кладистика
Кладистика ( / k l ə ˈ d ɪ s t ɪ k s / klə- DISS -tiks ; от древнегреческого κλάδος (kládos) 'ветвь') [ 1 ] — это подход к биологической классификации , при котором организмы распределяются по группам (« кладам ») на основе гипотез о недавнем общем происхождении . Доказательством гипотетических взаимоотношений обычно являются общие производные характеристики ( синапоморфии ), которых нет у более отдаленных групп и предков. Однако с эмпирической точки зрения общие предки - это выводы, основанные на кладистической гипотезе взаимоотношений таксонов, состояния признаков которых можно наблюдать. Теоретически последний общий предок и все его потомки составляют (минимальную) кладу. Важно отметить, что все потомки остаются в своей всеобъемлющей родовой кладе. Например, если бы термины «черви» или «рыбы» использовались в строгих кладистических рамках, эти термины включали бы людей. Многие из этих терминов обычно используются парафилетически , вне кладистики, например, как « степень », точное определение которых бесполезно. [ почему? ] особенно если включить вымершие виды. Радиация приводит к образованию новых субкладов путем бифуркации, но на практике половая гибридизация может размыть очень тесно связанные группы. [ 2 ] [ 3 ] [ 4 ] [ 5 ]
Как гипотеза, клада может быть отвергнута только в том случае, если некоторые группировки были явно исключены. Тогда может оказаться, что исключенная группа действительно произошла от последнего общего предка группы и, таким образом, возникла внутри группы. («Эволюционировало из» вводит в заблуждение, поскольку в кладистике все потомки остаются в группе предков). Чтобы сохранить только действительные клады, после обнаружения того, что группа таким образом является парафилетической, следует либо предоставить кладе такие исключенные группы, либо группу следует упразднить. [ 6 ]
Ветви до расхождения к следующей значимой (например, существующей) сестре считаются стволовыми группами клады, но в принципе каждый уровень стоит сам по себе и имеет уникальное имя. Для полностью раздвоенного дерева добавление группы к дереву также добавляет дополнительную (именованную) кладу и новый уровень в этой ветви. В частности, вымершие группы всегда помещаются в боковую ветвь, без учета того, был ли найден фактический предок других групп.
Методы и номенклатура кладистики применялись и к другим дисциплинам, помимо биологии. (См. филогенетическую номенклатуру .)
Результаты кладистики создают трудности для таксономии , поскольку ранг и (родовое) наименование установленных группировок могут оказаться противоречивыми.
Кладистика в настоящее время является наиболее часто используемым методом классификации организмов. [ 7 ]
История
[ редактировать ]


Оригинальные методы, используемые в кладистическом анализе и школе таксономии, заимствованы из работ немецкого энтомолога Вилли Хеннига , который называл это филогенетической систематикой (также название его книги 1966 года); но термины «кладистика» и «клада» были популяризированы другими исследователями. Кладистика в первоначальном смысле относилась к определенному набору методов, используемых в филогенетическом анализе, хотя теперь иногда она используется для обозначения всей области. [ 8 ]
То, что сейчас называется кладистическим методом, появилось еще в 1901 году в работе Питера Чалмерса Митчелла о птицах. [ 9 ] [ 10 ] а затем Роберт Джон Тиллард (по насекомым) в 1921 году, [ 11 ] и В. Циммерманн (для растений) в 1943 г. [ 12 ] Термин « клада » был введен в 1958 году Джулианом Хаксли после того, как его ввел Люсьен Куэно в 1940 году. [ 13 ] «кладогенез» в 1958 г., [ 14 ] «кладистика» Артура Кейна и Харрисона в 1960 году, [ 15 ] «Кладист» (для приверженца школы Хеннига) Эрнста Майра в 1965 году, [ 16 ] и «кладистика» в 1966 г. [ 14 ] Хенниг называл свой подход «филогенетической систематикой». Со времени его первоначальной формулировки и до конца 1970-х годов кладистика как аналитический и философский подход к систематике конкурировала с фенетикой и так называемой эволюционной таксономией . Фенетику в это время отстаивали числовые систематики Питер Снит и Роберт Сокал , а эволюционную таксономию - Эрнст Майр . [ 17 ]
Первоначально задуманная, хотя бы по сути, Вилли Хеннигом в книге, опубликованной в 1950 году, кладистика не процветала до ее перевода на английский язык в 1966 году (Lewin 1997). Сегодня кладистика является наиболее популярным методом построения филогенетических деревьев на основе морфологических данных.
В 1990-х годах разработка эффективных методов полимеразной цепной реакции позволила применить кладистические методы к биохимическим и молекулярно-генетическим особенностям организмов, что значительно расширило объем данных, доступных для филогенетики. В то же время кладистика быстро стала популярной в эволюционной биологии, поскольку компьютеры позволили обрабатывать большие объемы данных об организмах и их характеристиках.
Методология
[ редактировать ]Этот раздел нуждается в дополнительных цитатах для проверки . ( Апрель 2016 г. ) |
Кладистический метод интерпретирует каждое преобразование состояния общего характера как потенциальное свидетельство группировки. Синапоморфии (общие производные состояния признаков) рассматриваются как свидетельство группировки, а симплезиоморфии (общие наследственные состояния признаков) - нет. Результатом кладистического анализа является кладограмма — древовидная диаграмма ( дендрограмма ). [ 18 ] это интерпретируется как лучшая гипотеза филогенетических взаимоотношений. Хотя традиционно такие кладограммы создавались в основном на основе морфологических признаков и первоначально рассчитывались вручную, данные генетического секвенирования и компьютерной филогенетики теперь широко используются в филогенетическом анализе, а экономности многие филогенетики отказались от критерия в пользу более «сложных» но менее экономные эволюционные модели трансформации состояний характера. Кладисты утверждают, что эти модели неоправданны, поскольку нет доказательств того, что они восстанавливают более «истинные» или «правильные» результаты из реальных наборов эмпирических данных. [ 19 ]
Каждая кладограмма основана на определенном наборе данных, анализируемом с помощью определенного метода. Наборы данных представляют собой таблицы, состоящие из молекулярных , морфологических, этологических [ 20 ] и/или другие признаки и список операционных таксономических единиц (OTU), которыми могут быть гены, особи, популяции, виды или более крупные таксоны, которые предположительно являются монофилетическими и, следовательно, образуют все вместе одну большую кладу; филогенетический анализ позволяет сделать вывод о характере ветвления внутри этой клады. Разные наборы данных и разные методы, не говоря уже о нарушениях упомянутых предположений, часто приводят к разным кладограммам. Только научное исследование может показать, что из них более вероятно.
