Членистонный глаз

Прилагательные глаза являются наиболее распространенной формой глаза и, по -видимому, являются наследственной формой сложного глаза . Они встречаются во всех группах членистоногих , хотя они могли развиваться не раз в этом филоме. [ 1 ] Некоторые аннолиды и двустворчатые моллюски также имеют приложения. Они также обладают Limulus , подковообразным крабом, и есть предположения, что другие хелимераты разработали свои простые глаза путем уменьшения от составной отправной точки. [ 1 ] Некоторые гусеницы , похоже, развили сложные глаза от простых глаз противоположного способа. [ Цитация необходима ]
Членистоногие , обладающие составными глазами, обладали составными глазами , но тип и происхождение этого глаза варьируются между группами, а некоторые таксоны по вторичному разрабатывались простыми глазами. Развитие органа через происхождение можно оценить путем сравнения групп, которые рано разветвлялись, такие как бархатный червь и подковообразный краб с усовершенствованным состоянием глаза, обнаруженным у насекомых и других производных членистоногих.
Глаза и функции
[ редактировать ]Большинство членистоногих имеют по крайней мере один из двух типов глаз: боковые составные глаза и меньшие медианные глазки, которые являются простыми глазами. [ 2 ] Когда оба присутствуют, два типа глаз используются в концерте, потому что каждый имеет свое собственное преимущество. [ 3 ] насекомых Некоторые личинки , например, гусеницы , имеют другой тип простых глаз, известный как Stemmata . Эти глаза обычно обеспечивают только грубое изображение, но (как у личинок пиломалека ) они могут обладать разрешением способностей 4 градусов дуги, быть чувствительными к поляризации и способны повысить свою абсолютную чувствительность ночью в 1000 или более. [ 4 ] Летающие насекомые могут оставаться ровными с любым типом хирургического снятия глаз, но эти два типа объединяются, чтобы обеспечить лучшую производительность. [ 3 ] Оцелл может обнаружить более низкие уровни света, [ А ] [ 5 ] и имеют более быстрое время отклика, в то время как составные глаза лучше обнаружают края и способны формировать изображения. [ 3 ]
Анатомическое распределение сложных глаз
[ редактировать ]-
 Многие насекомые, такие как женская табанусная линеола , показывают дихоптические составные глаза
Многие насекомые, такие как женская табанусная линеола , показывают дихоптические составные глаза -
 Мужской табанус Lineola имеет голоптические составные глаза, с дорсальной ommatidia больше, чем вентральная ommatidia
Мужской табанус Lineola имеет голоптические составные глаза, с дорсальной ommatidia больше, чем вентральная ommatidia -
 У некоторых мужских майков глаза разделены на отдельные органы для различных визуальных функций
У некоторых мужских майков глаза разделены на отдельные органы для различных визуальных функций -
 Хихоптические глаза речного краба типичны для зрелых больших ракообразных. Отражение фотографа в разных областях поверхности каждого глаза указывает на основу стереоскопического зрения
Хихоптические глаза речного краба типичны для зрелых больших ракообразных. Отражение фотографа в разных областях поверхности каждого глаза указывает на основу стереоскопического зрения -
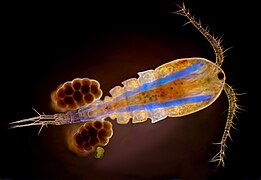 Многие винобочепод и другие маленькие ракообразные глаза объединяются в один циклоптический составной глаз в середине головы, видимый здесь как темное пятно между основаниями антенн.
Многие винобочепод и другие маленькие ракообразные глаза объединяются в один циклоптический составной глаз в середине головы, видимый здесь как темное пятно между основаниями антенн.





