспечивают некоторую защиту от хищников, когда они выходят из гнезда. [8] В случае с бабочкой-ножницей ( Liphyra brassolis ) гусеницы — нежеланные гости в гнездах древесных муравьев, питающиеся личинками муравьев. Взрослые особи, выходящие из куколок, покрыты мягкими рыхлыми клейкими чешуйками, которые стираются и прилипают к муравьям, когда они выбираются из гнезда после вылупления. [49]

Андрокония
[ редактировать ]Самцы чешуекрылых обладают особыми чешуйками, называемыми андрокониями (единственное число — androconium ), которые развились в результате полового отбора с целью распространения феромонов для привлечения подходящих самок. Андрокония может быть рассеяна по крыльям, телу или ногам или встречаться в виде пятен, называемых «таврами», «половыми клеймами» или «стигматами» на крыльях, обычно в впячиваниях верхней поверхности передних крыльев, иногда скрытых другие весы. Известно также, что Androconia встречается в складках крыльев. Эти марки иногда состоят из волосоподобных пучков, которые облегчают распространение феромона. роль андроконии в ухаживании бабочек-пиерид и нимфалид, таких как Pyronia tithonus и Dryas iulia . Экспериментально доказана [15] : 16–17 [50] [51] [52] [53]
Последовательные крупные планы чешуек павлиньего крыла.
Фотографические и светомикроскопические изображения Уменьшенный вид Aglais io . Крупный план чешуи того же экземпляра. Большое увеличение цветных чешуек (вероятно, другого вида). Электронно-микроскопические изображения Пятно крыла Весы крупным планом Единая шкала Микроструктура чешуи Увеличение Прибл. ×50 Прибл. ×200 ×1000 ×5000







Живот
[ редактировать ]Брюшко или тело состоит из девяти сегментов. У личинки оно варьируется от 5 до 13 сегментов. Одиннадцатый сегмент личинки содержит пару анальных зажимов, которые у некоторых таксонов выступают вперед и представляют собой гениталии. [11]
Многие семейства бабочек имеют специальные органы, помогающие обнаруживать эхолокацию летучих мышей . Эти органы известны как барабанная перепонка (единственное число – typanum ). и У Pyraloidea почти у всех Geometroidea барабанная перепонка расположена на переднем стерните брюшка. также У Noctuoidea есть барабанные перепонки, но у них барабанные перепонки расположены на нижней стороне заднегруди , строение и положение которых уникальны и являются таксономическим отличительным признаком надсемейства. [21] [54]
У самок некоторых бабочек на кончике брюшка расположен орган, выделяющий запах. [6]
Гениталии
[ редактировать ]
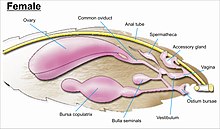
Гениталии . сложны и служат основой для видовой дискриминации в большинстве семей, а также при идентификации семьи [7] Гениталии возникают из десятого или самого дистального сегмента живота. Чешуекрылые имеют одни из самых сложных генитальных структур среди всех насекомых, с большим разнообразием сложных шипов, щетинок, чешуек и пучков у самцов, классеров различной формы и модификаций протоков у самок, через которые запасенная сперма переносится внутри самку прямо или косвенно во влагалище для оплодотворения. [13] [55] [56]
Расположение гениталий важно при ухаживании и спаривании, поскольку оно предотвращает межвидовое спаривание и гибридизацию. Уникальность гениталий вида привела к использованию морфологического изучения гениталий как одного из важнейших ключей при таксономической идентификации таксонов ниже уровня семейства. С появлением анализа ДНК изучение гениталий стало лишь одним из методов, используемых в систематике. [6]
У большинства чешуекрылых различают три основные конфигурации гениталий, основанные на том, как развивалось расположение у самок отверстий для совокупления, оплодотворения и откладки яиц:
- Экзопорий : Hepialidae и родственные семейства имеют внешнюю бороздку, по которой сперматозоиды переносятся из копулятивного отверстия (гонопора) в (яйцепор), и называются экзопориями. [7]
- Монотризийские : примитивные группы имеют единственное генитальное отверстие в конце брюшка, через которое происходит как совокупление, так и откладка яиц. Этот символ используется для обозначения Монотрисии. [7]
- Дитризии : Остальные группы имеют внутренние протоки, по которым переносится сперма и образуют дитризии, с отдельными отверстиями для совокупления и откладки яиц. [7]
Гениталии самца и самки у любого конкретного вида приспособлены друг к другу, как замок (самец) и ключ (самка). [6] У самцов девятый брюшной сегмент разделен на дорсальный «тегумен» и вентральный «виникулум». [56] Они образуют кольцевидную структуру для прикрепления половых частей и пару латеральных обхватывающих органов (класперов или «гарп»). У самца есть срединный трубчатый орган (называемый эдеагусом ) , который проходит через выворачивающуюся оболочку (или «везику») для оплодотворения самки. [55] У всех чешуекрылых самцы имеют парные семяпроводы; парные семенники у базальных таксонов раздельные, у продвинутых форм слиты. [55]
Если расположение внутренних половых протоков и отверстий женских половых органов зависит от таксономической группы, к которой принадлежит насекомое, то внутренняя женская репродуктивная система всех чешуекрылых состоит из парных яичников и дополнительных желез, вырабатывающих желтки и скорлупу яиц. У самок насекомых есть система сосудов и протоков, в которых сперма принимается, транспортируется и хранится. Яйцеводы самки соединяются, образуя общий проток (называемый «яйцеводом коммунис»), который ведет во влагалище. [55] [56]
Когда происходит совокупление, самец бабочки или мотылька помещает капсулу со спермой ( сперматофор ) в сосуд самки (называемый бурсой тела ). Сперматозоиды, высвобождаемые из капсулы, плывут непосредственно или через небольшую трубку в специальный семеприемник ( сперматека ), где сперма хранится до тех пор, пока она не выйдет во влагалище для оплодотворения во время откладки яйцеклетки, что может происходить часами, днями. или месяцев после спаривания. Яйца проходят через яйцепор . Яйцепор может находиться на конце модифицированного яйцеклада или окружен парой широких щетинкообразных анальных сосочков. [55] [56]
Бабочки Parnassinae ( семейство Papilionidae) и некоторые Acraeini посткопулятивную пробку, называемую сфрагисом , что предотвращает ее повторное спаривание. (семейство Nymphalidae) добавляют к брюшку самки после совокупления [13]
Самцы многих видов Papilionoidea обладают вторичными половыми признаками. Они состоят из органов, производящих аромат, кисточек и клейм или мешочков со специальными чешуйками. По-видимому, они выполняют функцию убеждения самки в том, что она спаривается с самцом нужного вида. [11]
Было зарегистрировано, что три вида бражников издают ультразвуковые щелчки, потирая свои гениталии; самцы издают звук, потирая жесткие чешуи на внешней стороне застежек, а самки издают звук, сокращая гениталии, что вызывает трение чешуек о живот. Функция этого шума не ясна, и выдвигаемые предложения включают в себя подавление эхолокации летучих мышей и рекламу того, что добыча летучих мышей - колючие и отличные летуны. [57]
 Citheronia regalis с закрытыми пряжками
Citheronia regalis с закрытыми пряжками Citheronia regalis с открытыми пряжками
Citheronia regalis с открытыми пряжками
 Крупный план: затвердевшая сфрагиса, выступающая на 2–3 мм позади брюшка Парнаса.
