Уральская сова
| Уральская сова | |
|---|---|

| |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Птицы |
| Заказ: | Стригиформы |
| Семья: | Стригиды |
| Род: | Стрикс |
| Разновидность: | С. ураленсис
|
| Биномиальное имя | |
| Стрикс уральский Паллада , 1771 г.
| |
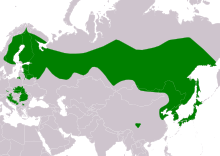
| |
| Ареал S. uralensis Резидент
| |

Уральская неясыть ( Strix uralensis ) — крупная ночная сова . Это член настоящего семейства сов , Strigidae . Уральская сова является представителем рода Strix , от которого также произошло название семейства в таксономии Линнея . [ 3 ] И его общее название, и научное название относятся к Уральским горам России , где типовой экземпляр был собран . Однако этот вид имеет чрезвычайно широкое распространение, которое простирается на запад до большей части Скандинавии , горной Восточной Европы и, спорадически, Центральной Европы , а затем простирается через Палеарктику через Россию до дальнего востока до Сахалина и всей Японии . [ 1 ] [ 4 ] Уральская неясыть может включать до 15 подвидов, но, скорее всего, их число может быть несколько меньше, если учитывать клинальные вариации . [ 5 ]
Эта лесная сова типична для обширных таежных лесов Евросибири, хотя встречается и в других типах лесов, включая смешанные леса и лиственные леса умеренного пояса . [ 5 ] [ 6 ] Уральская сова является своего рода универсальным диетологом, как и многие представители рода Strix , но обычно на местном уровне она питается мелкими млекопитающими , особенно мелкими грызунами , такими как полевки . [ 4 ] [ 7 ] Что касается репродуктивных привычек, уральские совы, как правило, энергично защищают определенную территорию, на которой они исторически гнездились в различных естественных местах гнездования, включая дупла деревьев и пни, а также гнезда, первоначально построенные другими птицами , но теперь во многих частях Ассортимент адаптирован к гнездовым ящикам, сделанным биологами и защитниками природы . [ 8 ] [ 9 ] Успех размножения часто сильно коррелирует с популяцией добычи . [ 10 ] Уральская неясыть в целом считается стабильным видом птиц, имеющим природоохранный статус согласно МСОП как вид, вызывающий наименьшее беспокойство . [ 1 ] Несмотря на некоторое локальное сокращение и исчезновение помогла уральской совы в Центральной Европе , реинтродукция ей . [ 11 ]
Описание
[ редактировать ]Как и у большинства видов Strix , у него широкая округлая голова с соответствующим круглым лицевым диском, за исключением крошечного V-образного углубления. У уральской совы исключительно длинный для совы хвост с клиновидным кончиком. По цвету он, как правило, простой от бледно-серовато-коричневого до беловатого в целом (с более подробным описанием их вариаций в зависимости от подвида), с немного более темной серо-коричневой или коричневой спиной и спиной с контрастными беловатыми отметинами. Нижняя часть тела от бледно-кремово-охристого до серо-коричневого цвета, ярко (хотя иногда и более тонко) покрыта темно-коричневыми полосами, без перекладин. Известно множество вариаций общей окраски оперения как на уровне подвида, так и на индивидуальном уровне. Однако уральская сова обычно выглядит как довольно бледная серо-коричневая сова, которой обычно не хватает более теплых и насыщенных цветовых тонов, как у многих других сов Стрикс , с отчетливыми полосами внизу. [ 4 ] [ 5 ] [ 8 ] В полете у уральской совы видно преимущественно охристо-белое нижнее крыло с тяжелыми темными полосами вокруг заднего края и кончика, а длинный хвост с белым кончиком часто кажется опущенным вниз. [ 8 ] Их стиль полета напоминает канюка , но с более глубокими и расслабленными взмахами крыльев, а их стиль полета часто создает впечатление довольно большой птицы. [ 8 ] Глаза темно-карие, относительно маленькие и близко посажены друг к другу, что, как полагают, придает им менее «свирепый» вид, чем у большой серой совы ( Strix nebulosa ). [ 5 ] [ 8 ] напоминают миндаль . Глаза по форме и цвету [ 4 ] Клюв . желтоватого цвета, с грязно-желтой восковицей Между тем, лапки и пальцы ног покрыты сероватым оперением, а когти желтовато-коричневые с более темными кончиками. [ 4 ] [ 5 ]
Уральская неясыть – довольно крупный вид. Общая длина взрослых особей составляет от 50 до 64 см (от 20 до 25 дюймов), что может сделать их примерно восьмыми самыми длинными видами сов в мире (хотя многие совы в среднем тяжелее). [ 5 ] [ 12 ] [ 13 ] [ 14 ] Размах крыльев у разных видов может варьироваться от 110 до 134 см (от 3 футов 7 дюймов до 4 футов 5 дюймов). [ 4 ] [ 12 ] Как и большинство хищных птиц , уральская неясыть демонстрирует обратный половой диморфизм в размерах: самка в среднем немного крупнее самца. [ 5 ] Сообщается, что размер когтей и масса тела — лучший способ отличить два пола уральской совы, кроме поведенческой дихотомии, основанной на наблюдениях в Финляндии . [ 15 ] Вес варьируется в европейской части ассортимента. Известно, что самцы весят от 451 до 1050 г (от 0,994 до 2,315 фунта), а самки - от 569 до 1454 г (от 1,254 до 3,206 фунта). [ 16 ] [ 17 ] Воус оценил типичный вес самцов и самок в 720 г (1,59 фунта) и 870 г (1,92 фунта) соответственно. [ 4 ] Это один из самых крупных видов рода Strix , который в целом примерно на 25% меньше большой серой совы , причем последняя, безусловно, является самым крупным из существующих видов Strix по всем методам измерения. Масса тела, зарегистрированная для некоторых более южных азиатских видов, таких как коричневая лесная сова ( Strix leptogrammica ) и пятнистая лесная сова ( Strix selopato ) (а также аналогичного размера, но не взвешенная пестрая лесная сова ( Strix ocellata )) показывает, что они в значительной степени перекрываются. по массе тела с уральской совой или даже несколько тяжелее, как правило, несмотря на то, что они несколько меньше в длине, несколько коренастые по телосложению, но короче хвост, чем уральская неясыть. [ 16 ] [ 18 ] Несмотря на отсутствие опубликованных данных о весе взрослых особей, сова Пер-Давида ( Strix davidi ), по-видимому, также имеет такой же размер, что и уральская сова. [ 19 ] Среди стандартных размеров у обоих полов хорда крыла может составлять от 267 до 400 мм (от 10,5 до 15,7 дюйма) по всему диапазону, а длина хвоста - от 201 до 320 мм (от 7,9 до 12,6 дюйма). Среди ныне живущих сов только у большой серой совы наверняка более длинный хвост. Хотя измерения проводятся реже, длина предплюсны может составлять от 44 до 58,5 мм (от 1,73 до 2,30 дюйма), а в Северной Европе общая длина клюва составляет от 38 до 45 мм (от 1,5 до 1,8 дюйма). [ 5 ] [ 17 ] [ 20 ] [ 21 ] У взрослых сов размах стопы обычно может достигать около 14,3 см (5,6 дюйма). [ 15 ]
Вокализации и морфология ушей
[ редактировать ]Песня самца представляет собой глубокую ритмичную серию нот с небольшой паузой после первых двух нот, транскрибируемую по-разному как виху хуу-хухуво или хуу-хуу-хуу . Фраза повторяется с интервалом в несколько секунд. [ 5 ] Песня самца может разноситься на расстояние до 2 км (1,2 мили) для человеческого восприятия, но обычно считается, что она не так уж и далеко распространяется. [ 22 ] Пиковое время пения в Финляндии весной приходится на 22:00–12:00 и более интенсивное в 1–3 часа ночи, что отличается от пикового времени посещения гнезд. [ 23 ] У самки похожая песня, но более хриплая и немного более высокая, что придает ей более «лающий» характер. [ 5 ] Нередко уральские совы образуют дуэт во время ухаживания. [ 5 ] глубокая, хриплая цапля типа кувата или коррвика . Кроме того, в качестве контактного призыва, вероятно, используется [ 5 ] [ 8 ] Они более удлиненные и резкие, чем ноты Кьюика, издаваемые желтовато-коричневыми совами ( Strix aluco ). [ 5 ] [ 8 ] Молодые просят милостыню хриплыми криками «хррре» , опять же похожими на крики молодых желтовато-коричневых сов, но более глубокие. [ 4 ] [ 5 ] [ 8 ] Вокальное поведение имеет тенденцию достигать пика ранней весной, пока птенцы не покинут гнездо, чаще всего во время инкубации и гнездования в форме встречного вызова добычи. [ 22 ] Тревожный сигнал самца, который обычно издается во время территориальных обходов, по совпадению аналогичен территориальной песне ушастой совы ( Asio flammeus ), которая считается несколько глухим уханьем. Сигнал тревоги слышен на расстоянии до 1500 м (4900 футов). Уральская неясыть также обладает особенно угрожающим поведением, когда она щелкает купюрами. [ 22 ] [ 24 ] Всего шведский биолог сообщил, что уральские совы издавали около девяти различных криков. [ 22 ] [ 25 ] [ 26 ] [ 27 ] Несмотря на диапазон криков, этот вид, как правило, очень тихий для большой совы и может не издавать вокала даже в часы пик в течение почти двух дней. [ 8 ]
Уши уральской совы довольно большие: в среднем около 24 мм (0,94 дюйма) слева и 27 мм (1,1 дюйма) справа, а размер предушного кожного лоскута составляет около 13 мм (0,51 дюйма). На самом деле их уши являются одними из самых больших среди сов. [ 4 ] [ 28 ] [ 29 ] В сочетании с большими ушами хорошо развитый лицевой диск показывает важность звука для охоты для этой и других сов, которые охотятся в бореальных зонах . Хотя уральская неясыть оказалась слишком развитой по сравнению с другими стриксами , такими как полосатая неясыть ( Strix varia ), она оказалась недоразвитой по сравнению с совами, более привязанными к ареалам истинного бореального типа, такими как большая серая неясыть и бореальная неясыть. ( Эголиус Фунереус ). [ 28 ] [ 29 ] [ 30 ]
Путаница с похожими видами сов.
[ редактировать ]Уральская сова — птица довольно своеобразного вида, но ее можно спутать с другими совами, особенно с другими совами рода Strix . [ 5 ] [ 31 ] Экстралимитная североамериканская полосатая неясыть выглядит очень похожей, основной отличительной чертой этих двух видов является отсутствие у уральской неясыни концентрических линий на лицевой пластине. Желто -коричневая сова намного меньше, у нее заметно более короткий хвост и относительно большая голова. Желтовато-коричневый вид, который встречается в серой, коричневой и красной морфах, имеет нижнюю часть тела с темными полосами на стержне и перекладинами, в отличие от тяжелых, но прямых полосатости уральской совы. [ 5 ] [ 30 ] Большая серая неясыть крупнее уральской совы, с огромной головой и относительно еще меньшими желтыми глазами, а их лицевой диск имеет четкие концентрические линии . По цвету большая серая неясыть заметно более однородная и несколько темно-серая, чем уральская неясыть. [ 5 ] [ 30 ] [ 31 ] Маловероятно, что уральскую сову можно принять за уральскую сову — это евразийский филин ( Bubo bubo ), который намного крупнее (значительно самая тяжелая и длинная крылатая сова в Европе) с выступающими пучками ушей, квадратной (не закругленной) формой головы и оранжевые глаза, а также с совершенно разными отметинами. [ 5 ] [ 31 ] Ушастые совы ( Asio otus ) намного меньше и стройнее, с выступающими кисточками на ушах, оранжевыми глазами и более заметными темными отметинами. [ 5 ] Близкородственная сова Пер-Давида , более похожая на любую другую в Европе, не встречается в том же ареале, что и (другие?) Уральские совы, но имеет более темное оперение, а также лицевой диск, отмеченный более темными концентрическими линиями. [ 5 ] [ 19 ] Из-за его частично дневного поведения в теплые месяцы некоторые авторы считают, что его можно спутать с совершенно другим на вид (но таким же крупным и длиннохвостым) евразийским ястребом-тетеревятником ( Accipiter gentilis ). [ 4 ] [ 32 ]
Таксономия
[ редактировать ]
Уральская сова была названа Питером Саймоном Палласом в 1771 году как Strix uralensis из-за того, что типовой экземпляр был собран в Уральских горах . Хотя Урал находится примерно в середине распространения этого вида, некоторые авторы, такие как Карел Вуз, не было получено более широко подходящее, чем Уральская сова сетовали на то, что для общего английского названия . [ 4 ] На других языках вид упоминается как Slaguggla , или «атакующая сова», по- Habichtskauz шведски , или « ястреб-тетеревятник », по- немецки или как «длиннохвостая сова» по-русски. [ 4 ] [ 11 ] [ 32 ] [ 33 ] Уральская неясыть — представитель рода Strix , которого довольно часто называют лесными совами. [ 13 ] По консервативным оценкам, в этом роде в настоящее время представлено около 18 видов, обычно это совы среднего и крупного размера, с характерной круглой головой и отсутствием пучков ушей, которые приспосабливаются к жизни в лесных частях различных климатических зон. [ 18 ] [ 34 ] Четыре совы, обитающие в неотропах, иногда дополнительно включаются в род Strix , но некоторые авторитеты также включили их в отдельный, но родственный род Ciccaba . [ 35 ] [ 36 ] Совы Стрикс имеют обширную летопись окаменелостей и уже давно широко распространены. [ 37 ] Генетическое родство настоящих сов несколько запутано, и различные генетические тесты по-разному показали, что совы Стрикс связаны с разнородными родами, такими как Pulsatrix , Bubo и Asio . [ 5 ] [ 17 ] [ 36 ] [ 38 ]
Считается, что серая неясыть является близким родственником уральской совы. Авторы выдвинули гипотезу, что происхождение разделения видов последовало за плейстоцена континентальными оледенениями , которые отделили юго-западную или южную группу в лесах умеренного пояса (т.е. желтовато-коричневые) от восточной группы, населяющей холодные бореальные ареалы (т.е. Урал). Видовой образец отражается у других видов птиц, например, у европейского зеленого дятла ( Picus viridus ) и у более северного трансконтинентального седого дятла ( Picus canus ). После отступления континентальных ледяных масс хребты в последнее время проникли друг в друга. [ 4 ] [ 29 ] [ 39 ] Хотя детали истории жизни коричневой и уральской совы во многом совпадают, тем не менее виды имеют ряд морфологических различий и в значительной степени адаптированы к разным климатическим условиям , времени активности и местам обитания. [ 4 ] [ 27 ] На основе Strix ископаемых видов из среднего плейстоцена (получивших название Strix intermedia ) в Чехии , Австрии и Венгрии , кости ног и крыльев указывают на животное промежуточной формы и размера между уральскими и коричневыми совами. [ 40 ] [ 41 ] [ 42 ] [ 43 ] Тем не менее, окаменелости более крупной и пропорциональной совы Strix , отличающейся от желто-коричневой совы, идентифицированной как Strix brevis , из Германии и Венгрии до плейстоцена (т.е. пиаченского периода ), а также диагностированные окаменелости уральской совы из разрозненных южных отложений на Сардинии из раннего Плейстоцен и отложения среднего плейстоцена в Паннонском бассейне , а также намного позже, в раннем голоцене , далеко на западе в Бельгии , Франции. и Швейцария предполагают более сложную историю эволюции и распространения. [ 4 ] [ 41 ] [ 42 ] [ 43 ] [ 44 ] [ 45 ] В неволе был зарегистрирован гибрид самца уральской совы и самки коричневой совы, которому удалось произвести на свет двух потомков промежуточного размера и иметь более сложную песню, которая также имела некоторые общие характеристики с вокализацией обоих видов. [ 46 ]
Некоторые виды в Америке, такие как полосатая неясыть , иногда считаются настолько близкими родственниками, что уральская и полосатая и пятнистая совы ( Strix occidentalis ) потенциально считаются частью комплекса видов. или даже внутри одного вида. [ 47 ] Однако нет никаких доказательств и вероятности того, что совы Стрикс между Америкой и Евразией когда-либо образовывали непрерывную популяцию, учитывая их адаптацию к хорошо засаженным деревьями территориям, а также тот факт, что полосатая неясыть более экологически похожа на более распространенную коричневую неясыть, несмотря на имеют промежуточный размер между желто-коричневой совой и уральской совой (ближе по размеру к последней), и что желто-коричневая не обитает где-либо близко к границе между Северной Америкой и Россией, как уральская. [ 4 ] [ 48 ] [ 49 ] [ 50 ] Конечно, самым неоднозначным аспектом взаимоотношений уральской совы является сова Пер-Давида , которая как исторически, так и в настоящее время считается либо изолированным подвидом уральской совы, либо отдельным видом. Считается, что Пер-Давид, вероятно, является ледниковым реликтом горных лесов западного Китая , где растительный и животный мир часто напоминает доледниковую жизнь. [ 4 ] [ 29 ] [ 51 ] Недавнее исследование показало, что сова Пер-Давида является действительным видом на основании различий во внешности, голосе и истории жизни, хотя генетические исследования показали несколько запутанное разнообразие между расами комплекса видов уральских сов. [ 19 ] [ 52 ] Он был признан Контрольным списком птиц мира Клементса. [ 53 ] но BirdLife International [ 54 ] и МСОП [ 55 ] до сих пор относят ее к подвиду уральской совы.
