Чешуйчатоногий брюхоногий моллюск
| Чешуйчатоногий / Морской панголин | |
|---|---|

| |
| Chrysomallon sqamiferum из Лунци. Масштабная линейка составляет 1 см. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Моллюска |
| Сорт: | Брюхоногие моллюски |
| Подкласс: | Неомфалионы |
| Заказ: | Неомфалида |
| Семья: | пельтоспириды |
| Род: | Хризомаллон Чен, Линс , Копли и Роджерс, 2015 г. |
| Разновидность: | C. sqamiferum
|
| Биномиальное имя | |
| Хризомаллон чешуйчатый Чен, Линс, Копли и Роджерс, 2015 г. [ 2 ]
| |
| Синонимы [ 2 ] | |
|
Crysomallon sqamiferum (орт. ошибка) | |

Chrysomallon sqamiferum , широко известный как чешуйчатый брюхоногий моллюск , чешуйчатая улитка , морской панголин или улитка-вулкан . [ 3 ] [ 4 ] — вид глубоководной гидротермальной улитки , морского брюхоногого моллюска семейства Peltospiridae . [ 2 ] Этот эндемичный жерлобрюхоногий моллюск известен только из глубоководных гидротермальных источников в Индийском океане , где он был обнаружен на глубинах около 2400–2900 м (1,5–1,8 мили). C. sqamiferum сильно отличается от других глубоководных брюхоногих моллюсков, даже от близкородственных неомфалин. [ 5 ] В 2019 году он был объявлен находящимся под угрозой исчезновения и занесен в Красный список МСОП. [ 6 ] первый вид, внесенный в список таковых из-за рисков, связанных с глубоководной разработкой его жерловой среды обитания. [ 7 ]
Оболочка имеет уникальную трехслойную конструкцию; внешний слой состоит из сульфидов железа , средний слой эквивалентен органическому периостракуму, обнаруженному у других брюхоногих моллюсков, а самый внутренний слой состоит из арагонита . Стопа также необычна: по бокам она покрыта минерализованными железом склеритами .
улитки В пищеводной железе обитают симбиотические гаммапротеобактерии, от которых улитка, по-видимому, получает питание. Этот вид считается одним из наиболее своеобразных глубоководных брюхоногих моллюсков гидротермальных источников и единственным известным из ныне живущих животных, у которого в скелете (как в склеритах, так и в панцире в качестве экзоскелета) присутствует сульфид железа. [ 2 ] Его сердце, пропорционально говоря, необычно велико для любого животного: сердце составляет примерно 4% объема его тела. [ 5 ]
Таксономия
[ редактировать ]Этот вид был впервые обнаружен в апреле 2001 года, а с 2001 года его называют «чешуйчатоногим» брюхоногим моллюском. [ 8 ] С 2003 года его называют Chrysomallon sqamiferum , но он не был официально описан в смысле Международного кодекса зоологической номенклатуры до тех пор, пока Chen et al. назвал его в 2015 году. [ 2 ] [ 9 ] Типовые экземпляры хранятся в Музее естественной истории в Лондоне . [ 2 ] Во времена, когда название еще не было официально оформлено, неправильным вариантом написания было « Crysomallon sqamiferum ». [ 2 ]
Chrysomallon sqamiferum — типовой вид и единственный вид рода Chrysomallon . [ 2 ] Родовое название Chrysomallon происходит от древнегреческого языка и означает «золотоволосый», поскольку пирит (соединение, встречающееся в его оболочке) имеет золотистый цвет. [ 2 ] Видовое название sqamiferum происходит из латинского языка и означает «чешуйчатый» из-за его склеритов. [ 2 ] Сначала было неизвестно, к какому семейству принадлежит этот вид. [ 8 ] Варен и др. в 2003 году отнес этот вид к семейству Peltospiridae, входящему в состав Neomphalina . [ 10 ] Молекулярный анализ, основанный на последовательностях генов цитохром-с-оксидазы I (COI), подтвердил отнесение этого вида к Peltospiridae . [ 2 ] [ 11 ] Морфотипы из двух местонахождений темные; морфотип из третьего местонахождения белый (пояснение местонахождений см. в следующем разделе). [ 2 ] [ 12 ] [ 13 ] Согласно результатам генетического анализа, эти разноцветные улитки кажутся просто «разновидностями» одного и того же вида. [ 2 ]
Распределение
[ редактировать ]
Чешуйчатоногий брюхоногий моллюск — это эндемичный брюхоногий моллюск, известный только из глубоководных гидротермальных источников Индийского океана, глубина которых составляет около 2780 метров (1,73 мили). [ 2 ] Вид был открыт в 2001 году, обитая на базах черных курильщиков в поле Кайрей гидротермальном , 25 ° 19,239' ю.ш., 70 ° 02,429' в.д. / 25,320650 ° ю.ш., 70,040483 ° в.д. , на Центрально-Индийском хребте, к северу от Тройной точки Родригес . [ 8 ] Впоследствии этот вид также был обнаружен на поле Пасьянс. 19 ° 33,413' ю.ш., 65 ° 50,888' в.д. / 19,556883 ° ю.ш., 65,848133 ° в.д. , Центрально-Индийский хребет , в исключительной экономической зоне Маврикия . [ 14 ] [ 15 ] и Лунци (в переводе с китайского означает «Флаг Дракона») [ 16 ] поле, 37 ° 47,027' ю.ш., 49 ° 38,963' в.д. / 37,783783 ° ю.ш., 49,649383 ° в.д. , Юго-Западный Индийский хребет . [ 17 ] [ 18 ] Типовым месторождением было обозначено месторождение Лунци; все типы материалов произошли из этого жерлового поля. [ 2 ] Расстояние между Кайрей и Солитер составляет около 700 км (430 миль). Расстояние между Пасьянсом и Лунци составляет около 2500 км (1600 миль). [ 2 ] Эти три участка принадлежат биогеографической провинции гидротермальных жерловых систем Индийского океана в смысле Роджерса и др. (2012). [ 19 ] Расстояние между участками велико, но общая площадь распространения очень мала, менее 0,02 квадратных километров (0,0077 квадратных миль). [ 20 ]

Известно, что улитки Peltospiridae в основном обитают в жерловых полях восточной части Тихого океана. Накамура и др. выдвинул гипотезу, что появление чешуйчатоногого брюхоногого моллюска в Индийском океане предполагает родство фаун гидротермальных жерл между этими двумя районами. [ 14 ]

Исследовательские экспедиции включали:
- 2000 г. - экспедиция Японского агентства по морским наукам и технологиям о Земле с использованием корабля RV Kairei и ROV Kaikō обнаружила жерловое поле Кайрей, но чешуйчатоногих брюхоногих моллюсков тогда обнаружено не было. [ 21 ] Это было первое жерловое поле, обнаруженное в Индийском океане. [ 21 ]
- 2001 г. – экспедиция американского научно-исследовательского судна RV Knorr с ROV Jason обнаружила чешуйчатоногих брюхоногих моллюсков в жерловом поле Кайрей. [ 8 ]
- 2007 г. – экспедиция теплохода Да Ян И Хао открыла жерловое поле Лунци. [ 2 ]
- 2009 г. - экспедиция НИС «Йокосука» с DSV Shinkai 6500 обнаружила месторождение «Солитер» и отобрала там образцы чешуйчатоногих брюхоногих моллюсков. [ 14 ]