До недавнего времени, например, кладограммы, подобные приведенным ниже, обычно считались точным представлением наследственных отношений между черепахами, ящерицами, крокодилами и птицами: [ 21 ]
▼
|
|
Если эта филогенетическая гипотеза верна, то последний общий предок черепах и птиц на ветке возле ▼ жил раньше, чем последний общий предок ящериц и птиц, недалеко от ♦ . большинство молекулярных данных дают кладограммы примерно такого вида: Однако [ 22 ]
Диапсида ♦
|
|
Если это верно, то последний общий предок черепах и птиц жил позже, чем последний общий предок ящериц и птиц. Поскольку кладограммы демонстрируют две взаимоисключающие гипотезы описания эволюционной истории, верна не более одной из них.
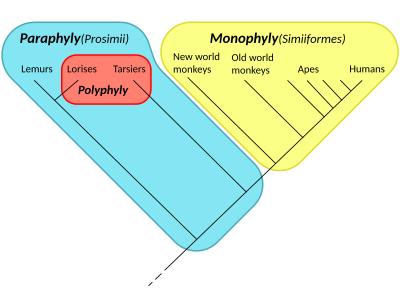
Кладограмма справа представляет текущую общепринятую гипотезу о том, что все приматы , включая стреппсирринов , таких как лемуры и лори , имели общего предка, все потомки которого являются или были приматами, и таким образом образуют кладу; поэтому для этой клады присвоено название «Приматы». Предполагается, что среди приматов все антропоиды (обезьяны, человекообразные обезьяны и люди) имели общего предка, все потомки которого являются или были антропоидами, поэтому они образуют кладу под названием Anthropoidea. С другой стороны, «просимиане» образуют парафилетический таксон. Название Prosimii не используется в филогенетической номенклатуре , в которой называются только клады; вместо этого «прозимианцы» разделены между кладами Strepsirhini и Haplorhini , где последняя содержит Tarsiiformes и Anthropoidea.
Лемуры и долгопяты, возможно, выглядели близкими родственниками человека в том смысле, что они были близки к человеку на эволюционном древе. Однако с точки зрения долгопята люди и лемуры выглядели бы одинаково близко. Кладистика придерживается нейтральной точки зрения, рассматривая все ветви (существующие или вымершие) одинаково. Это также вынуждает попытаться сделать заявления и честно принять во внимание выводы о точных исторических отношениях между группами.
Терминология состояний персонажа
[ редактировать ]Этот раздел нуждается в дополнительных цитатах для проверки . ( Апрель 2016 г. ) |
Следующие термины, придуманные Хеннигом, используются для определения общих или отдельных состояний характера среди групп: [ 23 ] [ 24 ] [ 25 ]
- Плезиоморфия — это состояние признака , («близкая форма») или предковое состояние которое таксон сохранил от своих предков. Когда два или более таксона, не вложенных друг в друга, имеют плезиоморфию, это симплезиоморфия ( от син- , «вместе»). Симплезиоморфии не означают, что таксоны, демонстрирующие этот признак, обязательно тесно связаны. Например, для рептилий традиционно характерно (помимо прочего) хладнокровие (т. е. не поддержание постоянной высокой температуры тела), тогда как птицы теплокровны . Поскольку хладнокровность является плезиоморфией, унаследованной от общего предка традиционных рептилий и птиц, и, следовательно, симплезиоморфией черепах, змей и крокодилов (среди прочих), это не означает, что черепахи, змеи и крокодилы образуют кладу, исключающую птицы.
- Апоморфия — («отдельная форма») или производное состояние это новация. Таким образом, его можно использовать для диагностики клады или даже для определения названия клады в филогенетической номенклатуре . Признаки, возникающие у отдельных таксонов (отдельного вида или группы, представленной одним терминалом в данном филогенетическом анализе), называются аутапоморфиями (от авто- , «сам»). Аутапоморфии ничего не говорят об отношениях между группами; клады идентифицируются (или определяются) синапоморфиями (от син- , «вместе»). Например, наличие пальцев , гомологичных пальцам человека разумного, является синапоморфией у позвоночных. Четвероногих вместе со всеми потомками этого позвоночного ( филогенетическое можно выделить как состоящих из первых позвоночных с такими пальцами, гомологичными пальцам Homo sapiens, основанное на апоморфии определение, ). [ 26 ] Важно отметить, что змеи и другие четвероногие, у которых нет пальцев, тем не менее, являются четвероногими: другие признаки, такие как амниотические яйца и диапсидные черепа, указывают на то, что они произошли от предков, которые обладали пальцами, гомологичными нашим.