Крупный план: затвердевшая сфрагиса, выступающая на 2–3 мм позади брюшка Парнаса.




Клоака
[ редактировать ]Чешуекрылые насекомые имеют клоаку на конце брюшка. Он может быть полным, включая анус , яйцепор и копулятивную пору , как в случае Dacnonypha , Zeugloptera и большинства Monotrysia ; или неполный, включающий только анус и яйцепор, как встречается у некоторых Monotrysia, Psychidae , а также у некоторых Choreutidae и Cossidae . [58]
Разработка
[ редактировать ]
Оплодотворенное яйцо созревает и из него вылупляется гусеница. Гусеница — это питательная стадия жизненного цикла чешуекрылых. Гусеница должна иметь возможность питаться и избегать быть съеденной, и большая часть ее морфологии эволюционировала для облегчения этих двух функций. [59] : 108 После роста и шелушения гусеница вступает в сидячую стадию развития, называемую куколкой (или куколкой), вокруг которой она может образовывать кожух. Насекомое развивается во взрослую особь на стадии куколки; Когда все готово, куколка вылупляется и возникает взрослая стадия или имаго бабочки или мотылька.
Яйцо
[ редактировать ]
Как и большинство насекомых, чешуекрылые являются яйцекладущими или «яйцекладущими». [40] Яйца чешуекрылых, как и яйца других насекомых, являются центролецитальными , поскольку яйца имеют центральный желток , окруженный цитоплазмой . Желток обеспечивает жидкое питание зародыша- гусеницы до тех пор, пока он не выйдет из скорлупы. [60] Цитоплазма окружена желточной оболочкой , а белковая мембрана, называемая хорионом, защищает яйцеклетку снаружи. [60] [61] Ядро зиготы расположено сзади. [61]
У некоторых видов чешуекрылых внутри хориона, примыкающего к желточному слою, присутствует восковой слой, который, как полагают, возник в ходе эволюции для предотвращения высыхания. У насекомых хорион имеет слой воздушных пор в твердом материале, который обеспечивает очень ограниченную способность к дыхательной функции. У чешуекрылых слой хориона над слоем воздушных пор имеет пластинчатую форму с последовательными листами белка, расположенными в определенном направлении и ступенчатыми, образуя спиральную структуру. [61]
Верхняя часть яйцеклетки вдавлена и образует небольшую центральную полость, называемую микропиле, через которую яйцеклетка оплодотворяется. [11] Микропиле расположено сверху у яиц шаровидной, конической или цилиндрической формы; у уплощенных или линзовидных яиц микропиле расположено по наружному краю или кайме. [18] [62]
Яйца чешуекрылых обычно округлые и маленькие (1 мм), хотя у Sphingidae и Saturniidae они могут достигать 4 мм . [13] : 640 Обычно они довольно простого цвета: белого, бледно-зеленого, голубовато-зеленого или коричневого цвета. Яйца бабочек и мотыльков бывают разной формы; некоторые сферические, другие полусферические, конические, цилиндрические или двояковыпуклые (в форме линзы). Некоторые имеют форму бочонка или блина, а другие имеют форму тюрбана или сыра. Они могут быть угловатыми или вдавленными на обоих концах, ребристыми или орнаментированными, пятнистыми или испорченными. [18] [62]
Яйца откладываются поодиночке, небольшими группами или массой и всегда на источнике пищи или рядом с ним. Известно, что пленные бабочки откладывают яйца в клетках, в которых они были изолированы. [18] [62] На размер яиц чешуекрылых влияет ряд факторов. Виды чешуекрылых, зимующие на стадии яйца, обычно имеют более крупные яйца, чем виды, у которых этого нет. Точно так же виды, питающиеся древесными растениями на личиночной стадии, имеют более крупные яйца, чем виды, питающиеся травянистыми растениями. Было отмечено, что яйца, отложенные старшими самками некоторых видов бабочек, меньше по размеру, чем их более молодые собратья. при отсутствии адекватного питания самки кукурузной мотыльки ( Ostrinia spp.) откладывают кладки с размерами яиц ниже нормы. Зарегистрировано, что [61]
Во время побега только что вылупившиеся личинки многих видов иногда поедают хорион, чтобы выйти на свет. Альтернативно, скорлупа яйца может иметь слабую линию вокруг шляпки, которая дает возможность выйти личинке. [61] Яичная скорлупа и небольшое количество желтка, попавшие в околоплодные оболочки, составляют первую пищу для большинства личинок чешуекрылых.