- 2009 г. - экспедиция теплохода Да Ян И Хао визуально наблюдала чешуйчатоногих брюхоногих моллюсков на жерлом Лунци. [ 2 ] [ 17 ]
- 2011 г. - экспедиция британского королевского исследовательского корабля RRS James Cook с ROV Kiel 6000 взяла пробы жерлового поля Лунци. [ 2 ] [ 22 ]
Описание
[ редактировать ]Склериты
[ редактировать ]
У этого вида стороны стопы улитки чрезвычайно необычны и покрыты сотнями минерализованных железом склеритов ; они состоят из сульфидов железа [ 10 ] грейгит и пирит . [ 23 ] Каждый склерит имеет ядро из мягкой эпителиальной ткани, конхиолиновую оболочку и самый верхний слой, содержащий пирит и грейгит. [ 2 ] До открытия чешуйчатоногих брюхоногих моллюсков считалось, что единственные современные моллюски, обладающие чешуйчатыми структурами, относятся к классам Caudofoveata , Solenogastres и Polyplacophora . [ 18 ] Склериты не гомологичны покрышке брюхоногих моллюсков . Склериты чешуйчатоногих брюхоногих моллюсков также не гомологичны склеритам хитонов (Polyplacophora). [ 18 ] Было высказано предположение, что склериты кембрийских халваксиид , таких как Halkieria, потенциально могут быть более аналогичны склеритам этой улитки, чем склериты хитонов или аплакофоран. [ 18 ] Еще в 2015 году подробный морфологический анализ для проверки этой гипотезы не проводился. [ 18 ]
Склериты C. sqamiferum преимущественно белковые (конхиолин — сложный белок); напротив, склериты хитонов преимущественно известковые. [ 18 ] На поперечных срезах склеритов видимых линий роста конхиолина нет. [ 18 ] Никакие другие современные или вымершие брюхоногие моллюски не имеют кожных склеритов. [ 18 ] и ни одно другое современное животное не использует сульфиды железа таким образом ни в своем скелете , ни в скелете . [ 2 ] или экзоскелет .
Размер каждого склерита у взрослых составляет около 1 × 5 мм. [ 2 ] У молодых особей чешуя расположена в несколько рядов, а у взрослых чешуя плотная и асимметричная. [ 24 ] Популяция улиток Solitaire имеет белые склериты вместо черных; это связано с недостатком железа в склеритах. [ 18 ] Склериты черепитчатые (накладываются друг на друга, напоминающие черепицу ). [ 5 ] Предполагается, что целью склеритов является защита или детоксикация . [ 25 ] Склериты могут помочь защитить брюхоногих моллюсков от вентиляционной жидкости, чтобы их бактерии могли жить рядом с источником доноров электронов для хемосинтеза. [ 5 ] Или, альтернативно, склериты могут возникнуть в результате отложения токсичных сульфидных отходов эндосимбионтов и, следовательно, представляют собой новое решение для детоксикации. [ 5 ] Но истинная функция склеритов пока неизвестна. [ 14 ] Склериты Кайрейской популяции, имеющие слой сульфида железа, ферримагнитны . [ 2 ] Нежелезосульфидно-минерализованный склерит морфотипа Solitaire показал большую механическую прочность всей конструкции при трехточечном изгибе (12,06 МПа), чем склерит морфотипа Kairei (6,54 МПа). [ 14 ]
При жизни на внешней поверхности склеритов обитает разнообразный набор эпибионтов : Campylobacterota (ранее Epsilonproteobacteria) и Thermodesulfobacteriota (ранее часть Deltaproteobacteria). [ 26 ] Эти бактерии, вероятно, обеспечивают их минерализацию. [ 26 ] Гоффреди и др. (2004) предположили, что улитка выделяет некоторые органические соединения, которые облегчают прикрепление бактерий. [ 26 ]
Оболочка
[ редактировать ]
Кайрей, Лунци, Пасьянс (слева направо)
Раковина этих видов имеет три оборота . [ 2 ] Форма корпуса шаровидная, шпиль сжатый. [ 2 ] Скульптура ракушки состоит из ребер и тонких линий роста. [ 2 ] Форма отверстия эллиптическая . [ 2 ] Вершина панциря хрупкая, у взрослых особей подвергается коррозии. [ 2 ]
Это очень крупный пельтоспирид по сравнению с большинством других видов, которые обычно ниже 15 миллиметров ( 3 ⁄ дюйма ). Длина корпуса [ 2 ] Ширина раковины 9,80–40,02 мм (0,39–1,58 дюйма); [ 2 ] максимальная ширина корпуса достигает 45,5 миллиметров (1,79 дюйма). [ 2 ] Средняя ширина раковины взрослых улиток составляет 32 мм. [ 2 ] Средняя ширина раковины в популяции Solitaire была несколько меньше, чем в популяции Kairei. [ 15 ] Высота раковины 7,65–30,87 мм (0,30–1,22 дюйма). [ 2 ] Ширина апертуры составляет 7,26–32,52 мм (0,29–1,28 дюйма). [ 2 ] Высота апертуры составляет 6,38–27,29 мм (0,25–1,07 дюйма). [ 2 ]
Структура оболочки состоит из трех слоев. Внешний слой толщиной около 30 мкм, черного цвета, выполнен из сульфидов железа, содержит грейгит Fe 3 S 4 . [ 27 ] Этот вид — единственное известное из ныне живущих животных, в скелете которого имеется этот материал. [ 2 ] Средний слой (около 150 мкм) эквивалентен органическому периостракуму, который встречается и у других брюхоногих моллюсков. [ 27 ] Периостракум толстый, коричневый. [ 2 ] Самый внутренний слой состоит из арагонита (толщиной около 250 мкм) — формы карбоната кальция , которая обычно встречается как в раковинах моллюсков, так и в различных кораллах. [ 27 ] Цвет слоя арагонита молочно-белый. [ 2 ]
Каждый слой раковины по-разному способствует эффективности защиты улитки. Средний органический слой, по-видимому, поглощает механическое напряжение и энергию, возникающую при сдавливании (например, клешнями краба), что делает панцирь намного более прочным. Органический слой также рассеивает тепло. [ 28 ] Характеристики этого композиционного материала находятся в центре внимания исследователей для возможного использования в гражданских и военных защитных целях. [ 27 ]
-
 Две разновидности чешуйчатоногих брюхоногих моллюсков
Две разновидности чешуйчатоногих брюхоногих моллюсков -
 C. sqamiferum из жерлового поля Кайрей
C. sqamiferum из жерлового поля Кайрей -
 C. sqamiferum из жерлового поля Солитер
C. sqamiferum из жерлового поля Солитер



Оперкулум
[ редактировать ]У этого вида форма жаберной крышки меняется в процессе роста: от округлой у молоди до изогнутой у взрослых. [ 14 ] Относительный размер жаберной крышки уменьшается по мере роста особи. [ 5 ] Около половины всех взрослых улиток этого вида имеют жаберную крышку среди склеритов в задней части животного. [ 14 ] Кажется вероятным, что склериты постепенно разрастаются и полностью покрывают всю стопу, обеспечивая защиту, а жаберная крышка теряет свою защитную функцию по мере роста животного. [ 14 ]
-
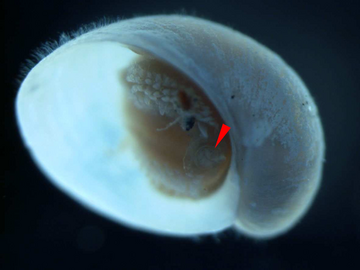 Молодь с жаберной крышкой обозначена красным указателем. Длина раковины около 2 мм.