Подвиды
[ редактировать ]
Некоторые авторитеты признали по крайней мере 15 подвидов, хотя некоторые считают, что действительных подвидов может быть всего лишь восемь. [ 5 ] [ 17 ] [ 20 ] Изучение филогеографической генетики уральских сов в Европе показало, что они встречаются примерно в пяти различных генетических кластерах, которые не соответствуют подвидам и не соответствуют им, как ожидалось, и, кроме того, генетический обмен значителен там, где ареалы подвидов граничат. Филогеографические данные указывают на непрерывную популяцию в не слишком далекой истории и дискредитируют общую классификацию вида как ледникового реликта , причем изоляция популяций, вероятно, во многом связана с вмешательством человека. [ 56 ]
- С. у. uralensis ( Pallas , 1771). Это номинальный подвид, который, как полагают, встречается во внутренней части Дальнего Востока России , от Сибири до Якутии и Охотского побережья к югу от Средней Волги , южного Урала , Тюмени и Ялуторовского . самцов Хорда крыла составляет от 334 до 375 мм (от 13,1 до 14,8 дюйма), а у самок - от 348 до 380 мм (от 13,7 до 15,0 дюйма). Небольшая выборка самцов весила от 500 до 712 г (от 1,102 до 1,570 фунта), а одна самка весила 950 г (2,09 фунта). [ 5 ] [ 17 ] В номинальной расе встречаются светлые и темные морфы, причем первые гораздо более многочисленны. Этот подвид, вероятно, является самой бледной формой уральской совы, причем некоторые сибирские совы настолько беловатые и со слабыми пятнами, что на мгновение напоминают полярную сову ( Bubo scandianus ). [ 8 ] Типичная светлая морфа имеет лицевой диск от грязно-беловатого до бледно-охристо-серого цвета, а кайма вокруг диска состоит из рядов мелких темных пятен. Верхняя часть тела бледно-серовато-коричневая, пестрая, пятнистая и с различными прожилками, беловатыми и темными, а лопатки имеют довольно большую площадь белого цвета. Горло беловатое, а остальная нижняя сторона от очень бледного серовато-коричневого до грязно-беловатого цвета с густыми, но не темно-коричневыми пестринами. Лапки и пальцы ног окрашены в цвет от бледно-серовато-коричневого до грязно-беловато-кремового. [ 5 ] [ 32 ]
- С. у. Давиди ( Шарп , 1875) — сова Папа Давида. Встречается в горах центрального Китая ( Цинхай и Сычуань ). Ведутся споры о том, является ли это отдельным видом или изолированным подвидом уральской совы.
- С. у. литурата ( Tengmalm , 1795) — эта раса встречается в северной Европе от восточной Норвегии , Лапландии и других мест Швеции , по всему ареалу в Финляндии , до Балтийского региона (в основном северной Белоруссии ), восточной Польши , восточных Альп вниз, самых восточных Карпаты , к востоку от Волги . Птицы этой расы иногда смешиваются в восточной части ареала с номинальным подвидом. Хорда крыла самцов составляет от 342 до 368 мм (от 13,5 до 14,5 дюймов), а у самок - от 349 до 382 мм (от 13,7 до 15,0 дюймов). Длина хвоста составляет от 253 до 282 мм (от 10,0 до 11,1 дюйма). Сообщается, что известная масса тела у мужчин составляет от 451 до 900 г (от 0,994 до 1,984 фунта). [ 5 ] [ 17 ] [ 30 ] Несмотря на то, что он линейно меньше, чем S. u. marcoura , большая выборка самок уральских сов в Финляндии, показала, что они были на удивление примерно на 16% тяжелее, чем особи в Румынии: 542 особи весили в среднем 1031 г (2,273 фунта) с диапазоном от 785 до 1350 г (от 1,731 до 2,976 фунта). [ 57 ] Однако некоторые самки этого подвида могут весить всего 520 г (1,15 фунта) и, в отличие от вида уральской совы в целом, могут демонстрировать некоторый уровень соответствия правилу Бергмана . [ 17 ] Эта раса внешне очень похожа на номинальную форму, но в среднем немного темнее, с более насыщенными и темными коричневыми полосами внизу у большинства птиц, которые доходят до нижнего края лицевого диска, и обычно менее обширными белыми пятнами и пестринами на спине и спине. мантия. [ 5 ] [ 17 ]

- С. у. macroura ( Wolf , 1810) — эта раса, близкая к S. u., но обычно встречающаяся в других областях. Liturata произрастает в западных Карпатах , Трансильванских Альпах и на юге Западных Балкан . У обоих полов хорда крыла может составлять от 354 до 415 мм (от 13,9 до 16,3 дюйма). Длина хвоста составляет от 282 до 315 мм (от 11,1 до 12,4 дюйма). Если судить по линейным измерениям, это самая крупная по размеру раса уральской совы, несмотря на то, что она имеет самый южный ареал среди всех рас в Европе. [ 5 ] [ 17 ] [ 30 ] В Румынии 40 мужчин весили в среднем 706 г (1,556 фунта) с диапазоном от 503 до 950 г (от 1,109 до 2,094 фунта), а 57 женщин весили в среднем 863 г (1,903 фунта) с диапазоном 569 г. до 1307 г (от 1,254 до 2,881 фунта). [ 16 ] Однако для этой расы известны как отдельные самцы, так и самки, которые были тяжелее самых крупных в приведенной выше выборке. [ 17 ] Вероятно, это самая темная окраска из рас уральских сов. Типичная взрослая особь покрыта коричневатым охристым налетом на лицевом диске и нижней стороне, который также имеет очень широкие черноватые полосы. Между тем, у типичной птицы спина темно-серо-коричневая, из-за чего белые части крыльев контрастируют. [ 5 ] Темные морфы также нередки. В выборке 522 С. е. macroura , 11,7% составляли темные морфы, тогда как ранее считалось, что только около 6% были темными морфами. На местном уровне даже до 15% птиц могут быть темными морфами. Темные морфы в целом имеют насыщенный темно-шоколадно-коричневый цвет, особенно на лицевом диске. Мантия темных морф темно-кофейно-коричневая со слегка отличающимися серо-коричневыми пестринами, без белых пятен, наблюдаемых в этой области у других уральских сов. На нижней стороне обычно видны темно-шоколадно-коричневые полосы поверх средне-коричневого основного цвета. [ 4 ] [ 58 ]
- С. у. yenisseensis ( Бутурлин , 1915). Эта раса встречается от Среднесибирского плоскогорья до Забайкалья и северо-западной Монголии, но в двух последних районах они в основном регистрируются как зимние залетные. самцов Хорда крыла составляет от 328 до 350 мм (от 12,9 до 13,8 дюйма), а у самок - от 348 до 370 мм (от 13,7 до 14,6 дюйма). Длина хвоста от 235 до 282 мм (от 9,3 до 11,1 дюйма). Эта раса очень похожа на номинальный подвид, но, как правило, в целом немного менее беловатая и, как правило, намного меньше, чем она, а также большинство европейских уральских сов. [ 17 ] [ 20 ] [ 21 ] Известно, что встречаются гибриды между этим и номинальным подвидом. [ 17 ] [ 20 ]
- С. у. Никольский ( Бутурлин , 1907) — по мнению некоторых авторов, к этой расе относятся ранее признанные подвиды S. u. даурика , С.у. тайбанай и С.у. корренсис . [ 5 ] Эта раса встречается в Забайкалье к северу и востоку от Витима , Сахалина и к югу от Корейского полуострова . Эта раса довольно мала: она меньше всех рас к западу от нее и лишь немного крупнее уральских сов Японии. самцов Хорда крыла составляет от 293 до 335 мм (от 11,5 до 13,2 дюйма), а у самок - от 317 до 355 мм (от 12,5 до 14,0 дюйма). Один самец весил 630 г (1,39 фунта), а небольшая выборка самок весила от 608 до 842 г (от 1,340 до 1,856 фунта). [ 5 ] [ 17 ] [ 20 ] Более других русских подвидов эта раса похожа на S. u. Liturata в Европе, но отдаленно разделены по распространению. По сравнению с этой расой, а также с иногда пересекающимися S. u. yenisseensis , в С. у. nikolski голова, затылок и плечи обычно имеют коричневатый цвет (наводящий на мысль о мысе ), который сильнее контрастирует с более белыми светлыми участками. [ 5 ] [ 21 ] [ 20 ]

- С. у. fuscescens ( Temminck & Schlegel , 1847) — эта японская раса встречается от западного и южного Хонсю до Кюсю . Эта раса довольно маленькая. Несмотря на то, что иногда их считают самой маленькой расой, на самом деле они немного крупнее уральских сов, обитающих на Хоккайдо . [ 4 ] [ 20 ] самцов Хорда крыла составляет от 301 до 311 мм (от 11,9 до 12,2 дюйма), а у самок - от 315 до 332 мм (от 12,4 до 13,1 дюйма). [ 5 ] [ 17 ] [ 20 ] Верхняя часть тела отмечена отчетливым желтоватым рыжевато-коричневым цветом. Низ же желтовато-ржавый с темно-коричневыми пестринами и часто покрыт округлыми белыми пятнами. Оперение пальцев ног и лапок сплошное буроватое. [ 5 ] [ 20 ]
- С. у. hondoensis (Clark 1907). К этой расе могут относиться S. u. японика . [ 5 ] Если сюда входит японская раса , то эта раса встречается по всему Хоккайдо, вплоть до северного и центрального Хонсю . Вопреки правилу Бергмана , северные совы на Хоккайдо являются самыми маленькими и даже самыми маленькими из всех известных видов уральских сов, тогда как южные совы немного крупнее. В остальном птицы с разных островов внешне похожи. Хорда крыла самцов составляет от 249 до 322 мм (от 9,8 до 12,7 дюйма), а у самок - от 295 до 347 мм (от 11,6 до 13,7 дюйма). Длина хвоста на Хоккайдо составляет от 201 до 235 мм (от 7,9 до 9,3 дюйма), а на Хонсю - от 220 до 244 мм (от 8,7 до 9,6 дюйма). [ 5 ] [ 17 ] [ 20 ] По сравнению с другими японскими птицами (например, S. u. fuscescens ) и большинством других рас в целом имеет довольно ржаво-коричневый цвет, в то время как у большинства особей практически нет белых пятен на голове, спине и мантии. [ 5 ] [ 20 ]
Распределение
[ редактировать ]Уральская неясыть имеет большое распространение. В материковой Европе его современное распространение довольно неоднородно: этот вид встречается в центральной Европе на юго-востоке Германии западной и восточной части , в центральных и восточных районах Чешской Республики , на юге Австрии , во всех районах, кроме западной Словении , а также точечно, но широко в нескольких районах западной, Европы. Южная и Северо-Восточная Польша . [ 1 ] [ 5 ] [ 18 ] [ 59 ] [ 60 ] [ 61 ] [ 62 ] Распространение в Германии особенно туманно (и, возможно, этому способствует реинтродукция, происходящая от хорошо известной баварской популяции), со свидетельствами того, что уральские совы, по-видимому, обитают (и, возможно, гнездятся) значительно дальше от известных в настоящее время мест обитания в Эгге далеко на западе и загадочным образом поворачивают скорее севернее, в Гарце и Люнебургской пустоши . [ 5 ] [ 59 ] [ 63 ] В Восточной Европе вид встречается в восточной Хорватии , Боснии и Герцеговине , западной Сербии , горной западно-центральной Болгарии , горной центральной Румынии , большей части Словакии , юго-западной Украине , южной и восточной Литве , северной Беларуси , восточной Латвии и большей части Эстонии. . [ 1 ] [ 5 ] [ 18 ] [ 59 ] В Скандинавии ее распространение довольно широко, хотя она встречается только в юго-восточной части Норвегии , поскольку уральские совы встречаются на большей части территории Швеции и Финляндии , но отсутствуют в северных районах, а также в южной Швеции (в основном в районе полуострова). ). [ 1 ] [ 18 ] [ 30 ] Его ареал в России обширен, но он отсутствует в районах с неблагоприятной средой обитания. В западной и европейской части России он встречается на юге примерно до Брянска , Москвы и северной Самары, непрерывно на север до Калининграда , южной части Кольского полуострова и Архангельска . [ 1 ] [ 32 ] В одноименном Уральском регионе встречается примерно от Коми на юг до Каменска-Уральского . [ 1 ] [ 32 ] На всей территории Сибири уральская неясыть встречается широко, прерывая свой типичный ареал в предгорьях Алтайских гор на западе и встречается на севере примерно до Батагая на востоке. [ 1 ] [ 32 ] [ 64 ] Распространение вида продолжается от Дальнего Востока России до Охотского побережья и Магадана , Хабаровского края и Сахалина . [ 1 ] [ 32 ] [ 20 ] За пределами России ареал уральской совы простирается до северо-восточной Монголии , северо-восточного Китая , почти до Пекина , вплоть до Шаньдуна и по всему Корейскому полуострову . [ 1 ] [ 5 ] [ 20 ] [ 65 ] [ 66 ] распространена на всех пяти основных островах Японии Уральская неясыть также (т.е. отсутствует только на Окинаве / островах Рюкю на юге). [ 1 ] [ 20 ] [ 12 ] Сообщалось о бродяжничестве в Европе и России, что может быть причиной появления этого вида почти по всей Германии. Кроме того, существует 16 записей этого вида в северной Италии . [ 4 ] [ 67 ]
среда обитания
[ редактировать ]
Уральские совы, как правило, обитают в зрелом, но не слишком густом девственном лесу , который может находиться в хвойных , смешанных или лиственных районах. Обычно они предпочитают находиться рядом с отверстием . Они часто представляют собой лесные болота с влажной почвой под ногами, заросшей смесью елей , ольхи и/или березы , или влажную пустошь с редкими деревьями. [ 4 ] [ 5 ] [ 8 ] Преобладающими деревьями на большей части ареала часто являются ели , пихты и сосновые леса на севере и ольха , бук и береза со смесью вышеуказанных хвойных пород на юге. [ 5 ] Довольно часто они приспосабливаются к высокогорным лесам в горах , но в отдаленных диких местностях они могут одинаково хорошо адаптироваться и к местам ниже уровня моря. [ 4 ] [ 8 ]
В Карпатах они, как правило, предпочитают почти исключительно буковые леса, обычно на высоте от 250 до 450 м (от 820 до 1480 футов) над уровнем моря. Лесные характеристики этих лесов с преобладанием бука показали, что в ходе ведения лесного хозяйства им необходимо не менее 100 га (250 акров) леса для сохранения, при этом некоторым частям леса должно быть не менее 45–60 лет. Карпатские уральские совы обычно обитают вдали от человеческих жилищ и опушек лесных массивов, не окруженных лесом, и обычно избегают участков леса с крутыми склонами или густым подлеском . Карпатские птицы часто предпочитали места с полянами с промежутками между деревьями, часто около 25 м (82 фута) или около того, и обычно с большим количеством сломанных деревьев . Молодые совы Карпатских птиц, переселившиеся после расселения, проявляют менее сильные предпочтения в отношении среды обитания и могут использовать лесные коридоры , которые часто связаны с оставшимися идеальными местами обитания. Сообщается, что страны Словакии , Словении , а затем Румынии имеют самую обширную идеальную среду обитания в Карпатах и, как следствие, имеют самую высокую локальную плотность уральских сов, возможно, во всей Европе. [ 8 ] [ 68 ] [ 69 ] [ 70 ] с преобладанием бука , также, по-видимому, предпочитали леса Уральские совы, вновь интродуцированные в Баварский лес , причем старые поросли предпочитали обильное пребывание на солнце. Баварские совы обитали в районах, которые также часто были богаты крупными млекопитающими , поскольку их предпочтение доступа к частям леса со сломанными деревьями и просветами часто совпадало. [ 71 ] Дальше на север в Латвии обитаемые леса обычно были намного старше, чем преобладали в региональной среде, обычно с предпочтением лесных участков с деревьями возрастом не менее 80 лет. [ 72 ] Финские популяции, по-видимому, чаще всего встречаются в лесах с преобладанием ели , обычно имея незаметно обособленные лесные предпочтения, за исключением симпатических видов сов, за исключением бореальной совы , которая также предпочитала еловые участки, но встречалась более регулярно, когда доминирующие уральские совы немногочисленны. [ 73 ] В тайге западной Финляндии было обнаружено, что биоразнообразие в окрестностях гнезд уральской совы постоянно выше, чем за пределами этих окрестностей, что делает уральскую неясыть, возможно, « ключевым видом » для местной экосистемы . [ 74 ] речные леса с березой и тополем часто используются В тайге , а еловые или пихтовые леса ( горная тайга ) в районе реки Уссури — . [ 5 ] [ 8 ] Обычно в северных регионах, таких как Финляндия и западная Россия , где в Лапландии уральская сова, вероятно, достигает самой северной части своего ареала, она адаптируется к субарктическим районам, возможно, вплоть до границы леса , но не адаптируется так же хорошо, как великая сова. серая сова - в районы карликового леса к югу от тундры , обычно ей нужны более высокие и зрелые леса к югу от нее. [ 4 ] [ 8 ] [ 30 ]