 Яйца пионера ( Anaphaeis aurota семейства Pieridae)
Яйца пионера ( Anaphaeis aurota семейства Pieridae) Яйца малиновой розы ( Atropaneura hector семейства Papilionidae)
Яйца малиновой розы ( Atropaneura hector семейства Papilionidae) Яйцо мальвы шкипера ( Carcharodus alceae, семейство Hesperiidae)
Яйцо мальвы шкипера ( Carcharodus alceae, семейство Hesperiidae) Яйцо большое медное ( Lycaena dispar семейство Lycaenidae)
Яйцо большое медное ( Lycaena dispar семейство Lycaenidae)




 Расположенные рядом яйца двукрылых чешуекрылых, кипариса листовертки ( Archips goyerena семейства Tortricidae )
Расположенные рядом яйца двукрылых чешуекрылых, кипариса листовертки ( Archips goyerena семейства Tortricidae ) Вертикальные яйца двукрылых чешуекрылых, лунной бабочки ( семейство Actias luna Saturniidae), отложенные в неволе на бумаге
Вертикальные яйца двукрылых чешуекрылых, лунной бабочки ( семейство Actias luna Saturniidae), отложенные в неволе на бумаге Яйца сосновой плодожорки ( Bupalus piniaria семейства Geometridae)
Яйца сосновой плодожорки ( Bupalus piniaria семейства Geometridae) Яйца лакуньской моли ( Malacosoma neustria семейства Lasiocampidae)
Яйца лакуньской моли ( Malacosoma neustria семейства Lasiocampidae)




Гусеница
[ редактировать ]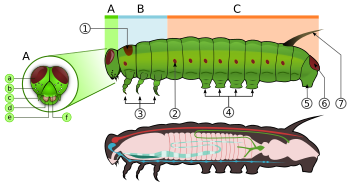
Гусеницы - это «характерные личинки -полиподы с цилиндрическим телом, короткими грудными ногами и брюшными ложноногами (псевдоподами)». [63] У них упрочненная ( склеротизированная ) головная капсула, челюсти (ротовой аппарат) для жевания и мягкое трубчатое сегментированное тело, которое может иметь волосообразные или другие выступы, три пары настоящих ног и дополнительные ложноножки (до пяти пар). . [2] Тело состоит из тринадцати сегментов, из которых три грудных (Т1, Т2 и Т3) и десять брюшных (от А1 до А10). [21]




У всех настоящих гусениц есть перевернутая Y-образная линия, идущая от макушки головы вниз. Между Y-образной линией лежит лобный треугольник или лоб. Наличник двумя , расположенный ниже лба, лежит между усиками . находится Верхняя губа под наличником. В центре верхней губы есть небольшая выемка, с которой соприкасается край листа, когда гусеница ест. [64]
У личинок имеются шелковые железы, расположенные на губах . Эти железы представляют собой видоизмененные слюнные железы . Они используют эти шелковые железы для изготовления шелка для коконов и укрытий. [21] Ниже верхней губы расположены нижние челюсти. [64] На каждой стороне головы обычно имеется шесть стемм чуть выше мандибул . Эти стемматы расположены полукругом. Под стеммами имеется небольшая пара усиков, по одному с каждой стороны. [21] [64]
Грудная клетка несет три пары ног, по одной паре на каждом сегменте. Переднегруди (Т1) имеет функциональное дыхальце, которое фактически происходит от среднегруди (Т2), тогда как заднегруди имеет уменьшенное дыхальце, которое не открыто снаружи и лежит под кутикулой. [59] : 114 Грудные ноги состоят из тазика, вертела, бедренной кости, предплюсны и коготка и имеют постоянную форму на протяжении всего отряда. Однако у некоторых минаторов они редуцированы, а у некоторых Notodontidae — удлинены . У Micropterigidae ноги трехчлениковые, так как тазик, вертел и бедренная кость срослись. [59] : 114
Каждый из брюшных сегментов с третьего по шестой и десятый может нести по паре более мясистых ног. [21] Грудные ноги называются настоящими ногами, а брюшные ноги — ложными. [64] Настоящие ноги у чешуекрылых мало различаются, за исключением редукции у некоторых минеров и удлинения у семейства Notodontidae . [59] : 114 Ложные ножки имеют на кончике несколько маленьких крючков, которые называются крючками. Семейства чешуекрылых различаются количеством и расположением ложноножек. Некоторые личинки, такие как дюймовые черви ( Geometridae ) и петлители ( Plusiinae ), имеют пять пар ложноножек или меньше, в то время как другие, такие как Lycaenidae и гусеницы слизней ( Limacodidae ), вообще не имеют ложноножек. [21] [64] У некоторых гусениц, добывающих листья, на брюшной стенке имеются петли, представляющие собой уменьшенные ложноножки, в то время как у других видов, добывающих листья, петли полностью отсутствуют. [65] Брюшные дыхальца расположены с каждой стороны тела на первых восьми брюшных сегментах. [64]
Гусеницы имеют разные типы выступов; щетинки (волосы), шипы, бородавки, бугорки и рога. Волосы бывают разных цветов и могут быть длинными или короткими; одиночные, группами или пучками; тоньше на конце или булавовидный на конце. Позвоночник может быть халазой (с одной точкой) или сколом (с несколькими точками). Бородавки могут представлять собой небольшие шишки или короткие выступы на теле. Бугорки представляют собой мясистые выступы тела, которые либо короткие и похожие на бугорки, либо длинные и похожие на нити. Обычно они встречаются парами или скоплениями на одном или нескольких сегментах. Рога короткие, мясистые, заостренные. Обычно они располагаются на восьмом брюшном сегменте. [65]
У большого количества видов семейств Saturniidae , Limacodidae и Megalopygidae есть жалящие гусеницы с ядовитыми щетинками, называемыми уртикирующими волосками , а в случае Lonomia — бразильского рода сатурнийид — могут убить человека из-за своего мощного антикоагулянтного яда. [13] : 644 Гусеницы многих таксонов, которые изолировали токсичные химические вещества от растений-хозяев или имеют острые крапивные волосы или шипы, демонстрируют апосематическую окраску и отметки. [66]
Гусеницы подвергаются шелушению и имеют несколько личиночных возрастов , обычно пять, но у разных видов они различаются. Новая кутикула мягкая и позволяет гусенице увеличиваться в размерах и развиваться, прежде чем она станет твердой и неэластичной. При последнем шелушении старая кутикула расщепляется и скручивается в небольшой шарик на заднем конце куколки и называется личиночной экзувией . [67] : 31
 Два возраста папилионида обыкновенного мормона с разными камуфляжными схемами, напоминающими птичий помет и растительность.