- Состояние признака является гомопластичным или «примером гомоплазии », если оно является общим для двух или более организмов, но отсутствует у их общего предка или у более позднего предка в линии, ведущей к одному из организмов. Поэтому предполагается, что оно возникло путем конвергенции или разворота. И млекопитающие, и птицы способны поддерживать высокую постоянную температуру тела (т. е. они теплокровны). Однако принятая кладограмма, объясняющая их существенные особенности, указывает на то, что их общий предок находится в группе, лишенной этого состояния признака, поэтому государство должно было развиваться независимо в двух кладах. Теплокровность — это отдельно синапоморфия млекопитающих (или более крупной клады) и птиц (или более крупной клады), но она не является синапоморфией какой-либо группы, включающей обе эти клады. Вспомогательный принцип Хеннига [ 27 ] утверждает, что состояния общего характера следует рассматривать как свидетельство группировки, если только они не противоречат весу других доказательств; таким образом, гомоплазия некоторых признаков среди членов группы может быть сделана только после того, как будет установлена филогенетическая гипотеза для этой группы.
Термины плезиоморфия и апоморфия относительны; их применение зависит от положения группы внутри дерева. Например, при попытке решить, образуют ли четвероногие кладу, важным вопросом является то, является ли наличие четырех конечностей синапоморфией самых ранних таксонов, включенных в состав четвероногих: все ли самые ранние члены четвероногих унаследовали четыре конечности от общего предка? , тогда как все остальные позвоночные этого не сделали или, по крайней мере, не гомологично? Напротив, для группы четвероногих, например птиц, наличие четырех конечностей является плезиоморфией. Использование этих двух терминов обеспечивает большую точность при обсуждении гомологии, в частности, позволяя четко выражать иерархические отношения между различными гомологичными признаками.
Может быть трудно решить, является ли состояние признака на самом деле одинаковым и, таким образом, может быть классифицировано как синапоморфия, которая может идентифицировать монофилетическую группу, или оно только кажется одинаковым и, таким образом, является гомоплазией, которая не может идентифицировать такие группа. Существует опасность замкнутого круга рассуждений: предположения о форме филогенетического дерева используются для обоснования решений о состояниях признаков, которые затем используются в качестве доказательства формы дерева. [ 28 ] Филогенетика использует различные формы экономии для решения таких вопросов; сделанные выводы часто зависят от набора данных и методов. Такова природа эмпирической науки, и по этой причине большинство кладистов называют свои кладограммы гипотезами взаимоотношений. Кладограммы, которые поддерживаются большим количеством и разнообразием различных типов символов, считаются более надежными, чем те, которые основаны на более ограниченных доказательствах. [ 29 ]
Терминология таксонов
[ редактировать ]Моно-, пара- и полифилетические таксоны можно понимать по форме дерева (как это сделано выше), а также по состояниям их признаков. [ 24 ] [ 25 ] [ 30 ] Их сравнение приведено в таблице ниже.
| Срок | Определение на основе узла | Определение на основе символов |
|---|---|---|
| Голофилия, монофилия | Клада . , монофилетический таксон, — это таксон, состоящий из последнего общего предка и всех его потомков [ 31 ] | Клада характеризуется одной или несколькими апоморфиями : производными состояниями признаков, присутствующими у первого члена таксона, унаследованными его потомками (если только они не потеряны вторично) и не унаследованными никакими другими таксонами. |
| Парафилия | Парафилетический комплекс — это комплекс, который создается путем взятия клады и удаления одной или нескольких более мелких клад. [ 32 ] (Удаление одной клады дает одиночный парафилетический комплекс, удаление двух дает двойной парафилетический комплекс и так далее.) [ 33 ] | Парафилетический комплекс характеризуется одной или несколькими плезиоморфиями : состояниями признаков, унаследованными от предков, но присутствующими не у всех их потомков. Как следствие, парафилетический комплекс усекается, поскольку он исключает одну или несколько клад из монофилетического таксона. Альтернативное название — эволюционный уровень , относящееся к наследственному состоянию характера внутри группы. Хотя парафилетические комплексы популярны среди палеонтологов и эволюционных систематиков, кладисты не признают, что парафилетические ансамбли имеют какое-либо формальное информационное содержание — они являются всего лишь частями клад. |
| Полифилия | Полифилетическая совокупность – это совокупность, которая не является ни монофилетической, ни парафилетической. | Полифилетическая сборка характеризуется одной или несколькими гомоплазиями : состояниями признаков, которые слились или ревертировались, чтобы стать одинаковыми, но которые не были унаследованы от общего предка. Ни один систематик не признает полифилетические комплексы таксономически значимыми объектами, хотя экологи иногда считают их значимыми ярлыками для функциональных участников экологических сообществ (например, первичных продуцентов, детритофагов и т. д.). |
Критика
[ редактировать ]Кладистика, как в целом, так и в конкретных приложениях, с самого начала подвергалась критике. Решения о том, являются ли определенные состояния персонажей гомологичными , что является предварительным условием того, что они являются синапоморфиями, были оспорены как предполагающие циклические рассуждения и субъективные суждения. [ 34 ] Конечно, потенциальная ненадежность доказательств является проблемой для любого систематического метода или, если уж на то пошло, для любого эмпирического научного исследования вообще. [ 35 ] [ 36 ]
Трансформированная кладистика возникла в конце 1970-х годов. [ 37 ] в попытке решить некоторые из этих проблем путем удаления априорных предположений о филогении из кладистического анализа, но это осталось непопулярным. [ 38 ]
Проблемы
[ редактировать ]Предки
[ редактировать ]Кладистический метод не идентифицирует ископаемые виды как фактических предков клады. [ 39 ] Вместо этого ископаемые таксоны идентифицируются как принадлежащие к отдельным вымершим ветвям. Хотя ископаемый вид мог быть фактическим предком клады, узнать это невозможно. Следовательно, более консервативная гипотеза состоит в том, что ископаемый таксон связан с другими ископаемыми и существующими таксонами, о чем свидетельствует характер общих апоморфных особенностей. [ 40 ]
Статус исчезновения
[ редактировать ]Вымершая в противном случае группа, имеющая какие-либо сохранившиеся потомки, не считается (буквально) вымершей. [ 41 ] и, например, не имеет даты исчезновения.