Молодь с жаберной крышкой обозначена красным указателем. Длина раковины около 2 мм. -
 Крышка молодой улитки. Масштабная линейка составляет 1 мм.
Крышка молодой улитки. Масштабная линейка составляет 1 мм. -
 Крышка взрослой улитки. Масштабная линейка составляет 1 мм.
Крышка взрослой улитки. Масштабная линейка составляет 1 мм. -
 Взрослые улитки с крышечкой, обозначенной красными стрелками. Масштабная линейка составляет 5 мм.
Взрослые улитки с крышечкой, обозначенной красными стрелками. Масштабная линейка составляет 5 мм.




Внешняя анатомия
[ редактировать ]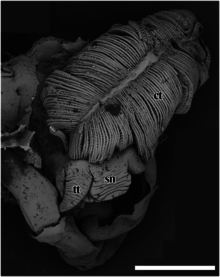
sn – морда,
тт – головное щупальце.
Масштабная линейка составляет 2 мм.
Чешуйчатоногий брюхоногий моллюск имеет толстую морду, которая сужается дистально к тупому концу. Рот представляет собой круглое кольцо мышц в сжатом и закрытом состоянии. [ 5 ] Два гладких головных щупальца толстые у основания и постепенно сужаются к тонким кончикам на дистальных концах. [ 5 ] У этой улитки нет глаз. [ 5 ] Специализированного копулятивного придатка нет. [ 5 ] Ножка красная и большая, и улитка не может полностью убрать ее в раковину. [ 2 ] отсутствует . Педальная железа в передней части стопы [ 5 ] Эпиподиальных щупалец также нет. [ 5 ]
Внутренняя анатомия
[ редактировать ]У C. sqamiferum мягкие части животного занимают примерно два оборота внутренней части раковины. [ 5 ] Панцирная мышца подковообразной формы, крупная, разделена на две части слева и справа и соединена более узким прикреплением. [ 5 ] Край мантии толстый, простой, без каких-либо отличительных черт. [ 5 ] Мантийная полость глубокая, доходит до заднего края раковины. [ 5 ] В медиальной левой части полости преобладает очень большой двугребешковый ктенидий . [ 5 ] Вентральнее висцеральной массы полость тела занята огромной пищеводной железой, которая простирается и заполняет вентральное дно мантийной полости. [ 5 ] [ 26 ]
проста Пищеварительная система , ее объем уменьшен до менее чем 10% от объема, типичного для брюхоногих моллюсков. [ 5 ] [ 26 ] Радула . «слабая», типа рифидоглоссана, с одной парой радулярных хрящей [ 5 ] [ 26 ] Формула радулы : ~50 + 4 + 1 + 4 + ~50. [ 2 ] Лента радулы длиной 4 мм, шириной 0,5 мм; [ 2 ] соотношение ширины к длине составляет примерно 1:10. [ 5 ] . Челюсти и слюнных желез нет [ 5 ] Часть переднего отдела пищевода быстро разрастается в огромную гипертрофированную пищеводную железу со слепыми концами, занимающую большую часть вентральной поверхности мантийной полости (около 9,3% объема тела). [ 5 ] Пищеводная железа растет изометрически вместе с улиткой, что соответствует улитке, зависящей от ее эндосимбионтных микробов на протяжении всей ее оседлой жизни. [ 24 ] Пищеводная железа имеет однородную структуру, хорошо васкуляризирована и имеет мелкие кровеносные сосуды. [ 5 ] Желудок пищеварительной имеет как минимум три протока в передней правой части, соединяющихся с железой . [ 5 ] Консолидированные гранулы имеются как в желудке, так и в задней кишке. [ 5 ] Эти гранулы, вероятно, представляют собой гранулы серы, производимые эндосимбионтом для детоксикации сероводорода. [ 5 ] Кишка редуцирована и имеет только одну петлю. [ 5 ] Обширная и рыхлая пищеварительная железа простирается назад, заполняя вершину раковины . [ 5 ] Прямая кишка не проникает в сердце, а проходит вентрально от него. [ 5 ] Анус . расположен на правой стороне улитки, над половым отверстием [ 5 ]
В выделительной системе нефридий . занимает центральное место, тяготеет к правой стороне тела, представляет собой тонкий темный слой железистой ткани [ 5 ] Нефридий находится в передней и вентральной части пищеварительной железы и соприкасается с дорсальной стороной передней кишки. [ 5 ]
Дыхательная система и система кровообращения состоят из единственного левого двугребешкового ктенидия (жаберного), очень большого размера (15,5% объема тела), поддерживаемого множеством крупных и подвижных кровеносных синусов, заполненных гемоцелем . [ 5 ] [ 24 ] При вскрытии заметными особенностями всей полости тела являются кровеносные синусы и комки материала гемоцеля. [ 5 ] Хотя система кровообращения у Chrysomallon в основном закрыта (это означает, что гемоцель в основном не покидает кровеносные синусы), выступающие кровеносные синусы кажутся временными и возникают в разных областях тела у разных людей. [ 24 ] По обе стороны от ктенидия имеются тонкие жаберные нити. [ 5 ] Двояковыпуклый ктенидий простирается далеко позади сердца до верхних оборотов раковины; он значительно крупнее, чем у Peltospira . Хотя этот вид имеет форму раковины и общую форму, сходную с другими пельтоспиридами, ктенидий пропорционален размеру Hirtopelta , у которого самые большие жабры среди родов пельтоспирид, которые были исследованы анатомически до сих пор. [ 5 ]
Ктенидий обеспечивает улитку кислородом, но ее кровеносная система расширена и выходит за рамки других подобных жерловых брюхоногих моллюсков. [ 5 ] Эндосимбионтов в жабрах C. sqamiferum и на них нет . [ 5 ] Увеличение жабр, вероятно, призвано облегчить извлечение кислорода в условиях низкого содержания кислорода, типичных для гидротермальных жерловых экосистем. [ 5 ]
В задней части ктенидия находится удивительно большое и хорошо развитое сердце. [ 5 ] Сердце необычайно велико для любого пропорционального животного. [ 5 ] В зависимости от объема отдельного предсердия и желудочка сердечный комплекс составляет примерно 4% объема тела (например, сердце человека составляет 1,3% объема тела). [ 5 ] Длина желудочка у молодых особей составляет 0,64 мм, длина оболочки 2,2 мм, у взрослых длина вырастает до 8 мм. [ 24 ] Это пропорционально гигантское сердце в основном всасывает кровь через ктенидий и снабжает хорошо васкуляризированную пищеводную железу. [ 5 ] У C. sqamiferum эндосимбионты обитают в пищеводной железе, где они изолируются от вентиляционной жидкости. [ 5 ] Таким образом, хозяин, вероятно, будет играть важную роль в снабжении эндосимбионтов необходимыми химическими веществами, что приводит к увеличению респираторных потребностей. [ 5 ] Детальное исследование гемоцеля C. sqamiferum позволит получить дополнительную информацию о его дыхательных пигментах. [ 5 ]

ct – ктенидий,
пм – педальная мышца,
сбн – весы,
си – кровяной синус,
ты – яичко.