Два возраста папилионида обыкновенного мормона с разными камуфляжными схемами, напоминающими птичий помет и растительность. Личинки нотодонтидных бабочек, таких как Stauropus fagi , имеют удлиненные грудные ноги.
Личинки нотодонтидных бабочек, таких как Stauropus fagi , имеют удлиненные грудные ноги. Личинка Lonomia obliqua , сатурниид из Бразилии, имеет крапивные волоски , содержащие смертельный яд -антикоагулянт .
Личинка Lonomia obliqua , сатурниид из Бразилии, имеет крапивные волоски , содержащие смертельный яд -антикоагулянт . Личинки седловидной моли ( Acharia стимулеа ) имеют апосематическую окраску в форме седла.
Личинки седловидной моли ( Acharia стимулеа ) имеют апосематическую окраску в форме седла. Нижняя сторона гусениц слизней Phobetron pithecium (семейство Limacododiae) с отсутствием ложноножек.
Нижняя сторона гусениц слизней Phobetron pithecium (семейство Limacododiae) с отсутствием ложноножек. Гусеница осины минёрки ( Phyllocnistis populiella )
Гусеница осины минёрки ( Phyllocnistis populiella ) Побег красного дерева ( Hypsipyla grandella ) повреждает красное дерево в Бразилии.
Побег красного дерева ( Hypsipyla grandella ) повреждает красное дерево в Бразилии. Гусеница мешочника (возможно, Hyalarcta huebneri семейства Psychidae ), выходящая из футляра
Гусеница мешочника (возможно, Hyalarcta huebneri семейства Psychidae ), выходящая из футляра Последний возраст синей мормонской растительности, напоминающей личинку.
Последний возраст синей мормонской растительности, напоминающей личинку.









Куколка или куколка
[ редактировать ]


Кокон представляет собой оболочку, сплетенную из шелка многими гусеницами моли и многочисленными другими голометаболических насекомых личинками в качестве защитного покрытия для куколки. Большинство личинок чешуекрылых либо образуют кокон и окукливаются внутри него, либо окукливаются в ячейке под землей. [21] за исключением бабочек и продвинутых бабочек, таких как совки , куколки которых обнажены. [13] Куколки бабочек обычно коричневые и гладкие, тогда как куколки бабочек часто разноцветные и их форма сильно различается. [21] У бабочек обнаженную куколку часто называют куколкой , что происходит от греческого термина «куколка»: χρυσός ( chrysós ) — золото , имея в виду золотистый цвет некоторых куколок. [68]
Гусеницы многих бабочек прикрепляются шелковой пуговицей к нижней стороне ветки, камня или другой выступающей поверхности. Они остаются прикрепленными к шелковой подушечке с помощью крючкообразного процесса, называемого кремастером. Большинство куколок свисают головой вниз, но в семействах Papilionidae, Pieridae и Lycaenidae куколка удерживается в более вертикальном положении с помощью шелкового пояса вокруг середины куколки. [21]
Куколки большинства чешуекрылых закрытые , со сросшимися или приклеенными к телу придатками, а у остальных куколки экзаратные , у них усики, ноги и крылья свободны и не приклеены к телу. [69]
На стадии куколки морфология взрослой особи развивается путем развития личиночных структур. [40] : 151 Общий вид взрослой особи виден до того, как затвердеет внешняя поверхность: голова, опирающаяся на грудную клетку, глаза, усики (выдвинутые вперед над головой), крылья, приложенные к грудной клетке, и шесть ног между крыльями и живот. [70] Среди особенностей, различимых в области головы куколки, - склериты, швы, пилиферы , нижние челюсти, окуляры, усики, щупики и верхние челюсти. Куколочная грудная клетка состоит из трех грудных сегментов, ног, крыльев, тегул, крыловых борозд и подмышечных бугорков. На брюшке куколки имеются десять сегментов, шипы, щетинки, рубцы личиночных ложноножек и бугорков, анальное и половое отверстия, а также дыхалец. У куколок бурильщиков имеются фланцевые пластинки, а у куколок специализированных чешуекрылых — кремастер. [14] : 23–29
Хотя куколка обычно неподвижна и неподвижна, у примитивных бабочек семейств Micropterigidae , Agathiphagidae и Heterobathmiidae есть полностью функциональные челюсти. [59] : 131 В основном они служат для того, чтобы взрослая особь могла выбраться из кокона. [14] : 34 Кроме того, все придатки и тело отделены от кожи куколки и обладают определенной степенью независимого движения. Все остальные надсемейства чешуекрылых более специализированы, имеют нефункциональные челюсти, придатки и тело, прикрепленные к коже куколки, и теряют некоторую степень самостоятельности движений. [14] : 20
Куколки некоторых бабочек способны извивать брюшко. Три каудальных сегмента брюшка куколки (сегменты 8–10) фиксированы; остальные сегменты в некоторой степени подвижны. В то время как более развитые Lepidoptera могут извиваться только последние два или три сегмента на конце брюшка, более базальные таксоны, такие как Micropterigidae, могут извивать оставшиеся семь сегментов брюшка; это, по-видимому, помогает им высовывать передний конец из куколочного чехла перед выходом из куколки . [14] : 28 [67] Куколки Hepialidae способны перемещаться вперед и назад по личиночному туннелю, извиваясь, чему способствуют не только шипы, но и выступы на спине. [67] Считается, что вращение живота пугает хищников и отпугивает их. У некоторых бражников , таких как Theretra latreillii , вращение брюшка сопровождается дребезжащим или щелкающим звуком, что усиливает эффект испуга. [67]
У некоторых видов, например Heliconius charithonia , спаривание может происходить внутри куколки самок с самцами. [71]
 Куколки папилионид обычно прикрепляются к субстрату кремастером, а голова удерживается шелковым поясом.