Гибридизация, скрещивание
[ редактировать ]Все, что связано с биологией и сексом, сложно и запутанно, и кладистика не является исключением. [ 42 ] Многие виды размножаются половым путем и способны скрещиваться в течение миллионов лет. Хуже того, за такой период многие ветви могли разветвиться, и могут потребоваться сотни миллионов лет, чтобы они сократились до двух. [ 43 ] Только тогда теоретически можно определить правильных последних общих предков группировок, которые случайно не включают более ранние ветви. [ 44 ] Таким образом, процесс истинной кладистической бифуркации может занять гораздо более длительное время, чем обычно думают. [ 45 ] На практике, в отношении недавних излучений, кладистические данные дают лишь грубое представление о сложности. Более подробный отчет даст подробную информацию о долях интрогрессий между группировками и даже о их географических вариациях. Это использовалось в качестве аргумента в пользу использования парафилетических группировок. [ 44 ] но обычно приводятся другие причины.
Горизонтальный перенос генов
[ редактировать ]Горизонтальный перенос генов — это мобильность генетической информации между различными организмами, которая может иметь немедленные или отсроченные последствия для ответного хозяина. [ 46 ] В природе существует несколько процессов, которые могут вызвать горизонтальный перенос генов . Обычно это не влияет напрямую на происхождение организма, но может затруднить определение этого происхождения. На другом уровне можно составить карту процессов горизонтального переноса генов, определив филогению отдельных генов с помощью кладистики.
Стабильность именования
[ редактировать ]Если во взаимоотношениях есть неясность, то возможных деревьев очень много. Присваивать имена каждой возможной кладе может быть неразумно. Более того, устоявшиеся имена отбрасываются в кладистике или, альтернативно, несут коннотации, которые могут больше не сохраняться, например, когда в них обнаруживаются дополнительные группы. [ 47 ] Изменения в названиях являются прямым результатом изменений в признании взаимных отношений, которые часто все еще находятся в процессе изменения, особенно для вымерших видов. Сохранение старых имен и/или коннотаций контрпродуктивно, поскольку они обычно вообще не отражают в точности реальные взаимоотношения. Например, археи, асгардские археи, протисты, слизевики, черви, беспозвоночные, рыбы, рептилии, обезьяны, ардипитеки , австралопитеки , Homo erectus — все они содержат Homo sapiens кладистически, в их sensu lato , смысле . Для первоначально вымерших стволовых групп sensu lato обычно означает щедрое сохранение ранее включенных групп, которые затем могут включать даже живые виды. Вместо этого часто принимается значение «обрезанный в строгом смысле слова» , но группу необходимо ограничить одной ветвью на стебле. Остальные ветки получают свое имя и уровень. Это соизмеримо с тем фактом, что более старшие стволовые ветви фактически более близки к образующейся группе, чем более базальные стволовые ветви; то, что только эти стволовые ветви могли жить в течение короткого времени, не влияет на эту оценку кладистики.
По дисциплинам, кроме биологии
[ редактировать ]Сравнения, используемые для получения данных, на которых могут быть основаны кладограммы, не ограничиваются областью биологии. [ 48 ] Любую группу людей или классов, которые, как предполагается, имеют общего предка и к которым может применяться или не применяться набор общих характеристик, можно сравнивать попарно. Кладограммы можно использовать для изображения гипотетических отношений происхождения внутри групп предметов во многих различных научных областях. Единственное требование состоит в том, чтобы предметы имели характеристики, которые можно идентифицировать и измерить.
Антропология и археология : [ 49 ] Кладистические методы использовались для реконструкции развития культур или артефактов с использованием групп культурных черт или особенностей артефактов.
Сравнительная мифология и народные сказки используют кладистические методы для реконструкции протоверсий многих мифов. Мифологические филогении, построенные на основе мифов, явно подтверждают низкие горизонтальные передачи (заимствования), историческую (иногда палеолитическую) диффузию и прерывистую эволюцию. [ 50 ] Они также являются мощным способом проверить гипотезы о межкультурных отношениях между сказками. [ 51 ] [ 52 ]
Литература : Кладистические методы использовались при классификации сохранившихся рукописей Кентерберийских рассказов . [ 53 ] и рукописи санскритской Чарака Самхиты . [ 54 ]
Историческая лингвистика : [ 55 ] Кладистические методы использовались для реконструкции филогении языков с использованием лингвистических особенностей. Это похоже на традиционный сравнительный метод исторической лингвистики, но более явно использует экономию и позволяет гораздо быстрее анализировать большие наборы данных ( вычислительная филогенетика ).
Текстуальная критика или стемматика : [ 54 ] [ 56 ] Кладистические методы использовались для реконструкции филогении рукописей одной и той же работы (и реконструкции утерянного оригинала) с использованием характерных ошибок копирования в качестве апоморфий. Это отличается от традиционного историко-сравнительного языкознания тем, что позволяет редактору оценивать и устанавливать генетическую взаимосвязь больших групп рукописей с большим количеством вариантов, с которыми было бы невозможно справиться вручную. Это также позволяет экономно анализировать зараженные традиции передачи, которые невозможно оценить вручную за разумный период времени.
Астрофизика [ 57 ] делает выводы об истории взаимоотношений между галактиками для создания гипотез диверсификации галактик в виде диаграмм ветвления.
См. также
[ редактировать ]![]() Биологический портал
Биологический портал
![]() Портал эволюционной биологии
Портал эволюционной биологии
Примечания и ссылки
[ редактировать ]- ^ Харпер, Дуглас. «клада» . Интернет-словарь этимологии .