Большинство видов членистоногих с сложными глазами имеют только два глаза, которые расположены отдельно и симметрично, по одному на каждой стороне головы. Эта договоренность называется Dichoptic . Примеры включают большинство насекомых и большинство крупных видов ракообразных, таких как крабы. Многие другие организмы, такие как позвоночные и цефалопода, являются аналогично и аналогично дихоптическим, что является общим состоянием у животных, которые являются членами Bilateria и имеют функционально сложные глаза. Тем не менее, есть вариации этой схемы. В некоторых группах животных, чьи предки первоначально были дихоптическими, глаза современных видов могут быть переполнены вместе в срединном плане; Примеры включают в себя многие археогната . В крайних случаях такие глаза могут фактически превратиться в один глаз, как и в некоторых из топодов , особенно в роде Cyclops . Одним из терминов для такого расположения глаз является циклоптическим .
С другой стороны, некоторые способы жизни повышают остроту зрения , которые в сложных глазах требуют большего количества Ommatidia , что, в свою очередь, требует больших сложных глаз. Результатом является то, что глаза занимают большую часть доступной поверхности головы, уменьшая площадь фрон и вершину и толпят глазко, если таковые имеются. Хотя технически такие глаза все еще могут считаться дихоптиками, результат в крайнем случае заключается в том, что границы таких глаз встречаются, эффективно формируя крышку над большинством головы. Такая анатомия называется голоптической . Эффективные примеры можно увидеть в анизоптере и в различных мухах, таких как некоторые Acroceridae и Tabanidae .
Напротив, необходимость в определенных функциях может не потребовать чрезвычайно больших глаз, но требуют большого разрешения и хорошего стереоскопического видения точных атак. Хорошие примеры можно увидеть в Mantodea и Mantispidae , в которых видение добычи от определенной Ommatidia в обоих сложных глазах одновременно указывает на то, что он находится в правильном положении, чтобы вырвать в засаде близкого расстояния. Их глаза соответственно расположены в хорошем положении для всестороннего зрения, а также особую концентрацию на передней срединной плоскости. Индивидуальные омматидии направлены во всех направлениях, и, соответственно, можно увидеть темное пятно ( псевдопупиль ), показывая, какая ommatidia покрывает это поле зрения; С любой позиции на средней плоскости, и нигде больше, два темных пятна симметричны и идентичны.
Иногда потребности в остроте зрения в различных функциях конфликта, и различные части глаз могут быть адаптированы к отдельным функциям; Например, Gyrinidae проводят большую часть своей взрослой жизни на поверхности воды, и их два сложных глаза разделяются на четыре половинки, две для подводного зрения и два для зрения в воздухе. Опять же, особенно в некоторых Diptera, Ommatidia в разных областях голоптического мужского глаза может заметно различаться по размеру; Верхняя Ommatidia, как правило, больше. В случае некоторых эфемероптер эффект настолько преувеличен, что верхняя часть глаза повышена, как воскресный кекс, в то время как его нижняя часть, служащая для обычного зрения, выглядит как отдельный орган.