Исторически сложилось так, что они обычно встречаются в отдаленных, малонарушенных районах вдали от человеческого жилья. [ 5 ] [ 8 ] Уральская неясыть в основном ограничена присутствием в районах, где фрагментация леса произошла или преобладают парковые условия, в отличие от более мелкой, более адаптивной желто-коричневой совы , которая благоприятно акклиматизируется в таких районах. [ 4 ] [ 8 ] Напротив, в некоторых пригородных районах России, таких как столичные парки и сады, при условии, что среда обитания благоприятна и поощряет популяцию добычи, известно, что уральская неясыть успешно встречается. Некоторые города, в регионе которых обитают некоторые популяции уральских сов, - это Чкалов , Киров , Барнаул , Красноярск и Иркутск , а иногда даже Ленинград и Москва . [ 4 ] [ 75 ] Изменения в способах гнездования в связи с возведением гнездовых ящиков почти позволили уральским неясытьм гнездиться необычно близко к жилищам человека в западной части ареала, особенно в Финляндии . [ 8 ] Исключительный рекорд синантропизации этого вида для Европы был зафиксирован в Кошице , Словакия , где в период с ноября по июнь наблюдалось 10-15-летнее явное увеличение неизвестного количества сов. [ 76 ] Было зарегистрировано, что по крайней мере одна уральская сова обитала в городе Любляна в Словении, но не было никаких доказательств того, что она смогла размножиться или обосноваться на территории, учитывая ограниченность лесных массивов в окрестностях. [ 77 ]
Поведение
[ редактировать ]Уральскую сову часто считают ночной , пик активности приходится на сумерки и незадолго до рассвета . Однако в целом и поскольку они в основном обитают в таежной зоне, где очень длинные летние дни являются нормой, а зимой - темное время суток, уральские совы нередко полностью активны в светлое время суток в теплые месяцы, высиживая потомство. Предположительно зимой они в основном активны ночью. Таким образом, этот вид правильнее классифицировать как катемерный, как и большую часть его основной добычи . [ 4 ] [ 5 ] [ 78 ] [ 79 ] На широкий диапазон времени активности и частичную адаптацию к дневной активности указывают также относительно маленькие глаза, которыми обладает этот вид. [ 4 ] Это сильно контрастирует с коричневой совой , которая почти всегда ведет ночной образ жизни. [ 4 ] [ 80 ] В течение дня уральские совы могут отдыхать на насесте, который чаще всего представляет собой ветку рядом со стволом дерева или в густой листве. [ 5 ] Обычно уральские совы не слишком пугливы, и к ним можно подойти довольно близко. [ 5 ] Исторически сложилось так, что европейские наблюдатели за птицами часто считают этот вид довольно неуловимым и трудным для наблюдения. Однако по мере того, как этот вид акклиматизировался в гнездовых ящиках ближе к местам частого пребывания человека, особенно в Фенноскандии , количество встреч резко возросло. [ 4 ] [ 80 ]
Территориальность и движения
[ редактировать ]Уральская неясыть — очень территориальный и жилой вид, который, как правило, склонен оставаться на одном и том же участке обитания в течение всего года. [ 5 ] В то время как большинство бореальных сов, таких как большая серая неясыть и бореальная неясыть , обычно склонны к кочевничеству и резким перемещениям , при этом почти вся популяция следует популяционному циклу своей основной добычи, уральская сова редко покидает свой ареал, даже когда популяции добычи снижаться. За исключением больших серых видов, таких как уральские, большинство видов сов рода Strix являются одновременно территориальными и немигрирующими. [ 30 ] [ 81 ] Территории обычно поддерживаются песнями, которые чаще всего произносит самец проживающей пары. Это вполне норма для сов почти во всех уголках мира. [ 4 ] [ 8 ] Вероятно, из-за их естественной нехватки, известно очень мало свидетельств из первых рук о территориальных боях между взрослыми особями, но предположительно они происходят, поскольку уральские совы могут быть довольно агрессивными совами (или, по крайней мере, в контексте защиты своих гнезд). [ 4 ] [ 8 ] [ 30 ] Однако, согласно исследованию, проведенному на юге Польши, уральские совы, как правило, менее агрессивны в периоды отсутствия размножения, чем коричневые совы по отношению к другим совам, и могут быть немного терпимы к более мелким видам сов на своем домашнем ареале, в то время как рыжие не так агрессивны. [ 79 ] То, что уральская неясыть немного менее агрессивно территориально, чем коричневая неясыть, также подтверждается исследованием, проведенным в Словении, когда коричневые неясыть более энергично кричали на записанные крики и начали более агрессивно атаковать чучела уральских , бореальных и сов своего собственного вида, чем это было раньше. уральские совы на любой из одних и тех же заявленных раздражителей. [ 82 ] Что касается движений, то в отличие от малоподвижных взрослых особей неполовозрелые особи могут преодолевать расстояния примерно до 150 км (93 мили). Случайная особь может забрести еще дальше и остаться на некоторое время в районе зимовки. [ 5 ] Небольшое количество сбившихся с места молодых уральских сов может нерегулярно встречаться в юго-восточной Европе за пределами типичного ареала этого вида. [ 5 ] [ 30 ] Сообщалось о некоторых косвенных свидетельствах того, что уральские совы спускались с гор в горах Японии во время снегопада . сильного [ 83 ] В суровые зимы сибирская популяция несколько перемещается на юг, поскольку численность хищных животных резко падает, а сами совы подвергаются риску замерзания. [ 4 ] [ 5 ] [ 30 ] [ 32 ]
Диетическая биология
[ редактировать ]Это довольно мощный вид совы. Однако, как и совы почти всех размеров, в основном он предпочитает ловить мелкую добычу относительно себя, особенно мелких млекопитающих . В подавляющем большинстве диетических исследований от 50 до 95% пищи приходится на млекопитающих. [ 4 ] [ 5 ] [ 7 ] [ 8 ] Добыча состоит в основном из различных видов грызунов , хотя землеройки и, в некоторых случаях, кроты также могут быть постоянным источником пищи. [ 4 ] [ 5 ] [ 8 ] Кроме того, довольно часто можно ловить любые виды мелких млекопитающих, размером с зайца (хотя обычно молодых), а также различное количество птиц , амфибий и беспозвоночных , причем рептилии и, возможно, рыбы являются очень редкой добычей. [ 4 ] [ 7 ] [ 30 ] Известно, что в широком распространении уральская неясыть потребляет более 200 видов добычи, из которых более 80 являются млекопитающими. [ 4 ] [ 7 ] [ 30 ] [ 84 ] [ 85 ] В результате компиляционного исследования рациона в трех европейских исследовательских регионах и в четыре разных года было обнаружено, что средняя масса тела добычи, взятой на одно гнездо, по оценкам, варьируется от 17,1 до 125 г (от 0,60 до 4,41 унции), при этом средний показатель оценивается в 43,83 г (1,546 г). унция). [ 84 ]
Большая часть охоты осуществляется с окуня. [ 5 ] Обычно они предпочитают добычу, которая попадает на открытые участки леса, а не ту, которая часто посещает лесную подстилку. [ 8 ] [ 86 ] В Словакии , помимо полян и лугов возле высоких лесов, уральские совы в некоторой степени приспособились к охоте на открытых территориях, антропогенных, включая участки фрагментации леса и даже сельскохозяйственные районы , а также небольшие города и деревни , если они примыкают к зрелому лесу. патчи. [ 7 ] Методы охоты этого хищника практически не изучались. Вполне возможно, что это в какой-то степени охотник, сидящий какое-то время на высоком дереве, пока поблизости не станет видна добыча. [ 87 ] Однако в Скандинавии сообщалось, что уральские совы охотятся скорее как ястребы-тетеревятники, а не как все еще охотящиеся серые совы, используя метод охоты с использованием окуня, при котором они перелетают короткими рывками с насеста на насест, при этом полеты должны быть незаметными до тех пор, пока не будет добыта добыча. обнаружен. Известно, что уральские совы не нападают на добычу в активном полете, а почти всегда падают на нее прямо с насеста. [ 4 ] [ 30 ] [ 88 ] [ 89 ] В Японии были предприняты экспериментальные исследования уральских сов, находящихся в полу-неволе, чтобы наблюдать, как совы выбирают, в каких местах охотиться и какую добычу выбирать. Когда уральские совы экспериментально подвергались воздействию участков, где присутствовала добыча, и участков, где добыча отсутствовала, уральские совы добывали корм в обоих случаях, но, похоже, узнавали, на каких участках с большей вероятностью будет еда, и впоследствии более активно добывали корм вокруг них. Когда те же биологи наблюдали за поведением полевых мышей , а также полевок, они аналогичным образом учились и отдавали предпочтение участкам, на которых обитали более крупные виды, а не более мелким. Судя по имеющимся данным, хотя выбор добычи в значительной степени является оппортунистическим, грызуны, подвергающиеся риску голодной смерти в японских исследованиях, с большей вероятностью рисковали добывать пищу на относительно открытых участках земли, поэтому уральские совы отдавали предпочтение им. Судя по японским исследованиям, уральские совы, похоже, способны улучшить свои модели использования ресурсов по мере накопления опыта в окружающей среде. [ 88 ] [ 90 ] [ 91 ] Различные исследования, проведенные в Японии, показали, что уральские совы способны охотиться на добычу, которая зимой прячется в мелком снегу или у поверхности снега, но полевки, прячущиеся в субнивальной зоне — эфемерных туннелях, проложенных полевками под глубоким снегом, в значительной степени недоступны для них. совы. [ 92 ] Излишки еды можно хранить в гнезде или на близлежащих складах. [ 5 ] Первый подтвержденный факт поедания падали ), хотя был зарегистрирован, когда уральская сова питалась тушей косули ( Capreolus capreolus существуют и более ранние записи о посещении уральской совой волчьей добычи (хотя неясно, был ли сова кормится в таком случае). [ 93 ] [ 94 ]
Млекопитающие
[ редактировать ]
Полевки , грызуны малого и среднего размера с характерно короткими хвостами, часто считаются наиболее значительной частью рациона почти во всех частях ареала. Как правило, в некоторых частях к западу от своего ареала уральская неясыть связана с двумя видами добычи полевок, в частности, полевок ( Microtus agrestis ) и рыжих полевок ( Myodes glareolus ). Эти полевки, как правило, ведут одиночный образ жизни и довольно широко рассредоточены, но приспосабливаются к благоприятным местам обитания для уральских сов, таким как открытые окраины обширных лесных массивов, и могут быть довольно многочисленными, когда популяция полевок достигает пика . [ 95 ] [ 96 ] [ 97 ] [ 98 ] [ 99 ] Согласно исследованию, проведенному в Финляндии, уральские совы ловили полевых и рыжих полевок, средний вес которых превышал средний вес пойманных биологом в полевых условиях, т.е. вес пойманных совами оценивался в 28,59 г (1,008 унции) для полевок против 26,44. г (0,933 унции) для рыжих полевок, а у пойманных биологами средний вес составил 22,78 г (0,804 унции) для поля против 18,75 г (0,661 унции) для банков. 56% полевых полевок, пойманных уральскими совами, были репродуктивно активны, тогда как 44% рыжих полевок были активны как таковые. [ 97 ] Гельминты, по-видимому, не делали полевок более уязвимыми для хищников, но уральские совы гораздо чаще ловили самцов полевок, чем самок. те, кого поймали в полях, были самцами. Однако беременные самки полевок оказались более уязвимыми, чем самки других возрастных полевок. [ 100 ] Более социально настроенные и сконцентрированные полевки, такие как обыкновенные полевки ( Microtus arvalis ), как правило, встречаются на более обширных открытых территориях, поэтому лесные совы, такие как уральские, обычно охотятся на них относительно редко, но только по сравнению со многими другими хищниками. [ 101 ] Самое крупное известное на сегодняшний день исследование диеты было проведено в Финляндии , где за несколько лет исследования было исследовано 5995 объектов добычи. Согласно этим данным, в рационе там, где она была, предпочтение отдавалось значительно более крупной полевке — водяной полевке ( Arvicola amphibius ). При расчетном среднем весе 177 г (6,2 унции) водяная полевка составляла примерно 22,15% добычи по численности и 52,2% биомассы , тогда как полевые полевки составляли 19,5% по численности и 18% биомассы. . Средний предполагаемый вес добычи в целом в рамках исследования составил 71,7 г (2,53 унции). [ 102 ] Другое, хотя и меньшее по размеру исследование в Центральной Финляндии показало, что полевки лидируют по численности над водяными полевками, 42,7% против 33,9%, среди 1739 объектов добычи соответственно, но при этом эти два вида составляют 17,4% и 69,1% биомассы добычи. [ 103 ] Исследования в Южной Финляндии показали резкие изменения в импорте добычи полевок из-за их популяционных циклов . В южной Финляндии из 3351 объекта добычи в год пика полевок 58,43% по численности и 42,8% по биомассе добычи приходилось на полевок, за которыми следовала рыжая полевка с 12,41% по численности (но только 5,73% по биомассе). и водяные полевки - 22,24% по биомассе (но только 10,1% по численности). В том же районе исследований в неурожайный для полевок год среди 860 объектов добычи полевки по численности отставали от других видов добычи, таких как птицы и землеройки , но водяных полевок здесь ловили более или менее в тех же количествах, что и в годы пика полевок. [ 84 ] Другое исследование оценило средний вес добычи финской уральской совы в 78,1 г (2,75 унции). [ 104 ] Аналогичные пищевые привычки были обнаружены у уральских сов в Швеции: в 2309 объектах добычи водяные полевки составляли 33,1% добычи по численности и 60% биомассы, полевки составляли 30,8% по численности и 14,5% биомассы, а рыжие полевки составляли до 11,8% добычи по численности и 3,3% по биомассе. [ 89 ] [ 105 ] Видеоловушки зафиксировали 187 объектов добычи уральских сов в 5 гнездах в графстве Вермланд , Швеция. Было обнаружено, что полевки уступали по скорости доставки обыкновенной землеройке ( Sorex araneus ) и различным птицам, а такая добыча небольшого размера требовала частых поставок, хотя это было компенсировано. с изрядным количеством молодых зайцев , очевидно, доступных этим совам. [ 106 ]