- ^ Колумбийская энциклопедия [ нужна полная цитата ]
- ^ «Введение в кладистику» . Ucmp.berkeley.edu . Проверено 6 января 2014 г.
- ^ Оксфордский словарь английского языка [ нужна полная цитата ]
- ^ Оксфордский словарь английского языка [ нужна полная цитата ]
- ^ Хикман, Кливленд П. младший (2014). Комплексные принципы зоологии (Шестнадцатое изд.). Нью-Йорк: Образование Макгроу-Хилл. ISBN 978-0-07-352421-4 . OCLC 846846729 .
- ^ «Необходимость кладистики» . www.ucmp.berkeley.edu . Проверено 12 августа 2018 г.
- ^ Бринкман и Лейпе 2001 , с. 323
- ^ Шу, Рэндалл. 2000. Биологическая систематика: принципы и приложения, стр.7 (цитата по Нельсону и Платнику, 1981). Издательство Корнельского университета (books.google)
- ^ Фолинсби, Кайла и др. 2007. 5 Количественных подходов к филогенетике, с. 172. Преподобный Мекс. Див. 225-52 (kfolinsb.public.iastate.edu)
- ^ Кроу, RC (1992). «Границы кладистики: тождество, различия и место в возникновении филогенетической систематики». В Гриффитсе, ЧП (ред.). Деревья жизни: Очерки философии биологии . Дордрехт: Клювер Академик. стр. 65–107. ISBN 978-94-015-8038-0 .
- ^ Шу, Рэндалл. 2000. Биологическая систематика: принципы и приложения, стр.7. Корнелл У. Пресс
- ^ Куэно 1940
- ^ Перейти обратно: а б Девятый новый университетский словарь Вебстера [ нужна полная цитата ]
- ^ Каин и Харрисон 1960
- ^ С 1984 г.
- ^ Лорен, Мишель (3 августа 2023 г.). Появление PhyloCode: продолжающаяся эволюция биологической номенклатуры . ЦРК Пресс. ISBN 978-1-000-91257-9 .
- ^ Вейгольд 1998
- ^ Риндал, Эйрик; Брауэр, Эндрю В.З. (2011), «Филогенетический анализ на основе моделей работает лучше, чем экономия? Тест с эмпирическими данными», Cladistics , 27 (3): 331–334, doi : 10.1111/j.1096-0031.2010.00342. х , PMID 34875779 , S2CID 84907350
- ^ Джерисон 2003 , с. 254
- ^ Бентон, Майкл Дж. (2005), Палеонтология позвоночных , Блэквелл, стр. 214, 233, ISBN 978-0-632-05637-8
- ^ Лайсон, Тайлер; Гилберт, Скотт Ф. (март – апрель 2009 г.), «Черепахи на всем пути вниз: болваны у корня хелонианского дерева» (PDF) , Evolution & Development , 11 (2): 133–135, CiteSeerX 10.1.1.695. 4249 , doi : 10.1111/j.1525-142X.2009.00325.x , PMID 19245543 , S2CID 3121166 , заархивировано (PDF) из оригинала 9 октября 2022 г.
- ^ Паттерсон 1982 , стр. 21–74.
- ^ Перейти обратно: а б Паттерсон 1988 г.
- ^ Перейти обратно: а б автор: Пинна, 1991 г.
- ^ Лорин и Андерсон, 2004 г.
- ^ Хенниг 1966
- ^ Джеймс и Пуртлесс IV 2009 , с. 25: «Синапоморфии используются для защиты гипотезы; гипотеза используется для защиты синапоморфий».
- ^ Брауэр, AVZ и Шу, RT. 2021. Биологическая систематика: принципы и приложения (3-е изд.). Издательство Корнеллского университета, Итака, штат Нью-Йорк
- ^ Паттерсон 1982
- ^ Подани, Янош (1 августа 2010 г.). «Монофилия и парафилия: дискурс без конца?» . Таксон . 59 (4): 1011–1015. дои : 10.1002/tax.594002 .
- ^ Многие источники дают словесное определение «парафилетичности», которое не требует, чтобы недостающие группы были монофилетическими. Однако когда представлены диаграммы, представляющие парафилетические группы, они неизменно показывают недостающие группы как монофилетические. См., например, Wiley et al. 1991 , с. 4
- ^ Тейлор 2003
- ^ Адрен, Эджкомб и Либерман, 2002 , стр. 56–57.
- ^ Орескес, Наоми , Кристин Шрейдер-Фрешетт и Кеннет Белитц. «Проверка, валидация и подтверждение численных моделей в науках о Земле». Наука 263, вып. 5147 (1994): 641-646.
- ^ Нильс Мёллер Андерсон, 2001 Влияние «филогенетической систематики» В. Хеннига на современную энтомологию Eur. Дж. Энтомол. 98:133-150 онлайн
- ^ Платник, Норман И. «Философия и трансформация кладистики». Систематическая зоология 28, вып. 4 (1979): 537–546.
- ^ Брауэр, Эндрю В.З. «Пятьдесят оттенков кладизма». Биология и философия 33, вып. 1-2 (2018): 8.
- ^ Крелл, Фрэнк-Т; Крэнстон, Питер С. (2004). «Какая сторона дерева более базальная?: Редакция» . Систематическая энтомология . 29 (3): 279–281. Бибкод : 2004SysEn..29..279K . дои : 10.1111/j.0307-6970.2004.00262.x . S2CID 82371239 .
- ^ Паттерсон, Колин. «Значение окаменелостей в определении эволюционных взаимоотношений». Ежегодный обзор экологии и систематики 12, вып. 1 (1981): 195–223.
- ^ Росс, Роберт М.; Олмон, Уоррен Д. (18 декабря 1990 г.). Причины эволюции: палеонтологическая перспектива . Издательство Чикагского университета. п. 133. ИСБН 978-0-226-72824-7 .