Составные глаза часто не являются полностью симметричными с точки зрения количества Ommatidia. Например, асимметрии были указаны в пчелах [ 6 ] и различные мухи. [ 7 ] Эта асимметрия коррелировала с поведенческой латерализацией у муравьев (поворот смещения). [ 8 ]
Генетические контроли
[ редактировать ]В фруктовой мух Drosophila melanogaster (наиболее изученные виды членистоногих в отношении биологии развития), одними из наиболее важных генов для паттерна глаз насекомых являются гомологи PAX6 ( ( EY) и близнец бездимочных игрушка ) . Вместе эти гены стимулируют пролиферацию клеток на ранних этапах развития глаз. Потеря любого из этих генов приводит к неудаче образования глаз. Активность EY и игрушки включает в себя активацию генов определения сетчатки SINE Oculis ( SO ) и отсутствующих глаз ( EYA ), которые образуют белковый комплекс, который регулирует транскрипцию нижестоящих генов -мишеней. [ 9 ] После этого две визуальные системы D. melanogaster по -разному. Паттерн передней головки контролируется ортодентином ( OTD ) , геном гомеобокс , который разграничивает сегменты от верхней среды головы до более латеральных аспектов. Оцли находятся в районе ОТД , а нарушение ОТД приводит к потере глаз, но не влияет на сложные глаза. [ 10 ] транскрипционных факторов Наоборот, для таблицы ( DAC ) необходима для паттерна составных глаз, но мутанты, в которых не хватает ЦАП , не демонстрируют потери глаз. [ 11 ] Различные опсины используются в стеклянных глазах сложных глаз. [ 12 ]
Визуальные системы Chelicerata (родственная группа до оставшейся членистоногих) менее хорошо изучены. Было показано, что гомологи многих генов паттерна глаз различно экспрессируются в глазах разных видов пауков, но функциональная значимость этих изменений в экспрессии не совсем понятна из -за отсутствия функциональных данных. [ 13 ] [ 14 ] Кроме того, в подковообразных и пауках было показано, что гомологи PAX6 не выражаются так же, как их аналоги у насекомых, что позволяет предположить, что PAX6 может не потребоваться в качестве переключателя с паттерном для глаз на верхнем уровне в хелисертах. [ 15 ] [ 16 ] Большинство функциональных данных о паттерне глаз в chelicerata взята из Opilio Phalangium phalangium daddy-longlegs , который использовался, чтобы показать, что глаза, отсутствующие, играет консервативную роль в паттерне как визуальных системах этого вида (пример сохранения генной функции , относительно насекомых) и что такшата влияет на паттерн боковых глаз, но не медианные глаза (еще один пример сохранения). [ 17 ]
Эволюция
[ редактировать ]гексаподы В настоящее время считается, что попадают в группу ракообразной короны; В то время как молекулярная работа проложила путь для этой ассоциации, их морфология и развитие глаз также заметно схожи. [ 18 ] Глаза поразительно отличаются от мирипод , которые традиционно считались родственной группой для гексаподы.
И глазок, и сложных глаз, вероятно, присутствовали в последнем общем холенистоне -прессе, [ 19 ] и может быть апоморфным с глазками в других филах, [ 20 ] такие как Аннелиды. [ 21 ] Среда стекла присутствует в хеликратах и глубине ; Боковые гласты также присутствуют в Челикрасе. [ 20 ]
Источник
[ редактировать ]Никакие ископаемые организмы не были идентифицированы как похожие с последним общим предком членистоногих; Следовательно, глаза, одержимые первым членистоножным, остаются вопросом гипотезы. Самая большая подсказка в их внешности получена от линии Onichophorans : группы STEM группы, которая расходилась вскоре перед первыми настоящими членистоногими. Глаза этих существ прикреплены к мозгу, используя нервы, которые входят в центр мозга, и есть только одна область мозга, посвященная зрению. Это похоже на проводку средних глаз (маленькие простые глаза), обладающие многими членистоногими; Глаза также следуют аналогичному пути через раннее развитие организмов. Это говорит о том, что глаза онийхофораны получены из простых глазей, а отсутствие других структур глаз подразумевает, что у наследственного членистоногих не хватало сложных глаз, и использовались только средние глазки для ощущения света и темноты. [ 2 ]