Куколки папилионид обычно прикрепляются к субстрату кремастером, а голова удерживается шелковым поясом. Подвесная куколка нимфалиды золотистого цвета ядра Euploea.
Подвесная куколка нимфалиды золотистого цвета ядра Euploea. Actias luna (семейство Saturniidae ) выходит из кокона
Actias luna (семейство Saturniidae ) выходит из кокона Специализированная куколка сфингидной моли, Agrius convolvuli , может извивать брюшком, издавая щелкающий звук, что может вызывать эффект испуга.
Специализированная куколка сфингидной моли, Agrius convolvuli , может извивать брюшком, издавая щелкающий звук, что может вызывать эффект испуга.



Защита и хищничество
[ редактировать ]


Чешуекрылые имеют мягкое тело, хрупкие и почти беззащитные, в то время как неполовозрелые стадии движутся медленно или неподвижны, следовательно, все стадии подвергаются хищничеству со стороны птиц, мелких млекопитающих, ящериц, земноводных, беспозвоночных хищников (особенно паразитоидных и паразитических ос и мух ). как грибы и бактерии. Чтобы бороться с этим, чешуекрылые разработали ряд стратегий защиты и защиты, которые включают камуфляж , апосематизм , мимикрию , а также развитие моделей и проявлений угрозы. [72]
Камуфляж — важная стратегия защиты, обеспечиваемая изменениями формы, цвета и маркировки тела. Некоторые чешуекрылые сливаются с окружающей средой, что затрудняет их обнаружение хищниками. Гусеницы могут иметь оттенки зеленого, соответствующие растению-хозяину. Другие напоминают несъедобные предметы, например, ветки или листья. Личинки некоторых видов, таких как обыкновенный мормон и западный тигровый махаон, похожи на птичий помет. [72] [73]
Некоторые виды чешуекрылых связывают или производят токсины, которые хранятся в тканях их тела, что делает их ядовитыми для хищников; примеры включают бабочку-монарх в Америке и виды Aтрофаневры в Азии. Хищники, поедающие ядовитых чешуекрылых, могут заболеть и вызвать сильную рвоту, поэтому им приходится избегать этих видов. Хищник, который ранее съел ядовитого чешуекрылого, может в будущем избегать других видов с похожими отметинами, тем самым спасая и многие другие виды. [72] [74] Токсичные бабочки и личинки имеют тенденцию приобретать яркую окраску и яркие узоры, что служит индикатором для хищников их токсичности. Это явление известно как апосематизм . [75]
Апосематизм также привел к развитию комплексов мимикрии бейтсовской мимикрии , где съедобные виды имитируют апосематические таксоны, и мюллеровой мимикрии , где несъедобные виды, часто принадлежащие к родственным таксонам, эволюционировали, чтобы походить друг на друга, чтобы извлечь выгоду из снижения частоты выборки за счет хищники во время обучения. Точно так же взрослые виды Sesiidae (также известные как бабочки-ясликрылки) имеют общий внешний вид, достаточно похожий на осу или шершня , что делает вероятным, что бабочки уменьшат хищничество за счет мимикрии Бейтса. [76]
Глазные пятна — это тип автомимикрии, используемый некоторыми чешуекрылыми. У бабочек пятна состоят из концентрических колец чешуек разного цвета. Предполагаемая роль глазных пятен — отвлекать внимание хищников. Их сходство с глазами провоцирует инстинкт хищника атаковать эти рисунки крыльев. [77] Было высказано предположение, что роль нитевидных хвостов у Lycaenidae сбивает хищников с толку относительно реального местоположения головы, что дает им больше шансов спастись живыми и относительно невредимыми. [78]
Некоторые гусеницы, особенно представители Papilionidae, содержат осметрий Y-образной формы, — выступающую железу обнаруженную в переднегрудном сегменте личинок. При угрозе гусеница испускает неприятные запахи из органа, чтобы отогнать хищников. [79] [80]
См. также
[ редактировать ]- Различия между бабочками и мотыльками
- Словарь энтомологических терминов
- Морфология насекомых
- Чешуекрылые
- Морфология (биология)
Сноски
[ редактировать ]- ^ Кристенсен, Нильс П.; Скобл, MJ; Каршольт, Оле (2007). З.-К. Чжан; В.А. Шир (ред.). Трехсотлетие Линнея: прогресс в таксономии беспозвоночных (PDF) . Том. 1668. стр. 699–747. дои : 10.11646/zootaxa.1668.1.30 . ISBN 978-0-12-690647-9 . S2CID 4996165 . Архивировано из оригинала (PDF) 15 мая 2013 года . Проверено 19 февраля 2011 г.
Глава: «Филогения и систематика чешуекрылых: состояние инвентаризации разнообразия бабочек и бабочек»
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Jump up to: а б Дагдейл, Дж. С. (1996). «Естественная история и идентификация питающихся подстилкой личинок Lepidoptera (Insecta) в буковых лесах, долина Оронгоронго, Новая Зеландия, с особым упором на рацион мышей ( Mus musculus )» (PDF) . Журнал Королевского общества Новой Зеландии . 26 (4): 251–274. Бибкод : 1996JRSNZ..26..251D . дои : 10.1080/03014223.1996.9517513 . [ постоянная мертвая ссылка ]
- ^ Jump up to: а б с д Скобл, MJ (1995). «Ротовой аппарат» . Чешуекрылые: форма, функции и разнообразие . Издательство Оксфордского университета . стр. 6–19. ISBN 978-0-19-854952-9 .
- ^ Боррор, Дональд Дж.; Триплхорн, Чарльз А.; Джонсон, Норман Ф. (1989). Введение в изучение насекомых (6, иллюстрированное изд.). Публикации колледжа Сондерс. ISBN 978-0-03-025397-3 . Проверено 16 ноября 2010 г. (Без предварительного просмотра.)
- ^ Jump up to: а б с Скобл (1995). Раздел «Сенсация», (с. 26–38).