- ^ «Введение в кладистику» . ucmp.berkeley.edu . Проверено 8 мая 2022 г.
- ^ Харрисон, Ричард Г.; Ларсон, Эрика Л. (1 января 2014 г.). «Гибридизация, интрогрессия и природа границ видов» . Журнал наследственности . 105 (С1): 795–809. дои : 10.1093/jhered/esu033 . ISSN 0022-1503 . ПМИД 25149255 .
- ^ Перейти обратно: а б Хёрандль, Эльвира; Стюсси, Тод Ф. (2010). «Парафилетические группы как естественные единицы биологической классификации» . Таксон . 59 (6): 1641–1653. дои : 10.1002/tax.596001 . ISSN 0040-0262 . JSTOR 41059863 .
- ^ Мехта, Рохан С.; Розенберг, Ной А. (1 октября 2019 г.). «Вероятность реципрокной монофилии генных линий у трех и четырех видов» . Теоретическая популяционная биология . 129 : 133–147. дои : 10.1016/j.tpb.2018.04.004 . ПМК 6215533 . ПМИД 29729946 .
- ^ Эмамалипур, Мелисса; Сейди, Халед; Зунуни Вахед, Сепиде; Джаханбан-Исфахлан, Али; Джайманд, Мехди; Мажди, Хасан; Амузгар, Зохре; Читкушев, Л.Т.; Джавахери, Тахере; Джаханбан-Исфахлан, Рана; Заре, Пейман (2020). «Горизонтальный перенос генов: от эволюционной гибкости к прогрессированию заболевания» . Границы клеточной биологии и биологии развития . 8 : 229. дои : 10.3389/fcell.2020.00229 . ISSN 2296-634X . ПМК 7248198 . ПМИД 32509768 .
- ^ Дюбуа, Ален (1 августа 2007 г.). «Именование таксонов по кладограммам: некоторая путаница, вводящие в заблуждение утверждения и необходимые разъяснения» . Кладистика . 23 (4): 390–402. дои : 10.1111/j.1096-0031.2007.00151.x . ISSN 0748-3007 . ПМИД 34905840 . S2CID 59437223 .
- ^ Мейс, Клэр и Шеннан 2005 , стр. 1.
- ^ Липо и др. 2006 г.
- ^ д'Юи 2012а, б; д'Юи 2013а, б, в, г
- ^ Росс и др. 2013 год
- ^ Тегерани, 2013 г.
- ^ «Проект Кентерберийские рассказы» . Архивировано из оригинала 7 июля 2009 года . Проверено 4 июля 2009 г.
- ^ Перейти обратно: а б Маас 2010–2011 гг.
- ^ Оппенгеймер 2006 , стр. 290–300, 340–56.
- ^ Робинсон и О'Хара, 1996 г.
- ^ Fraix-Burnet et al. 2006 г.
Библиография
[ редактировать ]- Адрайн, Джонатан М.; Эджкомб, Грегори Д. и Либерман, Брюс С. (2002), Окаменелости, филогения и форма: аналитический подход , Нью-Йорк: Kluwer Academic, ISBN 978-0-306-46721-9 , получено 15 августа 2012 г.
- Барон, К. и Хёг, Дж.Т. (2005), «Гулд, Шарм и палеонтологическая перспектива в эволюционной биологии», в Кенеманне, С. и Дженнер, Р.А. (ред.), Взаимоотношения ракообразных и членистоногих , CRC Press, стр. 3 –14, ISBN 978-0-8493-3498-6 , получено 15 октября 2008 г.
- Бентон, MJ (2000), «Стебли, узлы, коронные клады и списки без рангов: Линней мертв?» (PDF) , Biological Reviews , 75 (4): 633–648, CiteSeerX 10.1.1.573.4518 , doi : 10.1111/j.1469-185X.2000.tb00055.x , PMID 11117201 , S2CID 17851383 , заархивировано из оригинал ( PDF) 9 августа 2017 г. , получено 2 октября 2011 г.
- Бентон, MJ (2004), Палеонтология позвоночных (3-е изд.), Оксфорд: Blackwell Science, ISBN 978-0-632-05637-8
- Бринкман, Фиона С.Л. и Лейпе, Детлеф Д. (2001), «Филогенетический анализ» (PDF) , в книге Баксеванис, Андреас Д. и Уэллетт, Б.Ф. Фрэнсис (ред.), Биоинформатика: практическое руководство по анализу генов и белков. (2-е изд.), стр. 323–358, заархивировано из оригинала (PDF) 20 октября 2013 г. , получено 19 октября 2013 г.
- Каин, Эй Джей; Харрисон, Джорджия (1960), «Филетическое взвешивание», Труды Лондонского зоологического общества , 35 : 1–31, doi : 10.1111/j.1469-7998.1960.tb05828.x
- Куэно, Люсьен (1940), «Замечания к эссе о генеалогическом древе животного мира», Comptes Rendus de l'Académie des Sciences de Paris , 210 : 23–27. Доступно бесплатно в Интернете на сайте Gallica (прямой URL-адрес отсутствует). Это статья, в которой Хенниг 1979 г. приписывает первое использование термина «клада».
- Дюпюи, Клод (1984), «Влияние Вилли Хеннига на таксономическую мысль», Annual Review of Ecology and Systematics , 15 : 1–24, doi : 10.1146/annurev.ecolsys.15.1.1 , ISSN 0066-4162 .
- Фаррис, Джеймс С. (1977), «О фенетическом подходе к классификации позвоночных», в Hecht, MK; Гуди, ПК; Хехт, Б.М. (ред.), Основные закономерности эволюции позвоночных , Пленум, Нью-Йорк, стр. 823–850.