По мере того как уральская неясыть перемещается на юг, популяция добычи имеет тенденцию становиться немного менее цикличной, но также, по-видимому, сокращается очевидный доступ к крупной добыче, такой как водяная полевка. [ 30 ] [ 107 ] В относительно северной Латвии из 2615 объектов добычи полевки составляли чуть более половины рациона, а кормовая база была относительно разнообразной. Основными видами добычи латвийских сов были рыжая полевка (18,09 % по численности, 9,2 % по биомассе), полевки-полевки (17,13 % по численности, 13,85 % по биомассе), а также европейский крот ( Talpa europaea ) (9,83 % по численности, 12,3% биомассы), последний при зрелой массе около 90 г (3,2 унции)). [ 84 ] В Беларуси были довольно регулярной добычей в годы пика полевок: 45% по численности и 29,9% по биомассе, тогда как в кризисные годы их численность составляла всего 6,7% по численности и 4% по биомассе , виды Microtus в то время как менее цикличные рыжие полевки составляли 29% по биомассе. численности и 20,4% по биомассе в пиковые годы и 38,1% по численности, 23,8% по биомассе в годы кризиса. Более того, в Беларуси численность землероек и кротов увеличилась с 6,4% до 14,3% в годы пика и кризиса (биомасса в годы кризиса составляла 13,1%, хотя только 2,2% приходилось на землероек). [ 108 ] В Бескидах на юго-западе Польши средняя масса добычи из 1039 объектов была довольно низкой — 27,5 г (0,97 унции), при этом основным видом добычи была рыжая полевка, составлявшая в среднем 27,7% добычи по численности и 25% биомасса (средний расчетный вес 24,3 г (0,86 унции)), обыкновенные полевки составляют в среднем 24,9% жертва по количеству и 25,8% биомассы (средний расчетный вес 27,5 г (0,97 унции)), желтошейная мышь ( Apodemus flavicollis ) в среднем 15,73% количества жертвы и 17,4% биомассы (средний расчетный вес 30 г (1,1 унции) )) и европейская сосновая полевка ( Microtus subterraneus ) в среднем 8,9% количество добычи и 5,8% биомассы (средний расчетный вес 17,3 г (0,61 унции)). За исключением полевок и полевых мышей, других видов добычи в Польских Бескидах практически нет. [ 109 ] В Словакии среди 2134 объектов добычи основными видами добычи были желтошейная мышь (21,8%), обыкновенная полевка (18,3%), рыжая полевка (14,3%), полосатая полевая мышь ( Apodemus agrarius ) (4,9%) и европейская полевка. сосновая полевка (4,7%). [ 7 ] У вновь интродуцированных в Баварские леса уральских сов среди небольшой выборки из 117 объектов добычи оказались в основном отобраны сходные мелкие виды грызунов, такие как виды Apodemus (16,2%), полевки (15,3%), рыжие полевки (13,6%). ), а также неопознанные полевки. [ 110 ] Рационы уральских сов в юго-восточной части европейского ареала в основном зарегистрированы в Словении . Самое широкое исследование показало, что из 1268 объектов добычи 45,9% рациона составляли полевки, особенно рыжая полевка (38,1% по численности, 26,24% по биомассе), а еще 34,2% составляли виды Apodemus (а также 28,2% биомассы) и значительную часть сони , особенно большую съедобную соню ( Glis glis ), на 6% по численности и 25,5% по биомассе. Более локально, в меньшем квартале в Словении, до 58,8% по численности и 94,4% по биомассе в рационе уральской совы составляет съедобная соня, масса тела взрослой особи которой может варьироваться от 62 до 340 г (от 2,2 до 12,0 унций). [ 84 ] [ 111 ] [ 112 ] [ 113 ]
Дальше на восток и за пределы Европы уральские совы в целом полагаются на мелких грызунов. В одном из самых западных исследований, проведенном в Мордовии ( Россия ), из 426 объектов добычи основными видами добычи были обыкновенные полевки (41,8%), рыжие полевки (31,4%), тундровые полевки ( Microtus oeconomus ) (5,9%) и уральская полевая мышь. ( Apodemus uralensis ) (3,1%). [ 114 ] На пяти участках исследований в Приуралье среди 870 объектов добычи 75,2% составляли млекопитающие. 29,54% рациона на Урале составляли рыжие полевки, 16,55% - обыкновенные полевки, 5,52% - лесная мышь ( Apodemus sylvaticus ) и 4,83% - неопознанные Myodes . полевки [ 115 ] Дальше на восток общая зависимость от миодов , также известных под общим названием «красные полевки», за исключением далеко обитающих рыжих полевок, кажется, несколько возрастает. [ 116 ] Гораздо восточнее, в тихоокеанском побережье Приморского края , всего было исследовано 1163 объекта добычи. Здесь в основном уральские совы добывали в основном грызунов сходного характера, хотя в основном разных видов, таких как красная полевка ( Myodes rutilus ) (39,2% по численности), камышовая полевка ( Microtus fortis ) (24,24%), Apodemus сп. (9,7%), корейская полевая мышь ( Apodemus peninsulae ) (3,52%). [ 117 ] На островах Японии небольшие местные расы уральских сов, кажется, в некоторой степени занимают диетическую универсальную Стрикс совы экологическую нишу , аналогично тому, как это делает серая неясыть на западе, однако основные части ее рациона все еще занимают совы Стрикс. многие из тех же родов добычи, взятые в других местах. В горах Яцугатаке 1026 мелких млекопитающих были обнаружены в 17 гнездах уральской совы, из которых виды Apodemus составили 71%, за ними следовали полевки - 24% и японская землеройка ( Urotrichus talpoides ) - 5%. [ 92 ] В Косимидзу , Хоккайдо , из 266 объектов добычи из 111 общих погадок рацион возглавляли серая красная полевка ( Myodes rufocanus ) (25,2%), крупная японская полевая мышь ( Apodemus speciosus ) (17,7%) и мелкая японская полевая мышь. ( Apodemus argenteus ) (15,8%). [ 118 ]

Землеройки почти 20 видов ловятся более или менее по всему ареалу уральской совы. Хотя обычно это не имеет большого значения для массы добычи сов, поимка землеройок может позволить совам предотвратить голод. [ 7 ] [ 102 ] [ 109 ] [ 117 ] [ 118 ] [ 119 ] [ 120 ] Самая маленькая добыча млекопитающих и позвоночных, которую, как известно, добывают уральские совы, - это евразийская землеройка ( Sorex minutissimus ), средняя масса тела взрослой особи которой составляет около 2,5 г (0,088 унции). [ 102 ] Другими мелкими млекопитающими, которых уральские совы обычно ловят в довольно небольших количествах, являются летучие мыши , белки-летяги и ласки . [ 7 ] [ 102 ] [ 117 ] [ 118 ] Уральские совы также поедают более крупных млекопитающих, и их можно считать случайной дополнительной добычей, несмотря на то, что их биомасса часто составляет значительную часть. Этот вид поедает добычу млекопитающих, крупнее, чем типичные полевки и мыши, гораздо меньше, чем более крупные совы, такие как филины . [ 4 ] [ 121 ] Среди них есть несколько видов белок , большинство из которых во взрослом состоянии весят более 200 г (7,1 унции), в Европе исключительно дальневосточная красная белка ( Sciurus vulgaris ), а также крысы , особенно 300 г (11 унций). ) коричневая крыса ( Rattus norvegicus ), составлявшая почти 20% рациона в исследовании на Хоккайдо. [ 117 ] [ 92 ] [ 118 ] Взрослых ежей нескольких видов время от времени ловят в Европе, а европейские ежи ( Erinaceus europaeus ), пойманные в Финляндии, по оценкам, весят в среднем около 652 г (1,437 фунта). [ 7 ] [ 102 ] Завезенные ондатры ( Ondatra zibethicus ), которые в среднем весят около 1000 г (2,2 фунта) в выловленном состоянии, также используются в финских исследованиях. [ 103 ] Известно, что они охотятся на японскую гигантскую белку-летягу ( Petaurista leucogenys ), средний вес которой составляет около 1150 г (2,54 фунта). [ 122 ] Одной из крупных жертв среди млекопитающих, широко ассоциирующихся с уральскими совами, являются зайцы , хотя они редко встречаются в значительном количестве в рационе, но, по-видимому, на большей части ареала их поедают оппортунистически. Почти 25% их биомассы могут составлять зайцы, а зайцы являются крупнейшей известной добычей уральских сов. [ 4 ] [ 106 ] Охотясь на зайцев, уральские совы обычно ориентируются на относительно молодые экземпляры. В Европе как на зайца-беляка ( Lepus europaeus ), так и, особенно, на зайца-беляка ( Lepus timidus можно охотиться ). Средний размер зайцев-беляков, пойманных в Финляндии, по разным оценкам составляет от 173 до 2000 г (от 0,381 до 4,409 фунта), при этом средний средний вес составляет около 500 г (1,1 фунта). В исключительных случаях уральские совы могут ловить даже взрослых зайцев-беляков, средний вес которых составляет около 2900 г (6,4 фунта), но это может быть неподтверждено. [ 84 ] [ 102 ] [ 103 ] [ 23 ] [ 123 ] [ 124 ] Японских зайцев ( Lepus brachyurus ) также можно ловить в Японии. [ 125 ]
Другая добыча
[ редактировать ]
Второй по важности группой добычи после млекопитающих (хотя и значительно позади) являются птицы . Птицы также являются самой разнообразной группой добычи в рационе уральской совы: зарегистрировано более 100 видов птиц. [ 4 ] [ 7 ] [ 84 ] Согласно исследованиям, проведенным в Европе, птицы обычно составляют менее 10% рациона по численности. [ 7 ] [ 84 ] Исключительно высокая численность, условно говоря, отмечена в Фенноскандии , особенно когда численность полевок снижается. В годы плохих полевок в Финляндии из 860 объектов добычи 32,2% по численности и 40,25% по биомассе составляли птицы. В частности, дрозды рода Turdus были наиболее зарегистрированными кормовыми родами в эти годы, составляя в среднем 19,4% по численности и 23,6% по биомассе. [ 84 ] В округе Вермланд , Швеция, птицы составляли 25,6% добычи по численности и 32,67% биомассы. Сообщалось о широком разнообразии птиц, причем чаще всего выявлялись дрозды. [ 106 ] В Центральной и Южной Европе реинтродуцированная популяция Баварского леса Германии может показать самые высокие результаты для птиц - всего 11,1%. [ 110 ] Тем не менее, в ходе исследования диеты в городе Кошице , Словакия полагаются почти исключительно на голубей и голубей , было обнаружено, что адаптированные к городским условиям уральские совы в качестве пищи , а именно на евразийского ошейникового голубя ( Streptopelia decaocto ) и сизого голубя ( Columba livia ). что составило 76,7% и 13,4% содержания 16 гранул соответственно. [ 76 ] В России птицы в целом были гораздо более значимы на Урале по сравнению с другими известными исследованиями, составляя 17,01% от общего числа 870 объектов добычи в разные годы и 47,6% среди 146 объектов добычи, более локально вблизи городов и деревень Пермского края . [ 115 ] [ 117 ] В Японии птицы занимают несколько большее место в рационе уральских сов по сравнению с европейскими данными, обычно составляя более 10% продуктов питания. [ 92 ] [ 118 ] Наибольшее количество птиц, зарегистрированных в рационе японских уральских сов, было из 36 погадок, найденных в Сакаиде, Кагава , которые в основном включали мелких птиц, в частности евразийского воробья ( Passer montanus ) (78,6% по численности) и белощекого скворца. ( Sodiopsar cineraceus ) (3,6%). [ 126 ]
Охота уральской совы на птиц, по-видимому, почти полностью основана на возможности, поскольку существует мало свидетельств того, что она выслеживает птиц какого-либо конкретного типа, а просто из-за шанса встречи или обнаружения. [ 4 ] [ 88 ] [ 90 ] Дроздов, вероятно, ловят во многих частях ареала из-за их относительной общности в типах среды обитания, используемых уральскими совами, в то время как другие воробьиные широко распространенные птицы также часто имеют схожие предпочтения в среде обитания, такие как мухоловки Старого Света , зяблики и синицы . [ 7 ] [ 106 ] [ 85 ] [ 117 ] Большинство воробьиных, а также большинство идентифицированных птиц в целом, по размеру находятся между зябликом ( Fringilla coelebs ) со средним весом 23,9 г (0,84 унции) и евразийской сойкой ( Garrulus Glandarius ) со средним весом 160 г. (5,6 унций). [ 4 ] [ 8 ] [ 16 ] Добыча птиц иногда может достигать размера золотого гребня ( Regulus regulus ) весом 5,2 г (0,18 унции), который является самым маленьким видом птиц в Европе. [ 8 ] [ 16 ] Средний расчетный вес птиц, пойманных в Финляндии, составил 83,4 г (2,94 унции), что немного превышает расчетный вес пойманных там млекопитающих. [ 102 ] самых разных врановых Можно поймать , начиная от лазурнокрылых сорок ( Cyanopica cyanus ) весом 96 г (3,4 унции) и заканчивая воронами-падальщиками ( Corvus corone ) весом 570 г (1,26 фунта), но обычно их обнаруживают в в диетических исследованиях их численность невелика, и эти совы гораздо менее плодовитые охотники на врановых, чем некоторые дневные хищники, такие как ястребы . [ 4 ] [ 7 ] [ 101 ] [ 85 ] [ 115 ] [ 127 ] За исключением голубей и голубей, добычу птиц, не относящихся к воробьиным, обычно добывают относительно нечасто, хотя некоторые семейства добычи, такие как дикие птицы и дятлы, также добываются довольно широко. [ 7 ] [ 85 ] [ 128 ] Необычное предпочтение относительно крупной птицы было обнаружено в ходе исследования на Урале , где наиболее часто выявляемыми видами птиц были рябчик массой 429 г (15,1 унции) ( Bonasa bonasia 490 г (1,08 фунта) ) и ворона весом ( Corvus cornix ). ). [ 16 ] [ 115 ] В редких случаях уральские совы могут догнать добычу птиц размером примерно до их собственного размера или несколько больше, то есть до или чуть более 1000 г (2,2 фунта) при средней массе тела, например, взрослую крякву ( Anas platyrhynchos ), гоголь обыкновенный ( Bucephala clangula ), тетерев ( Tetrao tetrix ), фазан обыкновенный ( Phasianus colchicus ), курица ( Gallus Gallus Domesticus ), чернокоронковая кваква ( Nycticorax nycticorax ) и некоторые ястребиные , а также, насколько известно, только молодые особи более крупного еще западного глухаря ( Tetrao urogallus ). [ 7 ] [ 85 ] [ 114 ] [ 106 ] [ 118 ] [ 129 ] Помимо вышеупомянутых семейств, к другим нечасто добываемым птицам добыча относятся кукушки , козодои , кулики , крачки , катки и удоды . [ 7 ] [ 85 ] [ 102 ] [ 118 ]
Вероятно, в немалой степени из-за скудности герпетологического разнообразия добычи в умеренных и часто холодных местах обитания уральских сов, рептилии являются очень редкой добычей для этого вида. [ 108 ] [ 106 ] Однако иногда можно поймать лягушек , когда уральская сова попадается на них в теплые месяцы года. Обычно почти в любой части ареала лягушки и другие амфибии составляют менее 6% регионального рациона этого вида. [ 4 ] [ 7 ] [ 23 ] [ 80 ] [ 130 ] В небольшом шведском исследовании, проведенном в округе Вермланд , исключительные 12,2% доставленной добычи составляли обыкновенные лягушки ( Rana temporaria ). [ 106 ] Несмотря на исторические записи о редком обнаружении рыбьей чешуи в некоторых гранулах, в ходе диетических исследований нет известных сообщений о подтвержденном хищничестве рыбы со стороны уральских сов. [ 4 ] [ 8 ] [ 30 ] Существует мало свидетельств того, что на большей части ареала уральские совы тратят много времени на преследование или питание насекомыми и другими беспозвоночными , поскольку пищевое вознаграждение для довольно крупной совы, обычно охотящейся на такую добычу, которая может весить всего около 0,2 г, невелико. (0,0071 унции). [ 4 ] [ 106 ] Однако иногда скопления насекомых, таких как жуки-древоточцы . сову могут привлечь [ 8 ] [ 30 ] В Баварском лесу более 6% рациона составляли насекомые, в основном неопознанные жуки. [ 110 ] Несмотря на то, что большинство исследований, проведенных в Японии, показывают, что около 95% продуктов питания составляют млекопитающие и, во вторую очередь, птицы, иногда сообщается о более обширной связи с добычей насекомых. В префектуре Кагава исключительно 24% рациона составляли насекомые, а в Киото было замечено, что уральские совы регулярно преследуют и поедают японских жуков-носорогов ( Allomyrina dichotoma ). [ 131 ] В Приморском крае далее отмечались случаи охоты уральских сов на даурского рака ( Cambaroides dauricus ). [ 117 ]