- ^ Jump up to: а б с д и ж г час Хоскинс, Адриан. «Анатомия головы бабочки (и другие страницы)» . Узнайте о бабочках . Проверено 15 ноября 2010 г.
- ^ Jump up to: а б с д и ж г час я дж к л м н Пауэлл, Джерри А. (2009). «Чешуекрылые» . В Реше, Винсент Х.; Карде, Ринг Т. (ред.). Энциклопедия насекомых (2-е изд.). Академическая пресса . стр. 661–663. ISBN 978-0-12-374144-8 .
- ^ Jump up to: а б с д и ж г час я Скобл (1995). Раздел «Весы», (стр. 63–66).
- ^ Маллет, Джим (12 июня 2007 г.). «Подробные сведения о проектах таксом чешуекрылых и бабочек» . Проект таксомы чешуекрылых . Университетский колледж Лондона . Проверено 14 ноября 2010 г.
- ^ Jump up to: а б с Жилло, Седрик (1995). «Бабочки и мотыльки» . Энтомология (2-е изд.). Спрингер. ISBN 978-0-306-44967-3 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т Эванс, WH (1932). "Введение" . Идентификация индийских бабочек (2-е изд.). Мумбаи: Бомбейское общество естествознания . стр. 1–35.
- ^ «Чешуекрылые» . Британская энциклопедия . 2011 . Проверено 12 февраля 2011 г.
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с Хеппнер, Дж. Б. (2008). «Бабочки и мотыльки» . В Капинере, Джон Л. (ред.). Энциклопедия энтомологии . Виртуальная справочная библиотека Гейла. Том. 4 (2-е изд.). Справочник Спрингера. п. 4345. ИСБН 978-1-4020-6242-1 .
- ^ Jump up to: а б с д и ж Мошер, Эдна (2009) [1918]. Классификация чешуекрылых на основе признаков куколки (переиздание). БиблиоБазар, ООО. ISBN 978-1-110-02244-1 .
- ^ Jump up to: а б с д и Кристенсен, Нильс П. (2003). Чешуекрылые, мотыльки и бабочки: морфология, физиология и развитие, Том 2 . Том 4, часть 36 Справочника по зоологии. Вальтер де Грюйтер . ISBN 978-3-11-016210-3 .
- ^ Jump up to: а б с д Скобл (1995). Раздел «Голова взрослого человека – питание и ощущения» (стр. 4–22).
- ^ Хеппнер, Джон Б. (2008). «Шелкопряды (Lepidoptera: Bombycidae)». В Капинере, Джон Л. (ред.). Энциклопедия энтомологии Спрингер Нидерланды. стр. 100-1 3375–3376. дои : 10.1007/978-1-4020-6359-6_4198 . ISBN 9781402062421 .
- ^ Jump up to: а б с д Голландия, WJ (1903). «Введение» ( PDF ) . Книга Мотылька . Лондон: ISBN Хатчинсона и Ко. 978-0-665-75744-0 .
- ^ Мерлин, Кристина; Гегер, Роберт Дж.; Репперт, Стивен М. (2009). «Антеннальные циркадные часы координируют ориентацию солнечного компаса у мигрирующих бабочек-монархов» . Наука . 325 (5948): 1700–1704. Бибкод : 2009Sci...325.1700M . дои : 10.1126/science.1176221 . ПМЦ 2754321 . ПМИД 19779201 .
- ^ Робинсон, GS (1988). «Филогения Tineoidea (Lepidoptera)». Систематика и эволюция насекомых . 19 (2). Брилл: 117–129. дои : 10.1163/187631289x00113 . . «...у многих амфитерид (sl) сложный глаз самцов частично или полностью разделен горизонтально."
- ^ Jump up to: а б с д и ж г час я дж к Триплхорн, Чарльз А.; Джонсон, Норман Ф. (2005). Введение Боррора и Делонга в изучение насекомых . Бельмонт, Калифорния: Томсон Брукс/Коул. ISBN 978-0-03-096835-8 .
- ^ Агоста, Сальваторе Дж.; Янзен, Дэниел Х. (2004). «Распределение размеров тела крупных коста-риканских сухих лесных бабочек и основная связь между морфологией растений и опылителей». Ойкос . 108 (1): 183–193. дои : 10.1111/j.0030-1299.2005.13504.x .
- ^ Кунте, Крушнамег (2007). «Аллометрия и функциональные ограничения на длину хоботков бабочек» . Функциональная экология . 21 (5): 982–987. Бибкод : 2007FuEco..21..982K . дои : 10.1111/j.1365-2435.2007.01299.x .
- ^ Кренн, Х.В.; Пенц, CM (1 октября 1998 г.). «Ротовой аппарат бабочек Heliconius (Lepidoptera: Nymphalidae): поиск анатомических адаптаций к поведению при питании пыльцой». Международный журнал морфологии и эмбриологии насекомых . 27 (4): 301–309. дои : 10.1016/S0020-7322(98)00022-1 .
- ^ Маккензи, Дебора (20 декабря 2006 г.). «Мотыльки пьют слезы спящих птиц» . Новый учёный . Деловая информация Рида . Проверено 10 февраля 2012 г.
- ^ Хилгартнер, Роланд; Раойлисон, Мамисоло; Бюттикер, Вильгельм; Лиз, Дэвид С.; Кренн, Харальд В. (22 апреля 2007 г.). «Малагасийские птицы как хозяева мотыльков, часто посещающих глаза» . Письма по биологии . 3 (2): 117–120. дои : 10.1098/rsbl.2006.0581 . ПМК 2375961 . ПМИД 17251126 .
- ^ Jump up to: а б с Scoble (1995) Глава 3: «Грудная клетка взрослого человека – исследование функций и эффектов» (стр. 39–40).
- ^ Скобл, MJ; Айелло, Аннет (1990). «Бабочки, похожие на мотыльков (Hedylidae: Lepidoptera): краткое описание с комментариями к яйцу» (PDF) . Журнал естественной истории . 24 (1): 159–164. Бибкод : 1990JNatH..24..159S . дои : 10.1080/00222939000770101 . [ постоянная мертвая ссылка ]
- ^ Jump up to: а б Чепмен, РФ (1998). «Торакс» . Насекомые: структура и функции (4-е изд.). Издательство Кембриджского университета . п. 45 . ISBN 978-0-521-57890-5 . .