Масштабная линейка равна 1 см.
Чешуйчатоногий брюхоногий моллюск — хемосимбиотический холобионт . [ 26 ] Он содержит тиоавтотрофные (окисляющие серу) гаммапротеобактериальные эндосимбионты в значительно увеличенной пищеводной железе и, по-видимому, использует эти симбионты для питания. [ 26 ] [ 29 ] Ближайшим известным родственником этого эндосимбионта является улитка Alviniconcha . [ 30 ] У этого вида размеры пищеводной железы примерно на два порядка превышают обычные размеры. [ 26 ] Внутри пищеводной железы имеется значительное разветвление, где артериальное давление, вероятно, снижается почти до нуля. [ 5 ] Сложная сердечно-сосудистая система, скорее всего, развилась для насыщения эндосимбионтов кислородом в среде с низким содержанием кислорода и/или для снабжения сероводородом . эндосимбионтов [ 5 ] Тиоавтотрофные гаммапротеобактерии обладают полным набором генов, необходимых для аэробного дыхания , и, вероятно, способны переключаться между более эффективным аэробным дыханием и менее эффективным анаэробным дыханием в зависимости от доступности кислорода. [ 5 ] В 2014 году эндосимбионт чешуйчатоногого брюхоногого моллюска стал первым эндосимбионтом любого брюхоногого моллюска, полный геном которого был известен. [ 29 ] C. sqamiferum является единственным видом Peltospiridae, имеющим увеличенную пищеводную железу. Ранее считалось, что [ 2 ] но позже было обнаружено, что оба вида Gigantopelta имеют также увеличенную пищеводную железу. [ 11 ] Chrysomallon и Gigantopelta — единственные жерловые животные, за исключением сибоглинидных трубчатых червей, у которых эндосимбионты обитают в замкнутой части тела, не находящейся в прямом контакте с жерловой жидкостью. [ 24 ]
Нервная система крупная, а мозг представляет собой сплошную нервную массу без ганглиев. [ 5 ] Нервная система усложнена и увеличена в размерах по сравнению с другими таксонами неомфалинов. [ 5 ] Как это типично для брюхоногих моллюсков, нервная система состоит из переднего пищеводного нервного кольца и двух пар продольных нервных тяжей: вентральной пары, иннервирующей стопу, и дорсальной пары, образующей скручивание посредством стрептоневрии . [ 5 ] Фронтальная часть пищеводного нервного кольца крупная, соединяет два латеральных вздутия. [ 5 ] Огромная сросшаяся нервная масса непосредственно прилегает к пищеводной железе, где обитают бактерии, и проходит через нее. [ 5 ] В головные щупальца входят большие щупальцевые нервы. [ 5 ] Органы чувств чешуйчатоногих брюхоногих включают статоцисты, окруженные пищеводной железой, причем каждый статоцист содержит один статолит . [ 5 ] На кончиках жаберных лепестков имеются также чувствующие ктенидальные бурсикулы; Известно, что они присутствуют у большинства ветигастропод, а также у некоторых неомфалинов. [ 5 ]
Репродуктивная система имеет некоторые необычные особенности. Гонады взрослых улиток не находятся внутри раковины; они находятся в области головы и стоп на правой стороне тела. [ 5 ] У молоди с длиной раковины 2,2 мм гонады отсутствуют. [ 24 ] находятся У взрослых и семенники , и яичники на разных уровнях развития. [ 5 ] Семенник расположен вентрально; яичник располагается дорсально, а нефридий лежит между ними. [ 5 ] Рядом с семенником находится «орган упаковки сперматофоров». [ 5 ] Гонодукты семенника и яичника первоначально разделены, но, по-видимому, сливаются в один проток и возникают в виде единого генитального отверстия справа от мантийной полости. [ 5 ] У животного нет копулятивного органа. [ 2 ] [ 5 ]
 |

|

|
Предполагается, что разработанная стратегия размещения эндосимбиотических микробов в пищеводной железе стала катализатором анатомических инноваций, которые служат в первую очередь улучшению приспособленности бактерий сверх потребностей улитки. [ 5 ] Сильное увеличение пищеводной железы, защитные кожные склериты улитки, ее сильно увеличенная дыхательная и кровеносная системы, а также высокая плодовитость — все это считается адаптациями, полезными для ее эндосимбионтных микробов. [ 5 ] Эти адаптации, по-видимому, являются результатом специализации по удовлетворению энергетических потребностей в экстремальной хемосинтетической среде. [ 5 ]
Экология
[ редактировать ]среда обитания
[ редактировать ]Этот вид населяет гидротермальные жерла Индийского океана. Он обитает рядом как с кислой, так и с восстановительной жидкостью, на стенках дымоходов черных курильщиков или непосредственно в местах диффузного потока. [ 5 ]
Глубина месторождения Кайрей варьируется от 2415 до 2460 м (от 7923 до 8071 футов). [ 8 ] его размеры составляют примерно 30 на 80 м (98 на 262 фута). [ 8 ] Уклон поля составляет от 10° до 30°. [ 8 ] Порода - субстрат представляет собой троктолит и истощенный базальт срединно-океанического хребта . [ 31 ] Чешуйчатоногие брюхоногие Кайрейские поля обитают в низкотемпературных диффузных жидкостях одного дымохода. [ 14 ] Переходная зона, где были обнаружены эти брюхоногие моллюски, имеет ширину около 1–2 м (3–7 футов) и температуру 2–10 °C. [ 32 ] Предпочтительная температура воды для этого вида составляет около 5 °C. [ 33 ] Эти улитки живут в среде с высокой концентрацией сероводорода и низкой концентрацией кислорода. [ 33 ]