Межвидовые хищнические отношения
[ редактировать ]Уральские совы регулярно живут одновременно в разных частях своего ареала с другими видами сов. Большинство симпатрических видов также отдают предпочтение мелким млекопитающим , в основном полевок , особенно когда они встречаются в относительно северных местах с умеренным климатом, таких как места обитания уральских сов. [ 4 ] [ 30 ] Таким образом, уральскую сову часто считают конкурентом симпатрических видов сов. [ 4 ] [ 30 ] В частности, уральскую сову часто сравнивают и изучают в областях, близких к ее близкому родственнику, желтовато -коричневой сове . Рыжая неясыть сегодня может сосуществовать с уральскими совами во многих частях европейского ареала последних, включая южную Фенноскандию , восточную и центральную Европу и европейскую часть России . [ 1 ] [ 4 ] [ 89 ] Пищевые привычки этих двух видов в основном совпадают, как, например, в Уппланде , Швеция , где виды и группы добычи были почти идентичны по степени важности для их рациона (т. е. полевки, водяные полевки, виды Apodemus , птицы и лягушки). [ 89 ] На горе Крим в Словении и коричневая, и уральская неясыть питались одним и тем же основным видом добычи, но было показано, что коричневая неясыть способна адаптироваться к альтернативной добыче в течение низкой части цикла популяции полевок , в то время как уральская неясыть была менее адаптивна к альтернативной добыче. [ 132 ] Диета коричневых и уральских сов в Баварском лесу Германии была во многом схожей , но рыжие виды потребляли пропорционально меньше полевок и больше насекомых, и, по оценкам, они ловили добычу, вес которой в среднем на 43% меньше, чем у уральских сов. [ 110 ] В Финляндии ширина пищевой ниши уральской совы примерно на 73% перекрывалась с коричневой неясытью, но средний размер добычи у уральской совы был более чем в два раза больше: 38,4 г (1,35 унции) у коричневой совы против 78,1 г (2,75 унции) у уральской совы. , и было зарегистрировано, что коричневая неясыть охотится на добычу, не являющуюся млекопитающими, значительно чаще, чем уральские совы. [ 104 ] В Беларуси размеры добычи коричневой совы были в среднем на 31–49% меньше (с использованием двух разных методов расчета), чем у уральской совы, но более мелкие виды имели значительно большую ширину пищевой ниши , которая в среднем составляла 12,96 для коричневой совы и 5,48 для уральской. [ 101 ] По сравнению с желто-коричневой совой, уральская неясыть значительно крупнее по размеру и размеру когтей (что примерно на 30% больше на Урале), причем форма когтей еще раз указывает на их различную экологическую роль: у желто-коричневых сов они пропорционально короче и более изогнуты. В соответствии с ее более разнообразным рационом, а также более тяжелым и прямым у уральской совы для борьбы с относительно более крупными млекопитающими. [ 4 ] [ 73 ] Известно, что когда ареалы коричневой и уральской совы пересекаются, уральская неясыть имеет тенденцию доминировать, а иногда и убивать коричневую неясыть. [ 89 ] [ 104 ] [ 133 ] В Словении уральские совы на самом деле более последовательно реагировали на записанные крики коричневых сов (40% воздействий на крики, вызывающие голосовую или физическую реакцию), чем на записи криков представителей своего вида (34% воздействий на крики, вызывающие ответ). [ 134 ] Однако случаи межвидового убийства между видами довольно редки, и недавнее исследование показывает, что относительно неоднородное распространение коричневой совы в северных районах, таких как Фенноскандия, скорее всего, связано не с этой конкуренцией, а, скорее, с типом региональной среды обитания, благоприятствующим уральской сове. Рыжая неясыть не так хорошо подходит для таежных местообитаний, как уральские виды, и локально менее приспособлена к альтернативным местам гнездования (т. е. гнездованию в районах без дупл деревьев или гнездовых ящиков , тогда как на Урале локально можно также использовать старые птичьи гнезда и пни деревьев). ). [ 89 ] [ 104 ] [ 135 ] Потепление средних температур в Фенноскандии , по-видимому, отрицательно повлияло на многие виды сов из-за ограничения цикла полевок, но желтовато-коричневая неясыть, обитающая на юге, оказала наименьший эффект на местном уровне из-за снижения численности полевок (и, несмотря на их общая способность адаптироваться к различной добыче), в то время как уральская неясыть, по крайней мере, в центральной Финляндии, пока не демонстрирует такого резкого негативного эффекта из-за этого. [ 136 ] Точно так же коричневая неясыть и уральская неясыть в значительной степени разделены по высотному диапазону и среде обитания в Восточной Европе (например, Карпаты ) и Польше , при этом мало доказательств того, что это связано с межвидовой конкуренцией или хищничеством, а скорее с подходящими горными средами обитания. Здесь серая неясыть обычно встречается на более низких высотах, а уральская неясыть населяет предгорные леса на возвышенностях. В целом, несмотря на более высокую плотность Урала в некоторых горных частях Центральной и Восточной Европы, серая неясыть гораздо более многочисленна во всех странах этого региона, чем уральская неясыть. [ 137 ] [ 138 ] [ 139 ] В Словакии 13,3% территорий уральской совы пересекаются с территориями коричневых неясытей, что является относительно небольшим перекрытием, учитывая общую окружающую среду. [ 140 ] Несмотря на предполагаемое доминирование уральской совы, в Словении было замечено, что серая неясыть яростно нападала на уральскую сову, пока она не покинула окрестности, хотя неясно, было ли это территориальным столкновением или нападением хищников . [ 138 ] В Польше во время зимнего расселения рыжие совы смогли временно использовать части территорий уральских сов, воспользовавшись менее агрессивным поведением уральских неяс вне сезона размножения. [ 133 ]

Уральские совы обычно не встречаются с другими совами Стрикс , за исключением желтовато-коричневой совы, но на большей части своего ареала пересекаются также с большой серой совой , которая, кроме того, обитает дальше на север и в Америку . Несмотря на свои большие размеры, большая серая сова является специалистом по питанию полевок, полагаясь почти исключительно на них. На ограничения в питании указывает морфология стоп: у большой серой совы когти заметно тоньше, меньше и менее мощные на вид, чем у уральской совы, несмотря на немного больший общий размер стопы (в соответствии с массой тела) у обыкновенной совы. отличный серый. [ 4 ] [ 104 ] Было показано, что в тайге , которую предпочитают как большая серая, так и уральская неясыня, большие серые совы гнездятся в самых разных местах, причем в целом типов гнезд больше, чем на Урале, но они обычно не используют дупла деревьев , как две другие. Европейские совы Strix редко встречаются в районах, где хвойные деревья не преобладают нигде в своем ареале, например, в прибрежных зонах, где лес может разнообразиться и смешаться с лиственными деревьями. [ 102 ] [ 104 ] [ 23 ] Считается, что разница в использовании мест гнездования и более узкие пищевые привычки большой серой совы смягчают самую серьезную конкуренцию между большой серой совой и уральской совой. [ 78 ] [ 104 ] [ 80 ] Однако более крупная сова, чем любой другой вид, — это евразийский филин . Уральская неясыть имеет значительно меньшее тело (примерно в три раза легче), ее размах крыльев примерно на 35% меньше, а когти и лапы меньше, поэтому конкурентное преимущество очевидно даже между морфологическими видами этого вида. [ 4 ] [ 121 ] Филин также ловит большое количество мелкой добычи, такой как полевки , но заметно более изменчив в альтернативной пище. При аналогичном, но, как правило, гораздо более широком распространении (как по широте, так и по долготе ), филины потребляют примерно в три раза больше видов добычи, чем уральские совы, включая гораздо больше видов из всех таксонов добычи, а также встречаются в более широком диапазоне ареалов. типы. Будучи гораздо крупнее и (исходя из морфологии), предположительно гораздо более мощным, филин способен поймать гораздо более крупную добычу, чем уральская неясыть. [ 4 ] [ 141 ] [ 142 ] [ 143 ] В более локальном масштабе, в Финляндии, уральская неясыть съела пропорционально больше европейских водяных полевок , птиц и амфибий в целом, чем филины, но съела гораздо меньше охотничьих птиц . [ 80 ] Урал, вероятно, по возможности избегает филин. Судя по имеющимся данным, уральские совы довольно регулярно встречаются в Северной Остроботнии , где филины редки или отсутствуют, но в Южной Остроботнии уральские совы встречаются редко, возможно, потому, что филины относительно обычны. [ 121 ] Однако различия в среде обитания также могут привести к разделению этих двух видов. В Финляндии филин предпочитает леса с преобладанием сосны , а на Урале – леса с преобладанием ели . [ 73 ] В других местах филины могут часто посещать скалистые участки, где это возможно (для гнездования), и часто охотятся на более разнообразных и часто более открытых местах, а в таких местах уральские совы встречаются редко или вообще отсутствуют. [ 141 ] [ 144 ] Предполагается, что, поскольку уральская неясыть меньше большой серой совы и евразийского филина, в целом она может питаться меньшим количеством пищи, чем они. Подсчитано, что за 6-месячный период уральской сове требуется около 22,8 кг (50 фунтов) пищи, большой серой сове - около 27,4 кг (60 фунтов), а филину - около 54,8 кг (121 фунт). . [ 80 ] Уральская неясыть может столкнуться с другими более крупными совами, такими как полярная сова ( Bubo scandiacus ) зимой и рыбная сова Блэкистона ( Bubo blakistoni ) на дальнем востоке ареала, но вряд ли будет активно взаимодействовать, учитывая различия в среде обитания. использование. [ 8 ] [ 12 ] [ 66 ]
Помимо вышеупомянутых видов, большинство других видов сов, с которыми уральская неясыть встречается в своем ареале, значительно мельче. Более того, уральская сова имеет тенденцию доминировать над этими видами при встречах, и поэтому ее можно избежать. [ 133 ] Одним из видов, который часто имеет сходную среду обитания и предпочтения в отношении добычи с уральской совой, является бореальная сова . Однако, судя по расстоянию территории, бореальные совы могут более строго избегать желто-коричневой совы, которая, как известно, очень агрессивна в своем территориальном поведении круглый год, в то время как исследования показали, что территориальное исключение со стороны уральских сов в основном ограничивается сезоном размножения. . [ 82 ] было высказано предположение, Однако в лесистой местности на юге Польши что бореальные совы выбирали места для гнезд не из-за того, что избегали неясытых, а исходя из наличия подходящих мест для гнездования. В исследовании на юге Польши коричневые совы обычно встречались чаще в пихтово - еловых лесах. [ 133 ] В Словении было высказано предположение, что бореальные совы случайно выиграли от исключения желтовато-коричневых неясыней со стороны доминирующих уральских сов, когда эти три вида встречались в соседних средах обитания. [ 137 ] Однако данные показывают, что бореальные совы сталкиваются с более высокой смертностью, когда они гнездятся слишком близко к уральским совам, то есть в пределах примерно 2 км (1,2 мили). [ 145 ] Аналогичная связь была обнаружена между уральской совой и евразийской карликовой совой ( Glaucidium passerinum ), видом, размером менее половины бореальной совы. [ 30 ] [ 16 ] Однако карликовая сова обычно выбирала другие типы леса, чем уральские совы, в районах симпатии, в частности в еловых лесах, где Урал обычно редок или отсутствует. Однако экологические взаимодействия были обнаружены в Словении: было замечено, что карликовая сова проявляет антихищническое поведение по отношению к уральским совам. [ 138 ] Несмотря на разное предпочтительное лесное местообитание, 46,3% ареалов уральских и евразийских карликовых сов в Словакии перекрывались. [ 140 ] Довольно мало известно об экологических взаимодействиях между уральской совой и северной ястребом ( Surnia ulula ), еще одной бореальной совой, несмотря на общую склонность к использованию коряг в качестве мест гнездования. Ястребиные совы также являются обычными хищниками-полевками, но у них больше экологических характеристик с большой серой совой, чем у уральских сов, в частности, их кочевое поведение и агрессивные движения . Более того, более сильная склонность ястребиной совы к суточному образу жизни может дополнительно обеспечить определенную степень разделения. [ 4 ] [ 30 ] [ 73 ]