- Фаррис, Джеймс С. (1979a), «О естественности филогенетической классификации», Systematic Zoology , 28 (2): 200–214, doi : 10.2307/2412523 , JSTOR 2412523
- Фаррис, Джеймс С. (1979b), «Информационное содержание филогенетической системы», Systematic Zoology , 28 (4): 483–519, doi : 10.2307/2412562 , JSTOR 2412562
- Фаррис, Джеймс С. (1980), «Эффективная диагностика филогенетической системы», Systematic Zoology , 29 (4): 386–401, doi : 10.2307/2992344 , JSTOR 2992344
- Фаррис, Джеймс С. (1983), «Логическая основа филогенетического анализа», Платник, Норман И.; Фанк, Вики А. (ред.), Достижения в кладистике, том. 2 , издательство Колумбийского университета, Нью-Йорк, стр. 7–36.
- Фрэкс-Берне, Д.; Холер, П.; Дузери, EJP; Верхамм, А. (2006), «Астрокладистика: филогенетический анализ эволюции галактик II. Формирование и разнообразие галактик», Journal of Classification , 23 (1): 57–78, arXiv : astro-ph/0602580 , Bibcode : 2006JClas ..23...57F , doi : 10.1007/s00357-006-0004-4 , S2CID 2537194
- Хенниг, Вилли (1966), Филогенетическая систематика (тр. Д. Дуайт Дэвис и Райнер Зангерл) , Урбана, Иллинойс: Univ. Illinois Press (переиздано в 1979 и 1999 гг.), ISBN 978-0-252-06814-0
- Хенниг, Вилли (1975), « Кладистический анализ или кладистическая классификация?»: ответ Эрнсту Майру (PDF) , Systematic Zoology , 24 (2): 244–256, doi : 10.2307/2412765 , JSTOR 2412765 , в архиве ( PDF) из оригинала от 9 октября 2022 г., в ответ на сообщение Майра 1974 г.
- Хенниг, Вилли (1999), Филогенетическая систематика (3-е издание книги 1966 г.) , Урбана: University of Illinois Press, ISBN 978-0-252-06814-0 Переведено с рукописи на немецкий язык, опубликованной в 1982 году (Phylogenetische Systematik, Verlag Paul Parey, Берлин).
- Халл, Дэвид (1988), Наука как процесс , University of Chicago Press, ISBN 978-0-226-36051-5
- Джеймс, Фрэнсис К. и Пуртлесс IV, Джон А. (2009), Кладистика и происхождение птиц: обзор и два новых анализа (PDF) , Орнитологические монографии, № 66, Союз американских орнитологов, ISBN 978-0-943610-85-6 , заархивировано (PDF) из оригинала 9 октября 2022 г. , получено 14 декабря 2010 г.
- д'Юи, Жюльен (2012a). «Медведь в звездах: филогенетическое исследование доисторического мифа» . Предыстория Юго-Запада . 20 (1): 91–106.
- д'Юи, Жюльен (2012b), «Мотив Пигмалиона: афразийское происхождение и распространение в Африке». Сахара , 23: 49-59 [1] .
- д'Юи, Жюльен (2013a), «Полифем (Aa. Th. 1137)». «Филогенетическая реконструкция доисторической сказки». Nouvelle Mythologie Comparée / Новая сравнительная мифология 1, [2]
- д'Юи, Жюльен (2013b). «Филогенетический подход мифологии и его археологические последствия» . Исследование наскального искусства . 30 (1): 115–118.
- д'Юи, Жюльен (2013c) «Мифы развиваются посредством знаков препинания». Французская мифология , 252, 2013c: 8-12. [3]
- д'Юи, Жюльен (2013d) «Космическая охота в берберском небе: филогенетическая реконструкция палеолитической мифологии». Les Cahiers de l'AARS , 15, 2013d: 93-106. [4]
- Джерисон, Гарри Дж. (2003), «О теории сравнительной психологии», Штернберг, Роберт Дж.; Кауфман, Джеймс К. (ред.), Эволюция интеллекта , Махва, Нью-Джерси: Lawrence Erlbaum Associates, Inc., ISBN 978-0-12-385250-2
- Лорин М. и Андерсон Дж. (2004), «Значение названия четвероногих в научной литературе: обмен» (PDF) , Systematic Biology , 53 (1): 68–80, doi : 10.1080/10635150490264716 , PMID 14965901 , заархивировано (PDF) из оригинала 9 октября 2022 г. [ мертвая ссылка ]
- Липо, Карл; О'Брайен, Майкл Дж.; Коллард, Марк; и др., ред. (2006), Картирование наших предков: филогенетические подходы в антропологии и предыстории , Пискатауэй: Transaction Publishers, ISBN 978-0-202-30751-0
- Маас, Филипп (2010–2011), Юрген, Ханнедер; Маас, Филипп (ред.), «Компьютерная стемматика - случай пятидесяти двух текстовых версий Carakasaṃhitā Vimānasthāna 8.67–157», Wiener Zeitschrift für die Kunde Südasias , 52–53: 63–120, doi : 10.1553/wzks2009- 2010с63
- Мейс, Рут; Клэр, Клэр Дж.; Шеннан, Стивен, ред. (2005), Эволюция культурного разнообразия: филогенетический подход , Портленд: Cavendish Press, ISBN 978-1-84472-099-6
- Майр, Эрнст (1974), «Кладистический анализ или кладистическая классификация?» (PDF) , Журнал зоологической систематики и эволюционных исследований , 12 : 94–128, doi : 10.1111/j.1439-0469.1974.tb00160.x , заархивировано (PDF) из оригинала 9 октября 2022 г.
- Майр, Эрнст (1976), Эволюция и разнообразие жизни (Избранные эссе) , Кембридж, Массачусетс: Издательство Гарвардского университета, ISBN 978-0-674-27105-0 Переиздано в мягкой обложке в 1997 году. Включает перепечатку статьи Майра по антикладистике 1974 года, стр. 433–476, «Кладистический анализ или кладистическая классификация». Это статья, на которую Хенниг 1975 года является ответом.