Однако противоречивые взгляды отмечают, что сложные глаза появились во многих ранних членистоногих, включая трилобиты и европтериды. Это говорит о том, что составной глаз мог развиваться после расщепления линий онхофорана и членистоногих, но до излучения членистоногих. [ 20 ] Эта точка зрения поддерживается, если позиция STEM-Artropod поддерживается для компонентных глаз, несущих кембрийские организмы, такие как Radiodontids . Еще одна альтернатива заключается в том, что сложные глаза независимо развивались, несколько раз внутри членистоногих. [ 21 ]
Вероятно, в CHARTPROOD Concestor, вероятно, была только одна пара глазей, поскольку окаменелости в кембрийском лобоподе отображаются одну пару. И хотя многие членистоногие сегодня имеют три, четыре или даже шесть, отсутствие общего пути предполагает, что пара является наиболее вероятным исконным состоянием. Раковые ракообразные и насекомые в основном имеют три стекла, что позволяет предположить, что такая формация присутствовала в их концесторе. [ 2 ]
Считается вероятным, что составной глаз возник в результате «дублирования» отдельных глазелей. [ 20 ] В свою очередь, рассеяние сложных глаз, по -видимому, создало большие сети, казалось бы, независимых глаз у некоторых членистоногих, таких как личинки некоторых насекомых. [ 20 ] У некоторых других насекомых и мириаподов боковые глазки, по -видимому, возникли благодаря уменьшению боковых составных глаз. [ 20 ]
Трилобитовые глаза
[ редактировать ]

Глаза трилобитов появились в трех формах, называемых Холохроала , Шизохроала и Абатохроала глазами . Морфология глаз трилобитов полезна для вывода их способа жизни и может функционировать как индикаторы условий палео-среда. [ 22 ]
Голохроал глаз был наиболее распространенным и самым примитивным. Он состоял из многих небольших линз - от 100 до 15 000 - покрытых одной из мембраны роговицы. Это был самый древний вид глаз. Эта морфология глаз была обнаружена в кембрийских трилобитах (самые ранние) и выжила до исчезновения перм . [ 22 ]
Более сложный шизохроал глаз был обнаружен только в одном подпоряле трилобита, факопине (ордовик-силурийский). У современных животных нет точного аналога для шизохроала у современных животных, но несколько сходная структура глаз обнаруживается у взрослых насекомых -мужчин в порядке Strepsiptera . Шизохроал Глаза развивались как улучшение в Холохроале ; Они были более мощными, с перекрывающимися визуальными полями и были особенно полезны для ночного зрения и, возможно, для цвета и глубины . Шизохроал глаз имеет до 700 больших линз (большие по сравнению с линзами Holochroal ). У каждого объектива есть роговица, и у каждого есть отдельная склера , которая отделяет ее от окружающих линз. Множественные линзы для глаз были построены из одного кристалла кальцита . Ранние конструкции глаз Schizochroal кажутся случайными и нерегулярными - возможно, ограниченными геометрическими осложнениями упаковки линз идентичных размеров на изогнутой поверхности. Позже шизохроал -глаза имели размер градуированной линзы. [ 22 ]
Абатохроал Глаз является третьей морфологией глаз трилобитов, но он обнаружился только в эодисцине . Эта форма глаза состояла из 70 гораздо меньших линз. Роговица разделяла каждую линзу, а склера на каждой линзе заканчивалась поверх каждой роговицы. [ 22 ]
Подкова краб
[ редактировать ]
Подковообразные крабы традиционно использовались при исследованиях глаз, потому что он имеет относительно большую оматидию с большими нервными волокнами (что делает их легкими для экспериментов). Это также падает приближается к основанию хелицератов ; Считается, что его глаза представляют собой наследственное состояние, потому что они так мало изменились в течение эволюционного времени. Большинство других живых хелимератов потеряли свои боковые сложные глаза, развивая простые глаза на их месте, которые различаются по количеству. [ 24 ] В скорпионах встречается до пяти пар латеральных глаз, тогда как три пары боковых глаз типичны для тетрапульмоната (например, пауки ; Amblypygi ). [ 25 ]