Самый известный аспект межвидовых взаимодействий с другими совами и другими хищниками — это межвидовое хищничество. Уральская сова иногда уязвима для хищников со стороны более крупных хищников, когда происходят встречи. Несомненно, самым опасным хищником, вероятно, является евразийский филин . [ 4 ] [ 146 ] Среди других хищников, которые, как известно, охотятся на уральских сов, — беркут ( Aquila chrysaetos ), восточный могильник ( Aquila heliaca ), горный ястреб ( Nisaetus nipalensis ) и евразийская рысь ( Lynx lynx ), а также дневные хищники, которые лишь немного крупнее. , в том числе северный ястреб-тетеревятник и малый подорлик ( Clanga pomarina ). [ 146 ] [ 147 ] [ 148 ] [ 149 ] [ 150 ] [ 151 ] В большинстве упомянутых выше случаев возраст пойманной хищниками уральской совы обычно не указывается (хотя любой из первых трёх крупных орлов заведомо способен сразить уральскую сову потенциально любого возраста). Известно, что в некоторых случаях ястребы-тетеревятники убивали родителей уральских сов, по-видимому, совершенно вне конкуренции (несмотря на разные основные источники пищи) и откладывали яйца в гнездо, в котором все еще находились яйца сов. [ 4 ] [ 30 ] [ 146 ] Хищники, которые, как было отмечено, похищали молодых уральских сов, обычно примерно в молодом возрасте или вскоре после расселения (особенно тех, которые использовались при реинтродукции ), также включали рыжих лисиц ( Vulpes vulpes ), европейских сосновых куниц ( Martes martes ) и больших серых сов . [ 4 ] [ 146 ] [ 152 ] [ 153 ] В Приморском крае в азиатские барсуки ( Meles leucurus ) и енотовидные собаки ( Nyctereutes procyonoides ). качестве потенциальных или вероятных хищников гнезд уральских сов также упоминаются [ 117 ] В одном зарегистрированном случае было замечено, что уральская сова, которая, по-видимому, пыталась охотиться на некоренную американскую норку ( Neogale vison ), проиграла конфронтацию, когда норка перевернула ситуацию, очевидно одолев, убив и съев сову. [ 154 ] Хотя случаи нападения хищников на уральских сов широко известны, относительное количество таких случаев очень невелико и единично. К югу от Арктики его можно считать конкурентом большой серой совы как вида совы, который является вторым наименее уязвимым для хищников видом после высших хищников -филинов. [ 4 ] [ 30 ] [ 146 ] Уральская неясыть сама по себе является довольно грозным хищником более мелких видов сов, хотя и не столь плодовитым убийцей, как филины и северные ястребы-тетеревятники. Известно, что среди сов, на которых охотятся уральские совы, - индийская сплюшка ( Otus lettia ), сплюшка с воротником ( Otus bakkamoena ), северные ястребиные совы, серо-коричневые совы, евразийские карликовые совы, бореальные совы и ушастые совы . [ 4 ] [ 7 ] [ 30 ] [ 85 ] [ 117 ] [ 138 ] [ 146 ] Дневные хищники также иногда уязвимы для хищников со стороны уральских сов, в том числе серолицых канюков ( Butastur indicus ), перепелятника ( Accipiter nisus ), обыкновенной пустельги ( Falcotinniculus ) и даже явно взрослых северных ястребов-тетеревятников. Таким образом, уральская сова, в сочетании с очень низким известным уровнем естественных хищников, по-видимому, занимает такое же положение во внутригильдейной хищнической иерархии, что и ястреб-тетеревятник. [ 85 ] [ 84 ] [ 114 ] [ 117 ] В Финляндии как более мелкие совы, так и дневные хищники, такие как перепелятники и даже канюки, по-видимому, избегают гнездиться рядом с уральскими совами. [ 155 ] [ 156 ] Кроме того, известно, что уральские совы успешно отгоняли активно гнездящихся ястребов-тетеревятников, а также европейских осоев ( Pernis apivorus ), обыкновенных канюков ( Buteobuteo ) и черных коршунов ( Milvus migrans ), чтобы завладеть гнездами. [ 30 ] [ 155 ] [ 157 ] Несмотря на их потенциальное хищничество на более мелких мезохищников , когда гнездовые ящики для европейских мухоловок-пеструшек ( Ficedula hypoleuca ) были экспериментально размещены в районе гнезд уральской совы, продуктивность снизилась, и это, возможно, связано с тем, что мезохищников часто привлекала соответствующая пищевая база сов. . [ 158 ] Напротив, экспериментальные наземные гнезда, выложенные исследователями из случайных яиц домашней птицы в центральной Финляндии, оказались случайно защищены яростным присутствием уральских сов. На финской земле, расположенной почти непосредственно под уральским гнездом, ни на кого вообще не охотились, в то время как на территории, расположенной на расстоянии 100 м (330 футов), 8,3% подверглись нападениям хищников, а те, кто находился в пределах 200–400 м (660–1310 футов), испытали средний уровень хищничества 58,3%. На большом расстоянии, на высоте 500–900 м (1600–3000 футов), все наземные гнезда были истреблены хищниками. [ 159 ]
Разведение
[ редактировать ]Парное размещение
[ редактировать ]Уральская неясыть обычно создает пары на всю жизнь и сохраняет территорию в течение нескольких лет. Было обнаружено, что удержание партнера в Фенноскандии в среднем составляет 98–100% у самцов и 90–95% у самок, что делает их одними из самых моногамных хищных птиц любого отряда (или, по крайней мере, хорошо изученных видов хищных птиц). [ 160 ] [ 161 ] [ 162 ] Территории уральских неясыть в среднем примерно в 3 раза больше, чем территории неясыти. В Швеции на территории протяженностью 150 000 км встречается около 3000 пар уралов. 2 (58 000 квадратных миль). [ 4 ] [ 5 ] В Финляндии гнезда обычно находятся на расстоянии от 2 до 4 км (1,2–2,5 мили) друг от друга в дикой природе. [ 8 ] По финским данным, уральская неясыть занимала второе место по количеству найденных гнезд среди видов сов с 1985 по 1989 год: 901 гнездо уступало бореальной сове с 2265 гнездами. [ 161 ] В Хорватии в разных районах в среднем на 10 км приходилось от 1,1 до 5,4 пар. 2 (3,9 квадратных миль), при этом примерно 700-1000 гнездящихся пар осталось примерно на 37% территории Хорватии, которая все еще покрыта лесами. Пик хорватской популяции был зафиксирован в национальном парке «Плитвицкие озера» , преимущественно в смешанном елово - буковом горном лесу, где обитало около 38 гнездящихся пар. [ 6 ] Размножение в Словении происходит на средней высоте 850 м (2790 футов), причем высота над уровнем моря менее важна, чем среда обитания. В Словении использовалось множество типов леса, но елово - буковый чаще всего использовался смешанный лес. Самая высокая плотность в Словении составляла около 2-3 пар на 10 км. 2 (3,9 квадратных миль) для горы Крым и около 4-5 пар на 10 км. 2 (3,9 квадратных миль) для Снежника . [ 163 ] По оценкам, в 2016 году в Словении имеется 400–700 пар. [ 164 ] Было проведено исследование вида в Силезских Бескидах , популяция которого может отражать недавнее увеличение популяции. При трех попытках размножения плотность оценивалась в 0,6-0,9 пары на 10 км. 2 (3,9 квадратных миль), что в 3–10 раз ниже, чем в других близлежащих хребтах Восточной Европы, таких как Низкие Бескиды и Бещады . [ 165 ] Низкая плотность населения была обнаружена в Пенинском национальном парке , Польша : 0,9-1,3 на 10 км. 2 (3,9 квадратных миль). [ 166 ] Средняя плотность на польской Краковско-Ченстоховской возвышенности составляла 1 пару на 10 км. 2 (3,9 квадратных миль). [ 167 ] В северной части Моравии , Чехия , на рубеже XXI века было зарегистрировано 3-5 пар, что означает почти уверенный прирост популяции. [ 168 ] [ 169 ] отмечен рост популяции В районе Расточья на Украине , несмотря на то, что лес является смешанным, а не предпочтительной буковой доминантой подвида, сейчас до 1,7 пар на 10 км. 2 (3,9 квадратных миль). [ 170 ] Там, где были построены гнезда для уральских сов в Самарской области, показано, как совы поселились в ящиках на среднем расстоянии 1,5 км (0,93 мили); при этом среднее расстояние всех установленных коробок составляло 1,11 км (0,69 мили). [ 171 ]
Характеристики гнезда
[ редактировать ]

Потенциальные места гнездования включают большие естественные отверстия в деревьях, полости, оставленные большими отломанными ветвями, полые стволы с отломанными навесами (или «трубы дымоходов»), трещины или отверстия в скалах или между камнями и дыры в зданиях . [ 4 ] [ 5 ] Скалы деревьев и пни, используемые преимущественно в Центральной и Восточной Европе, довольно часто представляют собой березу обыкновенную ( Fagus sylvatica ) или иногда дуб обыкновенный ( Quercus robur ). [ 30 ] [ 164 ] Также уральские совы могут регулярно использовать гнезда из палочек более крупных птиц, таких как различные ястребы , в частности гнезда, построенные ястребами-тетеревятниками и канюками, а также гнезда черного аиста ( Ciconia nigra ), гнезда ворона ( Corvus corax ) и беличьих дреев , хотя дреи и гнезда более мелких птиц, таких как перепелятники и вороны, могут подвергаться риску регулярного разрушения, поскольку они могут быть слишком маленькими. и, возможно, хлипко построен. [ 4 ] [ 5 ] Весьма необычное с точки зрения регионального ареала место гнездования было зарегистрировано в Словакии , на Выходословенской равнине , в низменной пойме , внутри старого гнезда канюка. [ 172 ] Гнездо может располагаться вблизи небольших городов, как это было зарегистрировано на юге Сахалина , при условии, что в регионе имеется соответствующая среда обитания и доступ к добыче. [ 173 ] В одном случае в Норвегии уральская сова использовала отверстие в обыкновенной осине ( Populus tremula ) на том же дереве, тогда как обыкновенный крохаль ( Mergus merganser ) использовал другую нору. [ 174 ] В некоторых частях своего ареала они могут использовать гнездовые ящики, причем тенденция размещения гнездовых ящиков началась и, возможно, наиболее упорно используется в Фенноскандии . [ 5 ] Этот вид предпочитает большие гнездовые ящики с отверстием около 16 см (6,3 дюйма) в диаметре. [ 5 ] В других странах, как известно, использовалось около 29% гнездовых ящиков, установленных в Словении. [ 164 ] России В Алтайском крае , в районе реки Бия , в период с 2010 по 2012 год было использовано 15,2-48,9% построенных гнездовых ящиков с годовыми колебаниями, объясняемыми циклами первичных источников питания. [ 9 ] В Самарской области из 74 гнездовых ящиков, установленных на 4 исследуемых участках, к 2009 г. 41,9% использовались уральскими совами (около 14,9% - различными другими животными). В Самаре около 20,6% пар уральских сов в исследуемом блоке по-прежнему использовали естественные места гнездования, несмотря на наличие гнездовых ящиков. [ 171 ] Рядом с предыдущим участком, в лесных массивах под Бийском Алтайского края , выявлено 8 территорий, все 6 гнезд найдены в гнездах, построенных ястребиными. [ 9 ] Использование 15 гнездовых ящиков в Академгородке , Россия, за три года исследования сильно варьировалось, предположительно, в зависимости от циклов популяции добычи, при этом ежегодно использовалось от 0% до 50%. [ 175 ] Побочная польза была показана для нескольких видов жуков , а иногда и для других насекомых, которые гнездились в субстрате гнездовий уральской совы, даже в районах, где беспозвоночные редки или находятся под угрозой исчезновения. [ 176 ] [ 177 ] [ 178 ]
Из 250 гнезд, зарегистрированных в Финляндии за почти столетие, большая часть приходится на пни (34,3%), за ними следуют птичьи гнезда (28,5%), гнездовые ящики (23,1%), дупла деревьев (11,35%) и меньшее количество гнезд на земле или на камнях. лицо или здание. 53% финских пней были из ели и 38% из осин , высотой от 1,2 до 10 м (от 3,9 до 32,8 футов) и в среднем 4,5 м (15 футов). Пням отдавали предпочтение в северной и центральной части Финляндии. 67% финских полых гнезд были отломанными ветками берез , хотя черного дятла ( Dryocopus martius иногда использовались также сгнившие и расширившиеся норы ), при этом высота полых гнезд составляла от 1,5 до 12 м (от 4,9 до 39,4 футов). В южной части Финляндии предпочтение отдавалось кариесу. После 1960 года значение гнездовых ящиков возросло, и к 1969 году 50% зарегистрированных случаев использования гнезд во всей Финляндии приходилось на гнездовые ящики. [ 179 ] В Словении 56,2% зарегистрированных гнезд находились в дуплах деревьев , еще около 20% - на пнях, на высоте от 1 до 10 м (от 3,3 до 32,8 футов) над землей (в среднем 5,2 м (17 футов)). Реже словенские совы гнездятся в старых гнездах хищных птиц (16-19%). [ 163 ] [ 164 ] В Словакии леса с преобладанием бука выбирались примерно в 74% случаев. Среди 27 словацких гнезд 59,3% приходилось на буки обыкновенные, 14,8% на пихты , 11,1% на ели и 14,8% на другие лиственные деревья. [ 140 ] Гнезда на пнях были зарегистрированы дважды на Хоккайдо, но в остальном японские уральские совы, по-видимому, редко используют такие места для гнезд, отдавая предпочтение дуплам деревьев, птичьим гнездам и гнездовым ящикам, при этом мало свидетельств того, что они явно отдают предпочтение какому-либо одному типу мест гнездования. [ 180 ] [ 181 ] Даже в пиковые годы полевок некоторые части могут быть неспособны к размножению из-за отсутствия подходящих мест для гнезд. [ 8 ] Помимо не поддающейся количественной оценке угрозы глобального потепления, главной угрозой для этого вида является потеря старых лесов с мертвыми деревьями с обширными полостями. Исторически это особенно распространено при лесопользовании , и практика департаментов лесного хозяйства часто отдавала предпочтение серым неясытьм и наносила ущерб популяциям уральских сов. [ 4 ] [ 8 ] [ 182 ] Однако в Хедмарке , Норвегия , установка 80 гнездовых ящиков не увеличила популяцию уральской совы в стране: использовались только 4 ящика. Таким образом, отсутствие успеха норвежских популяций, по-видимому, не связано с отсутствием мест гнездования, а, как предполагается, в большей степени коррелирует с плохой популяцией их добычи. [ 183 ] Предыдущие исследования проводились в центральной и восточной Норвегии, несколько западнее Норвегии, чем считалось ранее, но не в широком диапазоне, при этом размер кладки был меньше, в среднем 2,85, в Скандинавии по сравнению с 2,93 в Швеции, 3 в южной Финляндии и 3,92. в северной Финляндии. [ 184 ]
Яйца и молодь
[ редактировать ]
Размер кладки обычно составляет 3-4 человека (редко от 1 до 6), яйца чисто-белые и довольно округлые. [ 5 ] Средние сроки откладки яиц в Фенноскандии приходится на середину марта и конец апреля. [ 185 ] В горной Словении, что несколько удивительно, это происходит немного позже, в конце марта - начале июня. [ 164 ] составил 2,93 Средний размер кладки в Швеции . [ 105 ] Средний размер кладки в Финляндии составлял 2,24, но мог колебаться в среднем от 2,08 до 3,98 в плохие и хорошие годы для добычи полевок. [ 185 ] [ 186 ] [ 187 ] Средний размер кладки в Словении составляет около 3,3. [ 164 ] Другое словенское исследование показало довольно меньший средний размер кладки - 2,4, а также доказательства того, что кладка сокращалась из-за отсутствия доступа к пище в начале сезона размножения. [ 188 ] Размер кладки в небольшой выборке из Нижнего Новгорода, Россия, составил в среднем 3,6. [ 189 ] Средний размер кладки в гнездовых ящиках Самарской области составил 2,4. [ 171 ] Размеры яиц обычно составляют от 46,5 до 52,3 мм (1,83 и 2,06 дюйма) в высоту и от 39 до 44 мм (от 1,5 до 1,7 дюйма) в диаметре, а вес свежих яиц в среднем составляет около 47 г (1,7 унции). По данным исследований, проведенных на юге Финляндии, среди 59 изученных самок размер яиц варьируется на 22,4% в течение года цикла, а самые большие яйца примерно в два раза превышают массу самых маленьких, что является очень значительным отклонением. [ 190 ] Яйца откладываются прямо на дно гнездовой поверхности примерно с двухдневным интервалом. [ 5 ] Насиживают только самки, обычно начиная с первого яйца, и все время их кормит партнер. [ 5 ] У 108 самок уральских сов из Финляндии начало инкубации варьировалось индивидуально, при этом синхронное вылупление оказалось неблагоприятным для общей продуктивности. Были обнаружены доказательства того, что самки повторяли время начала инкубации ежегодно, было умеренно сильным (повторяемость 26%), поэтому это может быть развитой чертой. [ 191 ] Инкубация длится от 28 до 35 дней и в среднем примерно на 6 дней дольше, чем инкубационный период коричневой совы. [ 5 ] [ 192 ] Птенцы ломаются одновременно с откладкой яиц (около 2 дней), самки остаются до оперения. [ 5 ] Пушистый птенец белый; на стадии, когда они обычно покидают гнездо (или на мезоптильной стадии), пух бледно-грязно-беловатый, с серовато-коричневыми полосами на голове, затылке, спине и нижней части тела. [ 5 ] Птенцы покидают гнездо примерно в 35-дневном возрасте и могут летать в 45-дневном возрасте. Их кормят и о них заботятся еще около двух месяцев после выхода из гнезда.