- Майр, Эрнст (1978), «Происхождение и история некоторых терминов в систематической и эволюционной биологии», Systematic Zoology , 27 (1): 83–88, doi : 10.2307/2412818 , JSTOR 2412818 .
- Майр, Эрнст (1982), Рост биологической мысли: разнообразие, эволюция и наследование , Кембридж, Массачусетс: издательство Гарвардского университета, ISBN 978-0-674-36446-2
- Оппенгеймер, Стивен (2006), Происхождение британцев , Лондон: Робинсон, ISBN 978-0-7867-1890-0
- Паттерсон, Колин (1982), «Морфологические признаки и гомология», в Джойси, Кеннет А.; Пятница, AE (ред.), Проблемы филогенетической реконструкции , специальный том 21 Ассоциации систематики, Лондон: Academic Press, ISBN 978-0-12-391250-3 .
- Паттерсон, Колин (1988), «Гомология в классической и молекулярной биологии», Molecular Biology and Evolution , 5 (6): 603–625, doi : 10.1093/oxfordjournals.molbev.a040523 , PMID 3065587
- де Пинна, MGG (1991), «Концепции и тесты гомологии в кладистической парадигме» (PDF) , Cladistics , 7 (4): 367–394, CiteSeerX 10.1.1.487.2259 , doi : 10.1111/j.1096-0031.1991 .tb00045.x , S2CID 3551391 , заархивировано из оригинала (PDF) 22 июля 2011 г. , получено 24 октября 2017 г.
- де Кейроз, К. и Готье, Дж. (1992), «Филогенетическая таксономия» (PDF) , Annual Review of Ecology and Systematics , 23 : 449–480, doi : 10.1146/annurev.ecolsys.23.1.449 , заархивировано из оригинал (PDF) 20 марта 2012 г. , дата обращения 28 июля 2012 г.
- Робинсон, Питер М.В. и О'Хара, Роберт Дж. (1996), «Кладистический анализ древнескандинавской рукописной традиции» , Research in Humanities Computing , 4 : 115–137 , получено 13 декабря 2010 г.
- Росс, Роберт М.; Гринхилл, Саймон Дж.; Аткинсон, Квентин Д. (2013). «Структура населения и культурная география сказки в Европе» . Труды Королевского общества B: Биологические науки . 280 (1756): 20123065. doi : 10.1098/rspb.2012.3065 . ПМЦ 3574383 . ПМИД 23390109 .
- Шу, Рэндалл Т. и Брауэр, Эндрю В.З. (2009), Биологическая систематика: принципы и приложения (2-е изд.), Cornell University Press, ISBN 978-0-8014-4799-0
- Тейлор, Майк (2003), Что означают такие термины, как монофилетический, парафилетический и полифилетический? , получено 13 декабря 2010 г.
- Техрани, Джамшид Дж., 2013, «Филогения Красной Шапочки», PLOS ONE , 13 ноября. [5]
- Трембле, Фредерик (2013), «Николай Хартманн и метафизическая основа филогенетической систематики» , Biological Theory , 7 (1): 56–68, doi : 10.1007/s13752-012-0077-8 , S2CID 84932063
- Вейгольдт, П. (февраль 1998 г.), «Эволюция и систематика Chelicerata», Experimental and Applied Acarology , 22 (2): 63–79, doi : 10.1023/A:1006037525704 , S2CID 35595726
- Уилер, Квентин (2000), Видовые концепции и филогенетическая теория: дебаты , Columbia University Press, ISBN 978-0-231-10143-1
- Уайли, Э.О.; Сигел-Кози, Д.; Брукс, Д.Р. и Фанк, Вирджиния (1991), «Глава 1. Введение, термины и концепции», The Compleat Cladist: Primary of Philogenetic Processes (PDF) , Музей естественной истории Канзасского университета, ISBN. 978-0-89338-035-9 , заархивировано из оригинала (PDF) 3 декабря 2010 г. , получено 13 декабря 2010 г.
- Уильямс, Пенсильвания (1992), «Путаница в кладизме», Synthese , 01 (1–2): 135–152, doi : 10.1007/BF00484973 , S2CID 46974048
Внешние ссылки
[ редактировать ] СМИ, связанные с кладистикой, на Викискладе?
СМИ, связанные с кладистикой, на Викискладе? - OneZoom: Tree of Life — все живые виды в виде интуитивно понятного и масштабируемого исследователя фракталов (адаптивный дизайн)
- Общество Вилли Хеннига
- Кладистика (научный журнал Общества Вилли Хеннига)
- Коллинз, Аллен Г.; Гуральник, Роб; Смит, Дэйв (1994–2005). «Путешествие в филогенетическую систематику» . Музей палеонтологии Калифорнийского университета . Проверено 21 января 2010 г.
- Фельзенштейн, Джо. «Программы филогении» . Сиэтл: Вашингтонский университет . Проверено 21 января 2010 г.
- О'Нил, Деннис (1998–2008). «Классификация живых существ» . Сан-Маркос, Калифорния: Колледж Паломар. Архивировано из оригинала 11 января 2010 года . Проверено 21 января 2010 г.
- Робинсон, Питер; О'Хара, Роберт Дж. (1992). «Отчет о проблеме текстовой критики 1991 года» . rjohara.net . Проверено 21 января 2010 г.
- Теобальд, Дуглас (1999–2004). «Букварь по филогенетике» . Архив TalkOrigins . Проверено 21 января 2010 г.
| Соответствующие поля | ||
|---|---|---|
| Основные понятия | ||
| Методы вывода | ||
| Текущие темы | ||
| Групповые черты | ||
| Типы групп | ||
| Номенклатура | ||