- ^ Кришна, Анирудх; Не, Сяо; Уоррен, Эндрю Д.; Льоренте-Бускетс, Хорхе Э.; Бриско, Адриана Д.; Ли, Джэхо (2020). «Инфракрасно-оптические и тепловые свойства микроструктур крыльев бабочек» . Труды Национальной академии наук Соединенных Штатов Америки . 117 (3): 1566–1572. Бибкод : 2020PNAS..117.1566K . дои : 10.1073/pnas.1906356117 . ISSN 0027-8424 . ПМК 6983360 . ПМИД 31919285 .
- ^ Роббинс, Роберт К. (1981). «Гипотеза «ложной головы»: хищничество и изменение рисунка крыльев бабочек-ликанид». Американский натуралист . 118 (5): 770–775. дои : 10.1086/283868 . S2CID 34146954 .
- ^ Jump up to: а б Скобл (1995). Раздел «Крылья». Стр. 55.
- ^ Jump up to: а б Дадли, Роберт (2002). Биомеханика полета насекомых: форма, функция, эволюция . Издательство Принстонского университета . ISBN 978-0-691-09491-5 .
- ^ Jump up to: а б Стокс, Ян (2008). «Соединение крыльев» . В Капинере, Джон Л. (ред.). Энциклопедия энтомологии . Виртуальная справочная библиотека Гейла. Том. 4 (2-е изд.). Справочник Спрингера. п. 4266. ИСБН 978-1-4020-6242-1 .
- ^ Скобл (1995). Раздел «Сцепка крыла», (стр. 56–60).
- ^ Горб, Станислав (2001). «Соединение частей тела» . Приспособления для прикрепления кутикулы насекомых . Спрингер . п. 305. ИСБН 978-0-7923-7153-3 .
- ^ Харпер, Дуглас. «Чешуекрылые» . Интернет-словарь этимологии . Проверено 21 ноября 2010 г. из « Lepidoptera » на сайте Dictionary.com.
- ^ Дауни, Джей Си; Аллин, AC (1975). «Морфология и номенклатура чешуи крыльев». Бык. Аллин Муз . 31 : 1–32.
- ^ Чепмен (1988). Раздел «Крылья и полет» (с. 190).
- ^ Jump up to: а б с Гуллан, П.Дж.; Крэнстон, PS (2005). Насекомые: Очерк энтомологии (3-е изд.). Уайли-Блэквелл . ISBN 978-1-4051-1113-3 .
- ^ Мейсон, CW (1927). «Структурные цвета насекомых – II». Журнал физической химии . 31 (3): 321–354. дои : 10.1021/j150273a001 .
- ^ Вукусич, П. (2006). «Структурная окраска чешуекрылых» (PDF) . Современная биология . 16 (16): Р621–Р623. Бибкод : 2006CBio...16.R621V . дои : 10.1016/j.cub.2006.07.040 . ПМИД 16920604 . S2CID 52828850 .
- ^ Прум, РОД; Куинн, Т.; Торрес, Р.Х. (2006). «Все анатомически разнообразные чешуйки бабочек создают структурные цвета за счет когерентного рассеяния» . Журнал экспериментальной биологии . 209 (4): 748–765. дои : 10.1242/jeb.02051 . HDL : 1808/1600 . ПМИД 16449568 .
- ^ Киносита, Шуичи (2008). Структурные цвета в мире природы . Всемирная научная. стр. 52–53 . ISBN 978-981-270-783-3 .
- ^ Михильсен, К.; Ставенга, генеральный директор (2008). «Гироидные кутикулярные структуры в чешуйках крыльев бабочки: биологические фотонные кристаллы» . Журнал интерфейса Королевского общества . 5 (18): 85–94. дои : 10.1098/rsif.2007.1065 . ПМК 2709202 . ПМИД 17567555 .
- ^ Поладиан, Леон; Уикхэм, Шелли; Ли, Кван; Большой, Мэриэнн СиДжей (2009). «Иризация фотонных кристаллов и ее подавление в чешуе бабочки» . Журнал интерфейса Королевского общества . 6 (Приложение 2): С233–С242. doi : 10.1098/rsif.2008.0353.focus . ПМК 2706480 . ПМИД 18980932 .
- ^ Аргирос, А.; Манос, С.; Большой, MCJ; Маккензи, доктор медицинских наук; Кокс, GC; Дварте, DM (2002). «Электронная томография и компьютерная визуализация трехмерного «фотонного» кристалла в масштабе крыла бабочки». Микрон . 33 (5): 483–487. дои : 10.1016/S0968-4328(01)00044-0 . ПМИД 11976036 .
- ^ Гираделла, Хелен (1991). «Свет и цвет на крыле: структурные цвета бабочек и мотыльков». Прикладная оптика . 30 (24): 3492–3500. Бибкод : 1991ApOpt..30.3492G . дои : 10.1364/AO.30.003492 . ПМИД 20706416 .
- ^ Винтер-Блит, Массачусетс (1957). Бабочки Индийского региона (перепечатка 2009 г. издательством Today & Tomorrows Publishers, изд. Нью-Дели). Мумбаи, Индия: Бомбейское общество естествознания. ISBN 978-81-7019-232-9 .
- ^ «Андроконий» . Британская энциклопедия . Проверено 30 октября 2010 г.
- ^ Холл, Джейсон П.В.; Харви, Дональд Дж. (2002). «Обзор андрокониальных органов Riodinidae (Lepidoptera)» (PDF) . Зоологический журнал Линнеевского общества . 136 (2): 171–197. дои : 10.1046/j.1096-3642.2002.00003.x .
- ^ Комсток, Джон Генри (2008) [1920]. Введение в энтомологию . Читайте книги, первоначально опубликованные издательской компанией Comstock. ISBN 978-1-4097-2903-7 .