Родительское поведение и характеристики
[ редактировать ]Молодые уральские совы официально достигают половой зрелости на следующий год после обретения независимости. [ 5 ] Однако на юге Финляндии было обнаружено, что самки, впервые гнездящиеся, обычно не пытались гнездиться, если это был плохой год для полевок, а самый большой процент (около 40%) впервые начал попытки размножения в промежуточные годы (между плохими и хорошими годами для полевок). ), чего часто не происходило, пока им не исполнилось 3–4 лет. [ 193 ] Тот же типичный возраст (3–4 года) был обнаружен в центральной Швеции, несмотря на то, что самки часто находились на территории в течение первого года жизни. [ 182 ] Изучение линьки и изнашивания перьев показало, что некоторые самки уральских сов размножаются на 2-й или 3-й год жизни, но большинство из них не размножаются до 4-го или 5-го года жизни. [ 194 ] Напротив, финское исследование показало, что для уральских сов обоих полов средний возраст начала размножения составляет 2,9 года, а средняя продолжительность жизни при размножении - 3,3 года. [ 195 ] Более того, в Финляндии было показано, что самки, начавшие размножаться в возрасте 2 или 3 лет, имели более высокую репродуктивную «приспособленность», чем самки, начавшие размножаться в 4 года и старше. Самки откладывают яйца в Финляндии в среднем 2,7 раза за свою жизнь и производят на свет в среднем 6,7 птенцов за репродуктивную жизнь (при этом наиболее продуктивные самки производят до 33 птенцов). [ 196 ] Пик активности насиживания, обычно с 9–14 посещениями в день, в финских исследованиях приходится на поздний вечер весной и летом, когда наблюдается уход за молодняком, с более слабым пиком активности ранним утром, оба раза приурочены к пику активности добычи. [ 23 ] Хорошо известно, что мать уральских сов грозно защищает свой выводок. [ 4 ] В Финляндии повторяемость защитного поведения самок при гнездовании оценивалась в 52,4%, начиная с лая и бегства, затем пролетая в направлении предполагаемой угрозы и заканчивая атаками и мощными ударами. Согласно этому исследованию, те самки, которые отложили яйца раньше, в среднем защищали свое гнездо более энергично, чем те, кто отложил кладку позже. [ 195 ] Сила и свирепость нападения привели к его общему названию на шведском языке ( Slaguggla или «нападающая сова»), и их атаки потенциально могут нанести тяжкий физический вред даже такому большому животному, как человек. По крайней мере, в некоторых случаях люди теряют один глаз или ослепляют из -за сломанных глазниц или глазных яблок, извлеченных в результате нападения уральской совы. [ 30 ] [ 159 ] [ 195 ] [ 197 ] Из-за ее нападений уровень хищничества уральской совы часто бывает исключительно низким. [ 195 ] Считалось, что частичная линька перьев самок не имела большого влияния на ее способность к насиживанию, поскольку она в основном была приурочена к периодам, когда самка (и ее выводок потомства) почти полностью кормились самцом пары. [ 198 ]
Успех разведения
[ редактировать ]По данным исследования, проведенного в южной Финляндии, выводки, имеющие более трех птенцов, иногда подвергаются сиблициду , но в остальном случаи сиблицида редки. [ 186 ] Большие кладки, по-видимому, являются страховым поведением, поскольку авторы показали, что у выводков из 4 или около того младший брат с большей вероятностью умрет раньше или позже до обретения независимости. [ 191 ] Продуктивность в значительной степени связана с доступом к добыче, при этом эффект, наиболее изученный в Фенноскандии, поскольку резкие трехлетние колебания численности добычи полевок, по-видимому, вызывают большие различия в продуктивности. [ 30 ] Коэффициент рождаемости может варьироваться от нуля до 2,9 в разные годы в зависимости от количества жертв в Швеции, при этом средний показатель составляет 1,03–1,12 птенцов на пару и 2,68 птенцов на успешную пару. Однако в Швеции было подсчитано, что в среднем 62% сов не доживают до первого года жизни, по сравнению с годовой смертностью взрослых самок, которая оценивается в 10,5%. [ 105 ] [ 182 ] Аналогичным образом, на юге Финляндии от 29 до 58 гнездящихся пар ежегодно размножаются в одном и том же районе исследования и могут производить от 1,17 до 3,21 птенцов в зависимости от цикла добычи (3,5 средних птенцов на успешную попытку в годы высокой полевки против 1,79 на успешное гнездо в годы низкой полевки). год полевки). Финское исследование показало, что 21,7% всех кладок не вылупились, а 4,3% полностью погибли после вылупления. [ 185 ] [ 199 ] Выводимость на юге Финляндии составляет в среднем 87% за все годы. [ 190 ] В более широком масштабе с 2009 по 2011 год в Финляндии было зарегистрировано от 422 до 1710 территорий, при этом количество успешных гнезд ежегодно колеблется от 168 до 1341. [ 199 ] Популяционные тенденции более или менее повторяют тенденции финских серых неясытей, но размер выводка желтовато-коричневых неяс в среднем на 0,68 больше, чем у уральских. [ 199 ] Перспективы размножения, как правило, невелики в низшую точку трехлетнего цикла добычи полевок Урала на юге Финляндии, но, как это ни парадоксально, когда за годом высокой полевки следует низкий год, количество попыток кладок выше, но скорость отказ гнезда выше. [ 10 ] Например, в рамках финского исследования 144 яйца были обнаружены в 63 гнездах во время фазы низкого уровня полевок, в фазе роста - 1212 из 338 гнезд и в пиковой фазе - 1259 яиц из 345 гнезд. Однако это исследование показало, что низкое пополнение из кладок было обнаружено как в годы роста, так и в годы пика, а осложнения и индивидуальные вариации каждой пары могут вызывать вариации, которые не учитываются предыдущими теориями. [ 200 ] Успех размножения на юге Финляндии также был обусловлен возрастом матери уральской совы: самки в возрасте 3–5 лет были менее продуктивными, чем самки в возрасте от 6 до 9 лет, а самки старше 10 лет имели тенденцию к размножению. чтобы нести более крупные и округлые яйца. [ 186 ] В Финляндии пары, получавшие дополнительное питание, гнездятся на 1 неделю раньше и производят на 0,6 больше яиц, чем те, которые этого не делали, поэтому ограничение еды, по-видимому, действительно влияет на продуктивность вида. [ 201 ] Самцов в Финляндии было немного больше среди оперившихся детенышей (56%), но выживаемость обоих полов была практически одинаковой, а масса тела у обоих полов в хорошие годы у полевок была в среднем примерно на 6% больше. [ 202 ] Исследования 274 взрослых финских самок показали, что 18% из них произвели на свет около половины птенцов. [ 196 ] В результате попыток размножения в Нижнем Новгороде в среднем из гнезда вышло 3,1 детеныша. [ 189 ]
находится южнее, чем многие вышеупомянутые исследования (например, из Фенноскандии), Несмотря на то, что Эстония в течение циклов популяции добычи одновременно наблюдалась сильно варьирующаяся степень успеха размножения. [ 107 ] В Словении около 80% попыток размножения удается произвести хотя бы одного птенца. [ 164 ] Другое словенское исследование показало, что общий успех размножения почти на 5% выше из-за пика численности съедобных сонь в конце лета (июль или позже). [ 139 ] Было показано, что 7 гнезд в Подкарпатском воеводстве , Польша , способны произвести в среднем 2,8 птенцов в хороший год, но в плохой год для грызунов гнезда найти не удалось. [ 203 ] Результаты показали, что родительские уральские совы, особенно взрослые самки, способны корректировать свой цикл размножения, чтобы максимизировать выживаемость своих детенышей, учитывая опыт, накопленный в течение их относительно долгой жизни, и относительно успешно смягчают такие проблемы, как паразитизм и заражения гнезда. коробки, если они используются. [ 204 ] Молодь после расселения из Венского леса в Австрии отслеживалась по радио, и ее среднее расстояние расселения от места выпуска составило 8778 м (28799 футов), а уровень смертности составил около 23%. [ 152 ]
Статус
[ редактировать ]Уральская неясыть не является густонаселенной птицей, но на местном уровне может быть нередкой. [ 5 ] , По оценкам МСОП в дикой природе во всем мире живет от 350 000 до 1 200 000 особей. [ 1 ] Наибольшее сокращение в новейшей истории было зарегистрировано в районах, где из лесов были вывезены дуплистые и сломанные деревья. [ 5 ] [ 205 ] В Эстонии в управляемых лесах почти всегда меньше уральских сов, чем в нетронутых лесах, из-за меньшего количества коряг и других естественных полостей, которые можно использовать. [ 167 ] [ 206 ] Однако общие тенденции положительны для большинства популяций европейской уральской совы. [ 1 ] [ 140 ] Возведение гнездовых ящиков привело к увеличению популяции в некоторых частях ареала, особенно в Финляндии. [ 5 ] [ 179 ] В Восточной Европе это один из наиболее устойчивых видов сов, хотя в целом он гораздо менее многочисленный, чем некоторые (например, желто-коричневые, ушастые и маленькие совы ( Athene noctua )). [ 207 ] В недавней истории в Центральной и Восточной Европе было обнаружено несколько увеличений и расширений популяций уральских сов, синхронно с другими совами, считающимися бореальными видами (т.е. большой серой , бореальной , евразийским пигмеем ). Предыдущие данные указывали на основные популяции уральских сов в 1980-х годах в западных Карпатах (по оценкам, 1000 пар) и северной Беларуси (50-100 пар). К 1990-м годам численность выросла до 1000-1500 пар в западных Карпатах и до 220-1350 пар в северной Беларуси. К 2005 году численность достигла 3500 пар в Карпатах и резко возросла до 2700-4300 пар в Беларуси. В Чешской Республике, отчасти из-за целенаправленной реинтродукции, численность выросла с 1–5 пар в 1985–89 годах до 25–40 пар к 2001–2003 годам. На отдельных участках юго-восточной Польши, Беларуси и Латвии плотность составила 1-2 пары на 100 км. 2 (39 квадратных миль) до 10 пар на 100 км 2 (39 квадратных миль). В этих трех странах плотность северного населения сейчас гораздо выше, чем южная, т.е. 5–8,1 пары на 100 км². 2 (39 кв. миль) на севере до 3,1-3,6 пар на 100 км. 2 (39 квадратных миль) на юге. [ 208 ] [ 209 ] [ 210 ] Расширение ареала уральских сов отмечено на западе Украины (в районе биосферного заповедника «Розточье» и Яворовского национального парка ). в 2005-07 гг. до 1,7-2 пары на 10 км. 2 (3,9 квадратных миль), тогда как в прошлом (т.е. до 1990-х годов) этот вид был редким бродягой на этой территории. Плотность украинского населения выше, чем, казалось бы, в большинстве стран Скандинавии и Беларуси, но ниже, чем в юго-западной Польше и Словении; хотя неизвестно, представляет ли это увеличение численности населения или просто его перемещение, и неизвестно, как это связано с лесным хозяйством. [ 211 ] В некоторых частях Словакии , таких как Сланские Врхи , Вигорлат и Ондавское нагорье, плотность пар может достигать пары на квадратный километр, что, возможно, является самым высоким из известных видов. [ 140 ] 400-500 словенских пар с 1973 по 1994 год, а по состоянию на 2006 год оценивается в 1400-2500 пар. В Оравском регионе Словакии население могло увеличиться в пять раз за вышеуказанные годы. [ 140 ] В то время как численность многих видов сов ( филин , ушастая , бореальная ), по-видимому, в целом сократилась в период 1982–2007 годов в Финляндии, численность уральских сов, напротив, увеличилась примерно на 1% (исключение из этих оценок было слишком трудным для анализа). северные кочевые виды сов). [ 212 ] Во всех отношениях, кроме количества окольцованных птенцов (в котором он также уступает евразийскому пигмею), было замечено, что уральская сова является второй по распространенности обнаруженной гнездящейся совой после бореальной совы в Финляндии: обнаружено 2545 территорий, обнаружено 1786 гнезд и 4722 гнезда. птенцы окольцованы. [ 213 ] Увеличение численности населения зафиксировано в Московской области, где сохранились высокие насаждения, несмотря на довольно развитую окружающую среду. [ 214 ]

Иногда уральские совы уязвимы для налетов на искусственные объекты. В большинстве частей ареала они менее уязвимы, чем многие другие крупные хищные птицы (отчасти из-за того, что они предпочитают отдаленные леса), но, безусловно, некоторые из них могут быть признаны таковыми. Многие такие случаи смертности происходят из-за столкновений с проводами и поражений электрическим током , которые, вероятно, увеличиваются, особенно по мере того, как население увеличивается и перемещается в районы, расположенные ближе к человеческому жилью. [ 215 ] [ 216 ] [ 217 ] Другие случаи гибели людей в результате столкновений, например, со стеклянными зданиями и, в широком смысле, с различными автомобилями, также потенциально могут расти. [ 218 ] [ 219 ] [ 220 ] Хотя исторически уральские совы подвергались некоторой степени преследований , они были избавлены от худших из них, возможно, благодаря тому, что они гнездились в отдаленных лесах и, возможно, в целом были менее хищными по отношению к мелкой домашней птице, дичи и т.п., чем крупные хищные птицы, такие как евразийские филины. , беркуты и северные ястребы-тетеревятники, все из которых подверглись жестоким преследованиям и, таким образом, сократились. [ 221 ] [ 222 ] [ 223 ] Как и другие дикие птицы, уральские совы могут быть в некоторой степени уязвимы к смертности из-за болезней и инфекций, но они вряд ли поставят под угрозу популяцию в целом. [ 30 ] случай бактериальной инфекционной болезни Туляремия У уральской совы выявлен вирус Усуту . , а у одной птицы - [ 224 ] [ 225 ] меньшее, но все еще присутствующее количество скребней и круглых червей . У 71,4% из 14 диких уральских сов в Японии были кровяные паразиты, в то время как в известных европейских данных было обнаружено [ 226 ] [ 227 ] [ 228 ] Многие японские уральские совы также оказались уязвимы к укусам вшей . [ 229 ]
Реинтродукция
[ редактировать ]Реинтродукция видов проводилась в некоторых частях Европы. Реинтродукция уральской совы, безусловно, лучше всего документирована в Богемском лесу , который простирается между странами и регионами Баварии в Германии , Чехии и верхней Австрии . Ранее вид здесь вымер к 1926 г. (к 1910 г. на австрийской стороне). Создание племенного поголовья в неволе произошло в период с 1972 по 2005 год (происхождение из 7 разных стран и смесь двух основных европейских подвидов). Для этой программы разведения изначально было выпущено 212 молодых уральских неясытей. В ходе исследования были произведены экспериментальные интродукции в гнезда серой неясыти, хотя это потенциально подвергало их риску гибридизации. В остальном родословная состояла из относительно недавно появившихся уральских сов. Было показано, что как желто-коричневые, так и уральские совы способны успешно выращивать молодые уральские выводки. Корм также предлагался парам в близлежащих загонах для выпуска и было построено 60 гнездовых ящиков, чтобы компенсировать нехватку мест для гнезд, особенно в районах вторичный лес . 33 уральских совы были обнаружены мертвыми, а еще 4 были ослаблены или ранены до такой степени, что не могли продолжать жить в дикой природе. Большая часть смертей произошла из-за поражения электрическим током или попадания под машины, но некоторые были застрелены незаконно. Первое дикое размножение уральской пары в Богемском лесу произошло в 1985 году, но первое успешное размножение произошло только в 1989 году (пара произвела 4 потомства). В период с 1981 по 2005 год всего было зарегистрировано 49 выводков, 31 из которых был успешным, при этом было произведено 59 детенышей (в среднем 1,3 на все попытки, 1,9 на успешную пару). К концу исследований было создано не менее 6 пар (возможно 5-10 гнездящихся пар), при этом продуктивность в пределах леса оценивается в 10 пар. Было сочтено, что для самоподдерживающейся размножающейся популяции необходимо не менее 30 пар на общей территории в пределах коридоров, ведущих в Богемский лес. Таким образом, в период с 1995 по 2006 год в близлежащий национальный парк Шумава было завезено 87 птиц , сейчас там обосновались примерно 2-3 гнездящиеся пары. Все еще неопределенная пилотная программа в Мюльфиртель , Австрия, возможно, тоже выпустил пару, а мог и не родить. В 2001 году из двух попыток реинтродукции в Австрии обе потерпели неудачу. Судя по всей реинтродукции в Баварии, было сказано, что в период с 1972 по 2014 год совы произвели в общей сложности 204 потомка, хотя многие из них, возможно, не выжили. [ 11 ] [ 230 ] [ 231 ] [ 232 ] [ 233 ] [ 234 ] Более успешной, чем австрийская часть реинтродукции баварских лесов, была реинтродукция в других частях Австрии, а именно в Венском лесу . В рамках этого проекта в период с 2009 по 2013 год было выпущено 67 молодых сов. Было запланировано использовать сеть скворечников из 127 ящиков, а также присутствовал один из крупнейших в Европе насаждений бука. В Венском лесу выживаемость была высокой - около 70,5%. К 2011–2012 гг. 10 пар попытались гнездиться, установив участки обитания в среднем около 300 га (740 акров), и на каждую успешную пару родилось 3,1 птенца. [ 153 ] [ 235 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л м н тот BirdLife International (2016). « Стрикс ураленсис » . Красный список исчезающих видов МСОП . 2016 : e.T22689108A93218506. doi : 10.2305/IUCN.UK.2016-3.RLTS.T22689108A93218506.en . Проверено 12 ноября 2021 г.
- ^ «Приложения | СИТЕС» . сайт цитирует . Проверено 14 января 2022 г.
- ^ Склетер, Польша (1879). Замечания о номенклатуре британских сов и о расположении орденских полос . Ибис, 21(3), 346–352.
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а к ап ак с как в В из хорошо топор является тот нет бб до нашей эры др. быть парень бг чб с минет БК с Вус, К.Х. (1988). Совы Северного полушария . Массачусетский технологический институт Пресс, ISBN 0262220350 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а к ап ак с как в В из хорошо топор является тот нет бб до нашей эры др. быть парень бг чб с минет БК с бм млрд быть б.п. БК бр Король, Клаус; Вейк, Фридхельм (2008). Совы мира (2-е изд.). Лондон: Кристофер Хелм. ISBN 9781408108840 .