Численность чешуйчатоногих брюхоногих на поле Кайрей была ниже, чем на поле Лунци. [ 2 ] Сообщество Кайрейского гидротермального источника состоит из 35 таксонов. [ 34 ] включая морские анемоны Marianactis sp., ракообразные Austinograea Rodriguezensis , Rimicaris kairei , Mirocaris indica , Munidopsis sp., род и виды Neolepadidae , Eochionelasmus sp., двустворчатые моллюски Bathymodiolus marisindicus , брюхоногие моллюски Lepetodrilus sp., Pseudorimula sp., Eulepetopsis sp., с сп ., и Alviniconcha marisindica , [ 35 ] Дебрюйерезия марисиндика , [ 36 ] Брюсиелла варени , [ 36 ] Phymorhynchus sp., Sutilizona sp., блюдечко sp. 1, щелевое блюдечко зр. 2, Ифинопсис бушети , [ 36 ] Соленогастрес Хеликорадомения ? sp., кольчатые черви Amphisamytha sp., Archinome jasoni , Capitellidae sp. 1, Ophyotrocha sp., Hesionidae sp. 1, Hesionoidae sp. 2, Branchinotogluma sp., Branchipolynoe sp., Harmothoe ? sp., Левенштейниелла ? sp., Prionospio sp., неопознанная Nemertea и неопознанная Platyhelminthes . [ 34 ] Чешуйчатоногие брюхоногие моллюски обитают колониями с улитками Alviniconcha marisindica , а над ними располагаются колонии Rimicaris kairei . [ 33 ]
Поле Пасьянс находится на глубине 2606 м (8550 футов), а его размеры составляют примерно 50 на 50 м (160 на 160 футов). [ 14 ] Порода-субстрат представляет собой обогащенный базальт срединно-океанического хребта. [ 14 ] [ 31 ] Чешуйчатоногие брюхоногие моллюски обитают вблизи высокотемпературных диффузных жидкостей дымовых труб жерлового поля. [ 14 ] Численность чешуйчатоногих брюхоногих на поле Солитер была ниже, чем на поле Лунци. [ 2 ] Сообщество гидротермальных источников Solitaire включает 22 таксона, в том числе: морские анемоны Marianactis sp., ракообразные Austinograea Rodriguezensis , Rimicaris kairei , Mirocaris indica , Munidopsis sp., Neolepadidae gen et sp., Eochionelasmus sp., двустворчатые моллюски Bathymodiolus marisindicus , брюхоногие моллюски Lepetodrilus sp. , Eulepetopsis sp., Shinkailepas sp., Alviniconcha sp. тип 3, Desbruyeresia sp., Phymorhynchus sp., кольчатые черви рода и вида Alvinellidae , Archinome jasoni , Branchinotogluma sp., иглокожие голотурии Apodacea gen et sp., рыбы рода Macrouridae и виды, неидентифицированные Nemertea и неопознанные Platyhelminthes . [ 34 ]

Поле жерл Лунци находится на глубине 2780 м (9120 футов). [ 2 ] его размеры составляют примерно 100 на 150 м (330 на 490 футов). [ 20 ] C. sqamiferum был густо заселен в районах, непосредственно окружающих диффузное жерло. [ 5 ] Сообщество гидротермальных источников Лунци включает 23 [ Примечание 1 ] Таксоны макро- и мегафауны: морские анемоны Actinostolidae sp., кольчатые черви Polynoidae n. ген. н. сп. «655», Бранчиполыное с. сп. «Дракон», Пейналеополиное с. сп. «Дракон», Гезиолира ср. bergi , Hesionidae sp. индет., Ophryotrocha n. сп. «F-038/1b», Прионоспио ср. одноламеллата , Ampharetidae sp. indet., мидии Bathymodiolus marisindicus , брюхоногие моллюски Gigantopelta aegis , [ 11 ] Дракогира подфускус , Лирапекс полированный , [ 16 ] Фиморинх н. сп. «SWIR» , Lepetodrilus n. сп. «SWIR», ракообразные Neolepas sp. 1, Rimicaris kairei , Mirocaris indica , Chorocaris sp., Kiwa n. сп. «SWIR»17, Munidopsis sp. и иглокожие голотурии Chiridota sp. [ 17 ] [ 37 ] Плотность Lepetodrilus n. сп. «SWIR» и чешуйчатоногих брюхоногих моллюсков – более 100 улиток на м2. 2 на близком расстоянии от источников жерловой жидкости на жерловом поле Лунци. [ 37 ]
Привычки питания
[ редактировать ]Чешуйчатоногий брюхоногий моллюск является облигатным симбиотрофом на протяжении всей постпоселенческой жизни. [ 24 ] На протяжении всей постличиночной жизни чешуйчатоногие брюхоногие получают все свое питание за счет хемоавтотрофии своих эндосимбиотических бактерий. [ 26 ] [ 24 ] Чешуйчатоногий брюхоногий моллюск не является фильтратором. [ 5 ] [ 24 ] и не использует другие механизмы для кормления. [ 5 ] Радула и хрящ радулы небольшие и составляют соответственно лишь 0,4% и 0,8% объема тела молоди по сравнению с 1,4% и 2,6% у миксотрофной молоди Gigantopelta Chessoia . [ 24 ]
Для идентификации трофических взаимодействий в среде обитания, где прямое наблюдение за пищевыми привычками затруднено, можно измерить состав стабильных изотопов углерода и азота. [ 32 ] Имеются обедненные значения δ 13 C в пищеводной железе (по отношению к органическому углероду, полученному фотосинтезом). [ 26 ] Источником такого углерода считались хемоавтотрофные симбионты. [ 26 ] Хемоавтотрофное происхождение стабильного изотопа углерода 13 С было подтверждено экспериментально. [ 29 ]
| Салфетка | д 13 С | д 15 Н |
|---|---|---|
| Пищеводная железа | −20.7 ± 0.9 ‰ | 3.3 ± 1.8 ‰ |
| Джилл | −18,3 ± 0,6 ‰, от −17,4 до −18,8 ‰ | 3,9 ± 0,6 ‰, от 3,1 до 4,2 ‰ |
| Мантия | от −17,5 до −18,6 ‰ | от 3,5 до 4,7 ‰ |
| Ступня | −18.2 ± 0.6 ‰ | 3.8 ± 0.5 ‰ |
| Весы | −16.7 ± 0.6 ‰ | 3.8 ± 0.9 ‰ |
Жизненный цикл
[ редактировать ]Этот брюхоногий моллюск одновременно является гермафродитом . [ 5 ] Это единственный вид семейства Peltospiridae, который, как известно, одновременно является гермафродитом. [ 5 ] Имеет высокую плодовитость . [ 5 ] Откладывает яйца, вероятно, лецитотрофного типа. [ 22 ] Яйца чешуйчатоногого брюхоногого моллюска обладают отрицательной плавучестью под атмосферным давлением. [ 15 ] По состоянию на 2016 год ни личинки, ни протоконх не известны, но считается, что этот вид находится на стадии планктонного расселения. [ 22 ] Самые маленькие из когда-либо собранных молодых особей C. sqamiferum имели длину раковины 2,2 мм. [ 24 ] Результаты статистического анализа не выявили генетической дифференциации между двумя популяциями в полях Кайрей и Солитер, что позволяет предположить потенциальную связь между двумя жерловыми полями. [ 15 ] Популяция Кайрей представляет собой потенциальную исходную популяцию для двух популяций Центрально-Индийского хребта. [ 15 ] Этих улиток трудно сохранить в искусственной среде; однако они выживали в аквариумах при атмосферном давлении более трех недель. [ 33 ]