- ^ Скотт, Джеймс А. (1997). Бабочки Северной Америки: естественная история и путеводитель . Стэнфорд, Калифорния: Издательство Стэнфордского университета. ISBN 978-0804720137 . OCLC 49698782 .
- ^ Скобл (2005). Глава «Высшая дитризия», стр. 328.
- ^ Jump up to: а б с д и «Чешуекрылые» . Британская энциклопедия . Проверено 16 ноября 2010 г.
- ^ Jump up to: а б с д Скобл (1995). Раздел «Живот взрослого человека», (с. 98–102).
- ^ Уотсон, Трейси (3 июля 2013 г.). «Бражники уничтожают летучих мышей звуковыми взрывами из гениталий» . Природа . дои : 10.1038/nature.2013.13333 . S2CID 180859622 . Проверено 5 июля 2013 г.
- ^ Дагдейл, Дж. С. (1974). «Конфигурация женских половых органов в классификации чешуекрылых» . Новозеландский журнал зоологии . 1 (2). Королевское общество Новой Зеландии: 132. doi : 10.1080/03014223.1974.9517821 . Проверено 3 мая 2020 г.
- ^ Jump up to: а б с д и Скобл (1995). Глава «Незрелые стадии», (с. 104–133).
- ^ Jump up to: а б Нация, Джеймс Л. (2002). Физиология и биохимия насекомых . ЦРК Пресс . ISBN 978-0-8493-1181-9 .
- ^ Jump up to: а б с д и Чепмен (1998). Раздел «Яйцо и эмбриология» (с. 325–362).
- ^ Jump up to: а б с Голландия, WJ (1898). «Введение» ( PDF ) . Книга «Бабочка» . Лондон: ISBN Хатчинсона и Ко. 978-0-665-13041-0 .
- ^ Пи Джей Галлан; PS Крэнстон (2010). «Модели и фазы жизненной истории». Насекомые: Очерк энтомологии (4-е изд.). Уайли-Блэквелл . стр. 156–164 . ISBN 978-1-4443-3036-6 .
- ^ Jump up to: а б с д и ж Вагнер, Дэвид Л. (2005). Гусеницы восточной части Северной Америки . Издательство Принстонского университета . ISBN 978-0-691-12144-4 .
- ^ Jump up to: а б Миллер, Джеффри К. (3 августа 2006 г.). «Морфология гусеницы» . Гусеницы лесов и редколесий северо-запада Тихого океана . Центр исследования дикой природы северных прерий . Проверено 16 ноября 2010 г.
- ^ Макослейн, Хизер Дж. (2008). «Апосематизм» . В Капинере, Джон Л. (ред.). Энциклопедия энтомологии . Виртуальная справочная библиотека Гейла. Том. 4 (2-е изд.). Справочник Спрингера. ISBN 978-1-4020-6242-1 .
- ^ Jump up to: а б с д Общее, IFB (1990). Мотыльки Австралии . Издательство «Брилл» . ISBN 978-90-04-09227-3 .
- ^ Харпер, Дуглас. «Хризалис» . Интернет-словарь этимологии . Словарь.com . Проверено 16 ноября 2010 г.
- ^ Штер, Фредерик В. (2009). «Куколка и пупарий» . В Реше, Винсент Х.; Карде, Ринг Т. (ред.). Энциклопедия насекомых (2-е изд.). Академическая пресса . стр. 970–973. ISBN 978-0-12-374144-8 .
- ^ Фигье, Луи (1868). Мир насекомых: популярный отчет об отрядах насекомых вместе с описанием привычек и экономики некоторых из наиболее интересных видов . Нью-Йорк: Д. Эпплтон и компания.
- ^ Сураков, Андрей (2008). «Спаривание куколки зебры длиннокрылой ( Heliconius charithonia ): фотографические свидетельства». Новости Общества лепидоптерологов . 50 (1): 26–32.
- ^ Jump up to: а б с «Защитные механизмы гусеницы и бабочки» . EnchantedLearning.com . Проверено 7 декабря 2009 г.
- ^ Латимер, Джонатан П.; Карен Стрей Нолтинг (2000). Бабочки . Хоутон Миффлин Харкорт . п. 12 . ISBN 978-0-395-97944-0 .
Тигровый махаон.
- ^ Кричер, Джон (1999). «6» . Неотропический спутник . Издательство Принстонского университета . стр. 157–158. ISBN 978-0-691-00974-2 .
- ^ Сантос, Джей Си; Каннателла, округ Колумбия (2003). «Множественные, повторяющиеся причины апосематизма и специализации в питании ядовитых лягушек» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (22): 12792–12797. Бибкод : 2003PNAS..10012792S . дои : 10.1073/pnas.2133521100 . ПМК 240697 . ПМИД 14555763 .
- ^ Насекомые и пауки мира . Том. 10. Корпорация Маршалл Кавендиш. Маршалл Кавендиш. Январь 2003 г., стр. 292–293. ISBN 978-0-7614-7344-2 .
{{cite book}}: CS1 maint: другие ( ссылка ) - ^ Кэрролл, Шон (2005). Бесконечные самые красивые формы: новая наука Эво Дево и создание царства животных . WW Norton & Co., стр. 205–210 . ISBN 978-0-393-06016-4 .
Защита глазных пятен бабочки.
- ^ Хеффернан, Эмили (2004). Симбиотические отношения между Anthene emolus (Lycaenidae) и Oecophylla smaragdina (Formicidae): обязательный мутуализм в тропических лесах Малайзии (PDF) ( магистерская диссертация). Университет Флориды .
- ^ «Осмеритий» . Мерриам-Вебстер . Проверено 9 декабря 2009 г.
- ^ Хэдли, Дебби. «Осмеритий» . Руководство About.com. Архивировано из оригинала 23 июля 2008 года . Проверено 9 декабря 2009 г.
Внешние ссылки
[ редактировать ]- СЭМ-изображение чешуи бабочки и ее ножки (третье сверху) .
- Изысканные потерпевшие кораблекрушение – фоторепортаж о яйцах чешуекрылых от National Geographic .
- Необычное видение – фоторепортаж о мотыльках от National Geographic .