Подковообразные крабы имеют два больших составных глаза по бокам головы. Дополнительный простой глаз расположен в задней части каждой из этих структур. [ 24 ] В дополнение к этим очевидным структурам, в нем также есть два меньших стекла, расположенных в среднем переднем фронте его панциря, что может быть внешне ошибочно принято за ноздри. [ 24 ] Еще простой глаз расположен под ними, на нижней части панциря; Этот глаз изначально сочетается во время эмбриональных стадий и предохранителей позже в разработке. [ 24 ] [ 15 ] Еще одна пара простых глаз расположена прямо перед ртом. [ 24 ] Простые глаза, вероятно, важны на эмбриональных или личиночных стадиях организма, причем сложные глаза и срединные глазки становятся доминирующими органами прицела во время взрослой жизни. [ 24 ] Эти глазки менее сложны и, вероятно, менее получены, чем у мандибулаты . [ 20 ] В отличие от составных глаз трилобитов, глазки подковообразных крабов имеют треугольную форму; У них также есть генеративная область на своей базе, но это удлиняется со временем. Следовательно, единственным омматидием на вершине треугольника был оригинальный «глаз» личиночного организма, с последующими рядами, добавленными по мере роста организма. [ 18 ]
Насекомые и ракообразные
[ редактировать ]
Обычно считается, что насекомые являются кладкой внутри ракообразной, и что ракообразные монофилетические . Это согласуется с наблюдением, что их глаза развиваются очень похожими. В то время как большинство ракообразных и некоторых личинок насекомых обладают только простыми медианными глазами, такими как Болвиг Органы дрозофилы . и глаз Науплера большинства ракообразных, в нескольких группах есть личинки с простыми или составленными латеральными глазами Соединенные глаза взрослых развиваются в области головы, отделенной от региона, в которой развивается медианный глаз личинки. [ 18 ] Новые омматидии добавляются в полукруглые ряды в задней части глаза; На первом этапе роста это приводит к тому, что индивидуальная Ommatidia будет квадратной, но позже в развитии они становятся шестиугольными. Гексагональный рисунок станет видимым только тогда, когда панцирь сцены с квадратными глазами будет лишена. [ 18 ]
Несмотря на то, что у некоторых видов ракообразных и у некоторых насекомых возникают глаза на цветоносы , только некоторые ракообразные, такие как крабы, приносят глаза на сочлененные цветоносы, которые позволяют складываться глазами с неприятностей.
Бесчисленные мириаподы
[ редактировать ]
Большинство бесчисленных мириапод несет в стволах - одноразовые глаза, которые, как полагают, развивались путем восстановления составного глаза. [ 20 ] Тем не менее, члены рода чилопод Scutigera имеют сложный глаз, который состоит из аспектов [ 26 ] и нет, как это имело более ранние интерпретации, кластерных стволов. [ 21 ] Считалось, что они растут в рядах, вставленные между существующими рядами стекла. [ 18 ]
Смотрите также
[ редактировать ]- Моллас глаз
- Темный глаз
- Простой глаз в беспозвоночных
- Видение в рыбе
- Оптическая доля (членистоногие)
Сноски
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а беременный Земля, Майкл Ф.; Фернальд, Рассел Д. (1992), «Эволюция глаз», Ежегодный обзор нейробиологии , 15 (1): 1–29, doi : 10.1146/annurev.ne.15.030192.000245 , PMID 1575438 .
- ^ Jump up to: а беременный в Mayer, G. (2006). «Структура и развитие онхофорановых глаз: что такое наследственный визуальный орган у членистоногих?». Членистоногие структура и развитие . 35 (4): 231–245. Bibcode : 2006artsd..35..231m . doi : 10.1016/j.asd.2006.06.003 . PMID 18089073 .
- ^ Jump up to: а беременный в Тейлор, Чарльз П. (1981). «Вклад составных глаз и глаз в рулевое управление саранчой в полете. I. Поведенческий анализ». J. Exp. Биол . 93 : 1–18. doi : 10.1242/jeb.93.1.1 . [ Полная цитата необходима ]
- ^ Meyer-Rochow, VB (1974). «Структура и функция личиночного глаза личинки пилот -личинки ». Журнал физиологии насекомых . 20 (8): 1565–1591. doi : 10.1016/0022-1910 (74) 90087-0 . PMID 4854430 .
- ^ Уилсон, М. (1978). «Функциональная организация саранчика». Журнал сравнительной физиологии . 124 (4): 297–316. doi : 10.1007/bf00661380 . S2CID 572458 .