Меры по сохранению и угрозы
[ редактировать ]Чешуйчатоногий брюхоногий моллюск не защищен. [ 1 ] [ 20 ] Его потенциальная среда обитания во всех гидротермальных жерлах Индийского океана оценивается не более чем в 0,27 квадратных километров (67 акров), в то время как три известных участка, в которых он был обнаружен, между которыми происходит лишь незначительная миграция, [ 38 ] добавить до 0,0177 квадратных километров (4,4 акра), [ 1 ] или менее одной пятой футбольного поля. [ 20 ]
Население жерлового поля Лунци может вызывать особую озабоченность. Юго-Западный Индийский хребет, в пределах которого он расположен, является одним из срединно-океанических хребтов , распространяющихся медленнее всего , а низкий уровень естественных нарушений связан с экологическими сообществами , которые, вероятно, более чувствительны к разрушениям и медленнее восстанавливаются после них. Медленно распространяющиеся центры могут также создавать более крупные месторождения полезных ископаемых, что делает эти чувствительные районы основными объектами глубоководной добычи полезных ископаемых . Более того, по генетическим показателям популяция Лунци плохо связана с популяциями жерловых полей Кайрей и Солитер, находящихся на расстоянии более 2000 км в пределах Центрально-Индийского хребта. [ 20 ]
Месторождение Солитер находится в исключительной экономической зоне Маврикия, тогда как два других участка находятся в пределах зон за пределами национальной юрисдикции (широко известных как открытое море ) под управлением Международного органа по морскому дну , который выдал лицензии на коммерческую разведку месторождений для обоих месторождений. . Лицензия на месторождение Кайрей принадлежит Германии (2015–2030 гг.), а месторождение Лунци – Китаю (2011–2026 гг.). По состоянию на 2017 год ни для одного из трех объектов не предложено и не введено никаких мер по сохранению. [ 20 ]
он внесен в Красный список исчезающих видов МСОП как вид, находящийся под угрозой исчезновения . С 4 июля 2019 года [ 1 ]
См. также
[ редактировать ]Примечания
[ редактировать ]- ^ По состоянию на 2016 год в Лунци был известен 21 вид, а в 2017 году были описаны два новых брюхоногих моллюска.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д Сигварт, Дж.; Чен, К.; Томас, Э.А. (2019). « Хризомаллон чешуйчатый » . Красный список исчезающих видов МСОП . 2019 : e.T103636217A103636261. doi : 10.2305/IUCN.UK.2019-2.RLTS.T103636217A103636261.en . Проверено 19 ноября 2021 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а к ап ак с как в Чен, Чонг; Линсе, Катрин; Копли, Джонатан Т.; Роджерс, Алекс Д. (2015). «Чешуйчатоногий брюхоногий моллюск»: новый род и вид эндемичных для гидротермальных источников брюхоногих моллюсков (Neomphalina: Peltospiridae) из Индийского океана» . Журнал исследований моллюсков . 81 (3): 322–334. дои : 10.1093/mollus/eyv013 .
- ^ «Познакомьтесь с причудливой морской улиткой, которая строит собственный железный доспех» . 15 ноября 2021 г.
- ^ Сигварт, Джулия Д.; Чен, Чонг; Томас, Элин А.; Олкок, А. Луиза; Бём, Моника; Седдон, Мэри (22 июля 2019 г.). «Красный список может защитить глубоководное биоразнообразие» . Экология и эволюция природы . 3 (8): 1134. Бибкод : 2019NatEE...3.1134S . дои : 10.1038/s41559-019-0930-2 . ISSN 2397-334X . ПМИД 31332328 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а к ап ак с как в В из хорошо топор является тот нет бб до нашей эры др. быть парень бг чб с минет БК с бм млрд быть б.п. БК бр бс БТ этот бв Чен, Чонг; Копли, Джонатан Т.; Линсе, Катрин; Роджерс, Алекс Д.; Сигварт, Джулия Д. (2015). «Сердце дракона: 3D анатомическая реконструкция чешуйчатого брюхоногого моллюска (Mollusca: Gastropoda: Neomphalina) раскрывает его необычную систему кровообращения» . Границы в зоологии . 12:13 . дои : 10.1186/s12983-015-0105-1 . ПМЦ 4470333 . ПМИД 26085836 .
- ^ «Красный список видов, находящихся под угрозой исчезновения» МСОП . Красный список исчезающих видов МСОП . Проверено 28 июля 2019 г.
- ^ Ламберт, Джонатан (22 июля 2019 г.). «Океанская улитка — первое животное, официально находящееся под угрозой исчезновения в результате глубоководной добычи полезных ископаемых» . Природа . 571 (7766): 455–456. Бибкод : 2019Natur.571..455L . дои : 10.1038/d41586-019-02231-1 . ПМИД 31337912 .
- ^ Перейти обратно: а б с д и ж г Довер, Синди Л. Ван; Хамфрис, ЮВ; Форнари, Д.; Кавано, CM; Кольер, Р.; Гоффреди, Шана К.; Хашимото, Дж.; Лилли, доктор медицины; Рейзенбах, Алабама; Шанк, ТМ; Фон Дамм, КЛ; Банта, А.; Галлант, РМ; Гоц, Д.; Грин, Д.; Холл, Дж.; Хармер, ТЛ; Уртадо, Луизиана; Джонсон, П.; МакКинесс, З.П.; Мередит, К.; Олсон, Э.; Пан, Иллинойс; Репа, М.; Вон, Ю.; Янг, CR 3-е место; Вриенгук, Р.К. (2001). «Биогеография и экологическая обстановка гидротермальных источников Индийского океана» . Наука . 294 (5543): 818–23. Бибкод : 2001Sci...294..818V . дои : 10.1126/science.1064574 . ПМИД 11557843 . S2CID 543841 .
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Буше, П. (2014). «Хризомаллон чешуйчатый» . Всемирный реестр морских видов . Проверено 22 апреля 2015 г.
- ^ Перейти обратно: а б Варен, Андерс; Бенгтсон, Стефан; Гоффреди, Шана К.; Дувр, Синди Л. Ван (2003). «Брюхоногий моллюск с горячими жерлами и дермальными склеритами из сульфида железа». Наука . 302 (5647): 1007. doi : 10.1126/science.1087696 . ПМИД 14605361 . S2CID 38386600 .
- ^ Перейти обратно: а б с Чен, Чонг; Линсе, Катрин; Ротерман, Кристофер Н.; Копли, Джонатан Т.; Роджерс, Алекс Д. (2015). «Новый род крупных гидротермальных эндемичных брюхоногих моллюсков (Neomphalina: Peltospiridae)» (PDF) . Зоологический журнал Линнеевского общества (Представлена рукопись). 175 (2): 319–335. дои : 10.1111/zoj.12279 .
- ^ (на японском языке) (2010) «Первое в мире открытие белой чешуйчатой стопы без сульфида железа ~ Результаты новых гидротермальных исследований в Индийском океане ~» Японское агентство морских наук и технологий о Земле , Токийский университет , Университет Коти . пресс-релиз). Проверено 16 июля 2016 г.
- ^ «Новый чешуйчатоногий брюхоногий моллюск обнаружен в Индийском океане; обнаружен белый чешуйчатоногий брюхоногий моллюск» . Южная жареная наука. 1 июля 2011 года . Проверено 16 июля 2016 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л Накамура, Кентаро, Хироми, Дзюнъити, Кен; Ногучи, Такуро, Сугуру; Мацузаки, Такуя, Такадзо; Орихаши, Юджи; Асада, Акира; Кунджул, Мира; Бидесси, Гириш; Тамаки, Шнур, Джоэл М. (2012) . Активность и хемосинтетическая фауна Центрально-Индийского хребта на 18°–20°ю.ш.» . PLOS ONE . 7 (3): e32965. Bibcode : 2012PLoSO...732965N . doi : 10.1371/journal.pone.0032965 . PMC 3303786 . ПМИД 22431990
- ^ Перейти обратно: а б с д и Бедессе, Гириш; Ватанабэ, Хироми; Огура, Томоми; Немото, Сугуру; Яхаги, Такуя; Накагава, Сатоши; Накамура, Кентаро; Такай, Кен; Кунджул, Мира; Мари, Дэниел EP (2013). «Высокая связанность популяций животных в глубоководных гидротермальных полях Центрально-Индийского хребта в связи с его геологической обстановкой» . ПЛОС ОДИН 8 (12):e8 Бибкод : 2013PLoSO... 881570B дои : 10.1371/journal.pone.0081570 . ПМЦ 3864839 . ПМИД 24358117 .
- ^ Перейти обратно: а б Чен, Чонг; Чжоу, Ядун; Ван, Чуньшэн; Копли, Джонатан Т. (2017). «Две новые улитки-пельтоспириды с горячими источниками (Gastropoda: Neomphalina) из гидротермального поля Лунци, юго-западный Индийский хребет» . Границы морской науки . 4 . дои : 10.3389/fmars.2017.00392 . ISSN 2296-7745 .
- ^ Перейти обратно: а б с Тао, Чуньхуэй, Го, Шицин, Ву, Хан, Сицю; . «Первые активные гидротермальные источники в центре сверхмедленного расширения: Юго-Западный Индийский хребет . , Цзян (2012 ) 40 (1) (этапы 1–2) и DY115-20 (этапы 4–7). Стороны: 47–50 Бибкод : 2012Geo....40...47T doi : 10.1130 /G32389.1 .
- ^ Перейти обратно: а б с д и ж г час я Чен, Чонг; Копли, Джонатан Т.; Линсе, Катрин; Роджерс, Алекс Д.; Сигварт, Джулия (2015). «Как у моллюска появилась чешуя: конвергентная эволюция склеритома моллюска» . Биологический журнал Линнеевского общества . 114 (4): 949–954. дои : 10.1111/bij.12462 .
- ^ Роджерс, Алекс Д.; Тайлер, Пол А.; Коннелли, Дуглас П.; Копли, Джон Т.; Джеймс, Рэйчел; Лартер, Роберт Д.; Линсе, Катрин; Миллс, Рэйчел А.; Гарабато, Альфредо Навейра; Панкост, Ричард Д.; Пирс, Дэвид А.; Полунин, Николай В.К.; Герман, Кристофер Р.; Шанк, Тимоти; Бурш-Супан, Филипп Х.; Алкер, Белинда Дж.; Аквилина, Альфред; Беннетт, Сара А.; Кларк, Эндрю; Динли, Роберт Джей-Джей; Грэм, Аластер Г.К.; Грин, Дэррил Р.Х.; Хоукс, Джеффри А.; Хепберн, Лора; Иларио, Ана; Ювенн, Верле А.И.; Марш, Ли; Рамирес-Льодра, Ева; Рид, Уильям Д.К.; Ротерман, Кристофер Н.; Свитинг, Кристофер Дж.; Татье, Свен; Цвирглмайер, Катрин (2012). «Открытие новых глубоководных гидротермальных сообществ в Южном океане и последствия для биогеографии» . ПЛОС Биология . 10 (1): –1001234. дои : 10.1371/journal.pbio.1001234 . ISSN 1545-7885 . ПМК 3250512 . ПМИД 22235194 .
- ^ Перейти обратно: а б с д и ж Сигварт, Джулия Д.; Чен, Чонг; Марш, Ли (2017). «Вредна ли разработка морского дна для моллюсков?». Наутилус . 131 (1): 43–49.
- ^ Перейти обратно: а б Хасимото, Джун; Гамо, Тоситака; Ямагути, Цучида, Синдзи; Ватабе, Хадзиме; Яманака, Китазава, Мицуко (2001) . 10.2108 / . : 18 717–721 (5 ) : zsj.18.717 . S2CID 85582014 doi
- ^ Перейти обратно: а б с Чен, Чонг; Копли, Джонатан Т.; Линсе, Катрин; Роджерс, Алекс Д. (2015). «Низкая связь между популяциями чешуйчатоногих брюхоногих моллюсков (Mollusca: Peltospiridae) в гидротермальных жерлах на Юго-Западном Индийском хребте и Центрально-Индийском хребте». Разнообразие и эволюция организмов . 15 (4): 663–670. дои : 10.1007/s13127-015-0224-8 . S2CID 18521717 .
- ^ Пикрелл, Джон (7 ноября 2003 г.). «Бронированная улитка обнаружена в глубоком море» . Национальные географические новости. Архивировано из оригинала 10 ноября 2003 года . Проверено 16 июля 2016 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л Чен, Чонг; Уэмацу, Кацуюки; Линсе, Катрин; Сигварт, Джулия Д. (2017). «Более чем одним способом: быстрая конвергенция в гидротермальных источниках, показанная трехмерной анатомической реконструкцией Gigantopelta (Mollusca: Neomphalina)» . Эволюционная биология BMC . 17 (1): 62. Бибкод : 2017BMCEE..17...62C . дои : 10.1186/s12862-017-0917-z . ISSN 1471-2148 . ПМК 5333402 . ПМИД 28249568 .