- ^ Seidl, R.; Кайзер В. (1981). «Размер поля зрения, бинокулярный домен и ommatidial массив сложных глаз у рабочих медоносных пчел». J. Comp. Физиол . 143 : 17–26. doi : 10.1007/bf00606065 . S2CID 41150125 .
- ^ Beersma, DG; и др. (1977). «Решетка сетчатки, визуальное поле и бинокулярность у мух». J. Comp. Физиол . 119 (3): 207–220. doi : 10.1007/bf00656634 . S2CID 6384399 .
- ^ Охота, эр; и др. (2018). «Асимметричное количество Ommatidia и поведенческая латерализация у муравей Темноторакс Альбипеннис » . Научные отчеты . 8 (5825): 5825. Bibcode : 2018natsr ... 8.5825h . doi : 10.1038/s41598-018-23652-4 . PMC 5895843 . PMID 29643429 .
- ^ Картю, Ричард В. (август 2007 г.). «Образование рисунка в глазах дрозофилы» . Текущее мнение в области генетики и развития . 17 (4): 309–313. doi : 10.1016/j.gde.2007.05.001 . PMC 2693403 . PMID 17618111 .
- ^ Royet, J; Finkelstein, R (1995), «Образование паттернов при развитии головы Drosophila: роль гена ортодентичного гомеобокс» (PDF) , разработка , 121 (11): 3561–3572, doi : 10.1242/dev.121.11.3561 , PMID 8582270
- ^ Mardon, G.; Соломон, NM, NM; Рубин, GM, GM (декабрь 1994). « Dachshund кодирует ядерный белок, необходимый для нормального развития глаз и ног у дрозофилы». Разработка . 120 (12): 3473–3486. doi : 10.1242/dev.120.12.3473 . ISSN 0950-1991 . PMID 7821215 .
- ^ С. Верхан, Уильям А. Зукер; Харрис, Вашингтон; Киршфельд, К; Wehrhahn, C; Zuker, CS (1988), «Целевая мисекспрессия гена drosophila opsin приводит к измененной визуальной функции», Nature , 333 (6175): 737–41, Bibcode : 1988nater.333..737f , DOI : 10.1038/337737A0 , PMID 2452330 , S2CID 4248264
- ^ Шомбург, Кристоф; Турецек, Наташа; Шахт, Магдалена Инес; Шнайдер, Джулия; Кирфель, Филипп; PRPIC, Никола-Майкл; Posnien, Nico (декабрь 2015 г.). «Молекулярная характеристика и эмбриональное происхождение глаз в общем доме паука Parasteatoda tepidariorum» . Эводево . 6 (1): 15. doi : 10.1186/s13227-015-0011-9 . ISSN 2041-9139 . PMC 4450840 . PMID 26034574 .
- ^ Самади, Лейли; Шмид, Аксель; Эрикссон, Бо Йоаким (декабрь 2015 г.). «Дифференциальная экспрессия генов определения сетчатки в главных и вторичных глазах Кубинена Салеи Кизерлинг (1877)» . Эводево . 6 (1): 16. doi : 10.1186/s13227-015-0010-x . ISSN 2041-9139 . PMC 4450993 . PMID 26034575 .
- ^ Jump up to: а беременный Блэкберн, Дэвид С.; Конли, Кевин У.; Plachetzki, David C.; Kempler, Карен; Баттель, Барбара-Энн; Браун, Надин Л. (август 2008 г.). «Выделение и экспрессия PAX6 и атональных гомологов в американском подковообразном крабе, Limulus polyphemus» . Динамика развития . 237 (8): 2209–2219. doi : 10.1002/dvdy.21634 . ISSN 1058-8388 . PMC 2577597 . PMID 18651657 .
- ^ Фридрих, Маркус (декабрь 2022 г.). «Наконец -то вступление в ясное зрение: происхождение и полученные события во время развития визуальной системы хелицерата» . Биологии . 44 (12): E2200163. doi : 10.1002/bies.202200163 . ISSN 0265-9247 . PMID 36317531 . S2CID 253246397 .
- ^ Гейнетт, Гилхерм; Клеменц, Бенджамин С.; Blaszczyk, Pula; Сеттон, Эмили В.В.; Мурайама, Габриэль П.; Виллемарт, Родриго; Gavish-Regev, Efrat; Шарма, Прахант П. (февраль 2024 г.). «Редиментарные органы изменяют размещение ископаемых в древней группе наземных хелилисов» . Текущая биология . Doi : 10.1016/j.cub.2024.02.011 . PMID 38401545 . S2CID 267821504 .
- ^ Jump up to: а беременный в дюймовый и Harzsch, S.; Hafner, G. (2006), «Эволюция развития глаз в членистоногих: филогенетические аспекты», Структура и развитие членистоногих , 35 (4): 319–340, Bibcode : 2006Artsd..35..319H , doi : 10.1016/j. ASD.2006.08.009 , HDL : 11858/00-001M-0000-0012-A87C-4 , PMID 18089079
- ^ Фридрих, М. (2006), «Древние механизмы развития органов визуального смысла, основанные на сравнении генных сетей, контролирующих личиночные глаза, глаз, и спецификации составных глаз у дрозофилы» (PDF) , структура и развитие членистоногих , 35 (4): 357 –378, Bibcode : 2006Artsd..35..357f , doi : 10.1016/j.asd.2006.08.010 , PMID 18089081
- ^ Jump up to: а беременный в дюймовый и фон глин час Bitsch, C.; Bitchch, J. (2005), «Эволюция структуры глаз и филогения членистоногих» , Raceancean -проблемы , 16 : 185–214, doi : 10.1201/9781420037548 , ISBN 978-0-8493-3498-6
- ^ Jump up to: а беременный в Paulus, HF (2000), «Филогения мириаподы-карастацеа-Insecta: новая попытка с использованием структуры фоторецепторов*», Журнал зоологической систематики и эволюционных исследований , 38 (3): 189–208, doi : 10.1046/j.1439, 38 (3): 189–208 , DOI: 10.1046/ -0469.2000.383152.x
- ^ Jump up to: а беременный в дюймовый Clarkson, Euan (2009). Беспозвоночные палеонтология и эволюция (4 -е изд.). Джон Уайли и сыновья. ISBN 9781444313321 - через Google Books.
- ^ Barlow, RB (2009). «Видение в подковообразных крабах» . В Танакреди, JT; Botton, ML; Смит, Д. (ред.). Биология и сохранение подковообразных крабов . Спрингер. С. 223–235. ISBN 9780387899589 - через Google Books.
- ^ Jump up to: а беременный в дюймовый и фон Battelle, Ba (декабрь 2006 г.), «Глаза Limulus polyphemus (Xiphosura, Chelicerata) и их афферентные и эфферентные проекции», « Структура и развитие членистоногих , 35 (4): 261–74, Bibcode : 2006Artsd..35 .. 261b , doi : 10.1016/j.asd.2006.07.002 , PMID 18089075 .
- ^ Митер, Себастьян Т.; Данлоп, Джейсон А. (июль 2016 г.). «Боковая эволюция глаз в арахнидах» . Арахнология . 17 (2): 103–119. doi : 10.13156/arac.2006.17.2.103 . ISSN 2050-9928 . S2CID 89428386 .
- ^ Мюллер, Chg; Розенберг, Дж.; Рихтер, с.; Meyer-Rochow, VB (2003). «Составной глаз Scutigera coleoptrata ( Linnaeus, 1758 ) (Chilopoda; notostigmophora): ультраструктурное повторное расследование, которое добавляет поддержку концепции Mandibulata». Zoomorphology . 122 (4): 191–209. doi : 10.1007/s00435-003-0085-0 . S2CID 6466405 .