- ^ Сузуки, Йохей; Копп, Роберт Э.; Коруге, Тошихиро; Суга, Акинобу; Такай, Кен; Цучида, Синдзи; Одзаки, Нориаки; Эндо, Казуёси; Хасимото, Джун; Като, Ясухиро; Мизота, Титоши; Хирата, Такафуми; Тиба, Хитоши; Нилсон, Кеннет Х.; Хорикоши, Коки; Киршвинк, Джозеф Л. (2006). «Образование склерита у «чешуйчатых» брюхоногих моллюсков гидротермальных источников - возможный контроль биоминерализации сульфида железа животным» (PDF) . Письма о Земле и планетологии . 242 (1–2): 39–50. Бибкод : 2006E&PSL.242...39S . дои : 10.1016/j.epsl.2005.11.029 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м Гоффреди, Шана К.; Варен, Андерс; Сирота, Виктория Дж.; Довер, Синди Л. Ван; Вриенгук, Роберт К. (5 мая 2004 г.). «Новые формы структурной интеграции микробов и брюхоногих моллюсков Индийского океана» . Прикладная и экологическая микробиология . 70 (5): 3082–3090. Бибкод : 2004ApEnM..70.3082G . дои : 10.1128/АЕМ.70.5.3082-3090.2004 . ПМК 404406 . PMID 15128570 .
- ^ Перейти обратно: а б с д Яо, Хаймин; Дао, Мин; Имхольт, Тимоти; Хуанг, Джейми; Уиллер, Кевин; Бонилья, Алехандро; Суреш, Субра; Ортис, Кристина (2010). «Защитные механизмы железной брони глубоководного брюхоногого моллюска гидротермальных источников» . ПНАС . 107 (3): 987–992. Бибкод : 2010PNAS..107..987Y . дои : 10.1073/pnas.0912988107 . ПМК 2808221 . ПМИД 20133823 .
- ^ «Железные доспехи улитки на глазах у военных» . Новости Си-Би-Си. 19 января 2010 г. Проверено 16 июля 2016 г.
- ^ Перейти обратно: а б с Накагава, Сигэру, Ёсихиро; Мураками, Сюн-ичи; Фуджиёси, Саяка; Маэда, Макита, Хироко; Нисимура, Шин-Ичиро; Вацуджи, Томо-о; Кен (2014). «Союзник с панцирными улитками: полный геном гаммапротеобактериального эндосимбионта» . Журнал ISME 8 ( 1): 40–51. Бибкод : 2014ISMEJ...8... 40N doi : 10.1038 PMC 3869010. / ismej.2013.131 PMID 23924784 .
- ^ Дистел, Дэниел Л.; Альтамия, Марвин А.; Линь, Чжэньцзянь; Шипвей, Дж. Рубен; Хан, Эндрю; Фортеза, Имельда; Антемано, Ровена; Лимбако, Ма Гвен Дж. Пеньяфлор; Тебо, Элисон Г.; Дешавес, Ранде; Альбано, Джули; Розенберг, Гэри; Консепсьон, Гизела П.; Шмидт, Эрик В.; Хейгуд, Марго Г. (17 апреля 2017 г.). «Открытие хемоавтотрофного симбиоза у гигантского корабельного червя Kuphus polythalamia (Bivalvia: Teredinidae) расширяет теорию деревянных ступенек» . Труды Национальной академии наук . 114 (18): Е3652–Е3658. Бибкод : 2017PNAS..114E3652D . дои : 10.1073/pnas.1620470114 . ISSN 1091-6490 . ПМЦ 5422788 . ПМИД 28416684 .
- ^ Перейти обратно: а б Накамура, Кентаро; Такай, Кен (2015). «Гидротермальные системы Индийского океана: гидротермальная активность морского дна, физические и химические характеристики гидротермальных флюидов и биологические сообщества, связанные с жерлами». В Исибаши Ж.-и.; и др. (ред.). Подводная биосфера, связанная с гидротермальными системами . Спрингер, Токио. стр. 147–161. дои : 10.1007/978-4-431-54865-2_12 . ISBN 9784431548645 .
- ^ Перейти обратно: а б с Довер, Синди Ван (2002). «Трофические взаимоотношения беспозвоночных на гидротермальном жерлом Кайрей (Центрально-Индийский хребет)». Морская биология . 141 (4): 761–772. Бибкод : 2002МарБи.141..761В . дои : 10.1007/s00227-002-0865-y . S2CID 189819863 .
- ^ Перейти обратно: а б с д «Обнаружена обширная популяция «редкого» чешуйчатого брюхоногого моллюска» . Японское агентство по морским наукам и технологиям о Земле , Университет Хоккайдо , Аквариум Эносима . 30 ноября 2009 года . Проверено 16 июля 2016 г.
- ^ Перейти обратно: а б с Ватанабэ, Хироми; Бидесси, Гириш (2015). «Вентфауна Центрально-Индийского хребта». В Исибаши Ж.-и.; и др. (ред.). Подводная биосфера, связанная с гидротермальными системами . Спрингер, Токио. стр. 205–212. дои : 10.1007/978-4-431-54865-2_16 . ISBN 9784431548645 . S2CID 127157740 .
- ^ Джонсон, Шеннон Б.; Варен, Андерс; Танниклифф, Верена; Довер, Синди Ван; Пшеница, К. Джеффри; Шульц, Томас Ф.; Вриенгук, Роберт К. (04 мая 2015 г.). «Молекулярная систематика и наименование пяти загадочных видов улиток Alviniconcha (Gastropoda: Abyssochrysoidea) из гидротермальных источников» . Систематика и биоразнообразие . 13 (3): 278–295. Бибкод : 2015SyBio..13..278J . дои : 10.1080/14772000.2014.970673 . ISSN 1477-2000 . S2CID 85253352 .
- ^ Перейти обратно: а б с Окутани, Такаши; Хасимото, Джун; Сасаки, Такенори (2004). «Новые таксоны брюхоногих моллюсков из гидротермального источника (поле Кайрей) в центральной части Индийского океана» (PDF) . Венера 63 (1–2): 1–10. Архивировано из оригинала (PDF) 0 октября 2013 г.
- ^ Перейти обратно: а б Копли, Дж. Т.; Марш, Л.; Гловер, АГ; Хюнербах, В.; Най, В.Е.; Рид, WDK; Свитинг, CJ; Уигэм, Б.Д.; Виклунд, Х. (2016). «Экология и биогеография мегафауны и макрофауны первых известных глубоководных гидротермальных источников на сверхмедленно спрединговом Юго-Западном Индийском хребте» . Научные отчеты . 6 : 39158. Бибкод : 2016NatSR...639158C . дои : 10.1038/srep39158 . ISSN 2045-2322 . ПМК 5155287 . ПМИД 27966649 .
- ^ Сигварт, Джулия Д. (зима 2017 г.). «Глубоководная охрана и« чешуйчатоногие брюхоногие моллюски » » (PDF) . Щупальца . 25 : 39–40.
Внешние ссылки
[ редактировать ] СМИ, связанные с Chrysomallon sqamiferum, на Викискладе?
СМИ, связанные с Chrysomallon sqamiferum, на Викискладе?