- ^ Jump up to: а б Тутиш В., Радович Д., Чикович Д., Баришич С. и Краль Дж. (2009). Распространение, плотность и взаимоотношения уральской совы Strix uralensis macroura в Хорватии . Ардеа, 97(4), 563-571.
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р Обуч Дж., Данко Ш., Михок Дж., Караска Д. и Шимак Л. (2014). Рацион уральской совы (Strix uralensis) в Словакии . Словацкий журнал Raptor, 7, 59–71.
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но Хьюм, Р. (1991). Совы мира . Бегущая пресса, Филадельфия.
- ^ Jump up to: а б с Важов С.В., Бахтин Р.Ф. и Важов В.М. (2016). Об использовании гнездовий для изучения экологии Strix uralensis . Международный журнал прикладных и фундаментальных исследований, 333: 498-498.
- ^ Jump up to: а б Броммер Дж. Э., Пиетияйнен Х. и Колунен Х. (2002). Размножение и выживание в изменчивой среде обитания: уральские совы (Strix uralensis) и трехлетний цикл полевок . Аук, 119 (2), 544–550.
- ^ Jump up to: а б с Шерзингер, В. (2006). Восстановление встречаемости уральской совы Strix uralensis в Чешском лесу . Журнал орнитологов Баварии и Баден-Вюртемберга, 45 (2/3).
- ^ Jump up to: а б с д Бразилия, М. (2018). Птицы Японии . Издательство Блумсбери.
- ^ Jump up to: а б Совы мира: Фотопутеводитель Микколы, H. Firefly Books (2012), ISBN 9781770851368
- ^ Сильвен, А. (1907). Охота в нашей стране . Издательство П. А. Норстедт и сыновья.
- ^ Jump up to: а б Кивеля, Дж. (2011). Изучение и отлов гнездящихся уральских сов в западной части Финляндии . Сохранение хищников, (21).
- ^ Jump up to: а б с д и ж г Справочник CRC по массе тела птиц , автор Джон Б. Даннинг-младший (редактор). ЦРК Пресс (1992), ISBN 978-0-8493-4258-5 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот Вейк, Фридхельм (2007). Совы (Strigiformes): аннотированный и иллюстрированный контрольный список . Спрингер. ISBN 978-3-540-39567-6 .
- ^ Jump up to: а б с д и Холт Д.В., Беркли Р., Деппе К., Энрикес Роша П., Петерсен Дж.Л., Рангел Салазар Дж.Л., Сегарс К.П., Вуд К.Л. и Маркс Дж.С. (2019). Типичные совы . В: дель Ойо Дж., Эллиотт А., Саргатал Дж., Кристи Д.А. и де Хуана Э. (ред.). Справочник птиц мира живых . Lynx Editions, Барселона.
- ^ Jump up to: а б с Шерзингер В., Фанг Ю., Сан Ю.Х. и Клаус С. (2014). Пересмотренное описание совы Пере Давида Strix davidi на основе полевых наблюдений в Центральном Китае . Орнитол. Анз., 53: 54–93.
- ^ Jump up to: а б с д и ж г час я дж к л м н Момияма, Т.Т. (1928). Новые и известные формы уральской совы (Strix uralensis) из юго-восточной Сибири, Маньчжурии, Кореи, Сахалина и Японии . Аука, 177–185.
- ^ Jump up to: а б с Бутурлин, С.А. (1907). Палеарктика формируется раньше Syrnium Savign . Журнал орнитологии, 55: 332–336.
- ^ Jump up to: а б с д Лундберг, А. (1980). Вокализации и брачное питание уральской совы Strix uralensis . Орнис Скандинавица, 65–70.
- ^ Jump up to: а б с д и Корпимяки Э. и Хухтала К. (1986). Частота посещения гнезд и характер активности уральских сов Strix uralensis . Орнис Фенника, 63(2), 42–46.
- ^ Отто-Спрунк, А. (1967). Прыгаемость уральской совы (Strix uralensis) . Орнис Фенн. 44:78.
- ^ Линдблад, Дж. (1967). На совиных полях . Стокгольм.
- ^ Холмберг, Т. (1974). Исследование сипухи Strix uralensis small . Вр Фдгельв, 33, 140-146.
- ^ Jump up to: а б Шерзингер, В. (1980). К этологии размножения и развития молоди уральской совы (Strix uralensis) в сравнении с неясытью (Strix aluco) . Бонн. Зоол. Монгр, 15.
- ^ Jump up to: а б Келсо, Л. (1940). Изменение наружного ушного отверстия у Strigidae . Бюллетень Уилсона, 24–29.
- ^ Jump up to: а б с д Вус, К.Х. (1964). Лесные совы родов Strix и Ciccaba . Зоологические коммуникации, 39(46), 471-478.
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и Миккола, Х. (1983). Совы Европы . Т. и А.Д. Пойзер.
- ^ Jump up to: а б с Уордхау, А.А. (1983). Совы Британии и Европы . Бланфорд.
- ^ Jump up to: а б с д и ж г час Дементьев Г.П., Гладков Н.А., Птушенко Е.С., Спангенберг Е.П., Судиловская А.М. (1966). Птицы Советского Союза, т. 1 . Израильская программа научных переводов, Иерусалим.
- ^ Хагер, А. (1975). Исследования с дуплами летучей мыши Strix uralensis . Вар Фагельварльд, 34(4), 321.
- ^ Сибли, К.Г., и Алквист, Дж.Э. (1990). Филогения и классификация птиц: исследование молекулярной эволюции . Издательство Йельского университета.
- ^ Джилл, Фрэнк ; Донскер, Дэвид, ред. (2019). « Совы » . Всемирный список птиц, версия 9.1 . Международный союз орнитологов . Проверено 2 апреля 2019 г.
- ^ Jump up to: а б Уинк М., Эль-Сайед А.А., Зауэр-Гюрт Х. и Гонсалес Дж. (2009). Молекулярная филогения сов (Strigiformes) установлена на основе последовательностей ДНК митохондриального цитохрома b и ядерного гена RAG-1 . Ардеа, 97(4), 581-592.
- ^ Мликовский, Ири (2002): Кайнозойские птицы мира, Часть 1: Европа . Архивировано 20 мая 2011 г. в Wayback Machine . Нинокс Пресс, Прага.
- ^ Ли, М.Ю., Ли, СМ, Чон, Х.С., Ли, Ш., Пак, Дж.И. и Ан, Дж. (2018). Полный митохондриальный геном северной ушастой совы (Asio otus Linnaeus, 1758), определенный с помощью секвенирования нового поколения . Митохондриальная ДНК, часть B, 3(2), 494-495.
- ^ Саломонсен, Ф. (1931). Отчет о поездке на Фарерские острова . Датский Орн. Объединенный. Тисскр, 25, 3-37.
- ^ Янош Д. (1972). Die mittelpleistozäne Vogelfauna der Stránská skála . – В: Мусил Р. (ред.): Странска скала И. – Антропос (Брно) 20: 35-64.
- ^ Jump up to: а б Янош Д. (1976). Die Felsnische Tarkő und die Vertebratenfauna ihrer Ausfüllung . Исследование пещеры Карстеш 8: 3-106.
- ^ Jump up to: а б Янош, Д. 1978. Новый тонкий стратиграфический уровень в палеонтологической серии венгерского плейстоцена . Географические бюллетени 26 (1–3): 161–174.
- ^ Jump up to: а б Мурер-Шовир, К. (1975). Плейстоценовые фауны птиц Франции: систематика, эволюция и адаптация, палеоклиматическая интерпретация . Геобиос, 8(5), 333-ИН11.
- ^ Беккер, К., и Пипер, Х. (1982). По свидетельствам уральской совы Strix uralensis на неолитическом поселении на берегу озера в Швейцарии . Орнитологический обозреватель, 79, 159–162.
- ^ Гоффетт, К., Денис, М., Пёллат, Н., и Ван Нир, В. (2016). Изменение исторического ареала уральской совы в Европе . Бельгийский зоологический журнал, 146 (1), 33–43.
- ^ Шерзингер, В. (1983). Наблюдения за гибридами коричневой совы и ястребиной совы: (Strix aluco x Strix uralensis) .
- ^ Эк, С. (1968). Параллелизм рисунка вариации Strix . Зоол. Трактат Состояние Муз. животное в. Дрезден, 29: 283-288.
- ^ Вус, К.Х. (1990). Границы видов у нетропических сов Северного полушария . Вклад в зоологию, 60 (3/4), 163–170.
- ^ Кляйншмидт, О. (1934). Хищные птицы родины . Лейпциг.
- ^ Майр, Э., и Шорт, LL (1970). Видовые таксоны птиц Северной Америки: вклад в сравнительную систематику . Орнитологический клуб Наттолла.
- ^ Штреземанн, Э. (1923). Зоологические результаты экспедиций Вальтера Штёцнера в Сычуань, Восточный Тибет и Чили. 3 часть. Авес. Пассеры и Пикарии I. Деп. Муз. животное в. Фёлькерк. Дрезден, 16(2), 11-21.
- ^ Шерзингер, В. (2005). Замечания о сычуаньской лесной неясыти Strix uralensis davidi по наблюдениям на юго-западе Китая . БЮЛЛЕТЕНЬ-БРИТАНСКИЙ КЛУБ ОРНИТОЛОГОВ, 125(4), 275.
- ^ Клементс, Дж. Ф.; и др. (2015). « Контрольный список птиц мира eBird/Clements: v2015 » (таблица Excel) . Проверено 13 сентября 2015 г.
- ^ БердЛайф Интернэшнл . « Стрикс Давиди » . Архивировано из оригинала 4 марта 2016 г. Проверено 13 сентября 2015 г.
- ^ МСОП . «Красный список видов, находящихся под угрозой исчезновения МСОП» . Проверено 13 сентября 2015 г.
- ^ Хаускнехт, Р., Джейкобс, С., Мюллер, Дж., Цинк, Р., Фрей, Х., Сольхейм, Р., Врезец, А., Кристин, А., Михок, Дж., Кергалве, И. , Саурола П. и Кюн Р. (2014). Филогеографический анализ и распознавание генетических кластеров для сохранения уральских сов (Strix uralensis) в Европе . Журнал орнитологии, 155 (1), 121–134.
- ^ Пиетиайнен, Х., и Колунен, Х. (1993). Состояние тела самки и размножение уральской совы Strix uralensis . Функциональная экология, 726–735.
- ^ Врезец, А. (2009). Меланизм и изменчивость оперения макроуры уральской совы . Датч Бёрд, 31, 159–170.
- ^ Jump up to: а б с Уитц, М. (2011). Возможное распространение уральской совы Strix uralensis macroura в Центральной и Юго-Восточной Европе (Докторская диссертация, uniwien).
- ^ Мальчик, П. (2018). Третья встреча уральской совы Strix uralensis в Мазовии . Кулон, 23.
- ^ Пробст Р. и Малле Г. Уральская сова (Strix uralensis) в Каринтии . Сильва Фера, 2:26-30.
- ^ Цихоцкий, Влодзимеж и Оелизовский, Яцек и Боченский, Збигнев. (2004). Заметки о совах Польских Татр, юг Польши . Acta Zoologica Cracoviensia. 47. 9-16.
- ^ Стейнборн, Г. (2013). Свидетельства существования уральской совы Strix uralensis в природном парке Эггских гор/Южном Тевтобургском лесу . Харадриус, 49, 139–143.
- ^ Айе Р., Швейцер М. и Рот Т. (2012). Птицы Средней Азии . Издательство Блумсбери.
- ^ Ким, CH, Канг, Дж.-Х., Ким, Н.-Х. и Донг-Вон, К. (2011). Распространение длиннохвостой совы (Strix uralensis) в национальном парке Одэсан . Корейское общество науки о птицах, 13: 79-86.
- ^ Jump up to: а б Бразилия, М. (2019). Полевой справочник по птицам Восточной Азии . Издательство Блумсбери.
- ^ Глуц фон Блотцхайм, ООН, Бауэр, К.М., и Беззель, Э. (1980). Справочник птиц Центральной Европы . Аудитория, Висбаден.
- ^ Быличка М., Кайточ Л. и Фигарски Т. (2010). Особенности среды обитания и ландшафта, влияющие на встречаемость уральской совы Strix uralensis в агролесомелиоративной мозаике . Acta Ornithologica, 45(1), 33-42.
- ^ Болбоака, Л.Е., Балтаг, Э.С., Покора, В., и Ион, К. (2013). Избирательность среды обитания симпатрической неясыти (Strix aluco) и уральской совы (Strix uralensis) в горных лесах северо-восточной Румынии . Научные летописи Университета «Александру Иоан Куза» из Ясс, с. Биология животных, 59, 69–76.
- ^ Больбоака, Л.Е., Иордаш, И., и Ион, К. (2018). Факторы, связанные с распространением уральской совы Strix uralensis macroura в Восточной Румынии . Северо-Западный зоологический журнал, 14 (2).
- ^ Шерзингер, В. (1996). Динамика леса и требования к среде обитания уральской совы (Strix uralensis) . н/д.
- ^ Авотиньш, А. (2014). Анализ факторов, влияющих на выбор мест гнездования уральской совы Strix uralensis в Латвии .
- ^ Jump up to: а б с д Корпимяки, Э. (1987). Состав сообществ сов в четырех районах западной Финляндии: значение местообитаний и межвидовая конкуренция . Acta Regiae Soc Sci Litt Gothob Zool, 14, 118–123.
- ^ Бургас Д., Бихольм П. и Парккима Т. (2014). Хищники как суррогаты биоразнообразия в ландшафтном градиенте . Журнал прикладной экологии, 51(3), 786-794.
- ^ Резанов А.А. и Резанов А.Г. (2013). Гнездование длиннохвостой совы Strix uralensis вблизи человеческого жилья в Истринском районе Московской области: оценка степени синантропизации . Рус. орнитол. Журнал, 951.
- ^ Jump up to: а б Дравецкий М. и Обуч Дж. (2009). Вклад в знания о синантропизации и пищевой специализации уральской совы (Strix uralensis) в городской среде города Кошице (Восточная Словакия) . Словацкий журнал Raptor, 3, 51–60.
- ^ Врезец, А. (1996). Гнездится ли Strix uralensis в Люблянских болотах? Акроцефал, 78 (17), 160-162.
- ^ Jump up to: а б Корпимяки Э., Хаккарайнен Х., Лааксонен Т. и Васько В. (2009). Реакция сов и пустельг на пространственно-временные изменения их основной добычи . Ардеа, 97(4), 646-648.
- ^ Jump up to: а б Кайточ Л., Матысек М. и Фигарски Т. (2016). Пространственно-временные закономерности совиных территорий в фрагментированных лесах находятся под влиянием высшего хищника (уральской совы) . В Annales Zoologici Fennici (т. 53, № 3–4, стр. 165–175). Финское зоологическое и ботаническое издательство.
- ^ Jump up to: а б с д и ж Миккола, Х. (1970). О питании большой серой совы (Strix nebulosa), уральской совы (Strix uralensis) и филина (Bubo bubo) в Финляндии летом . Финская игра, 22, 97-104.
- ^ Лундберг, А. (1979). Оседлость, миграция и компромисс: адаптация к нехватке мест гнездования и пищевая специализация у трех видов фенноскандинавских сов . Экология, 41(3), 273-281.
- ^ Jump up to: а б Жлендер, Н. (2016). Территориальные и хищнические реакции европейской землеройки (Strix uralensis) на более мелких синтопических конкурентов: диссертация . университетское обучение (Докторская диссертация, Н. Жлендер).
- ^ Хотта, М., Маекава, М., Такизава, К., и Хосоно, Т. (2002). Высотное перемещение уральских сов Strix uralensis в зависимости от высоты снежного покрова . Вестник Научно-исследовательского института охраны природы Нагано (Япония).
- ^ Jump up to: а б с д и ж г час я дж Врезец А., Саурола П., Авотинс А., Косьянчич С. и Сулкава С. (2018). Сравнительное исследование диеты уральской совы Strix uralensis в период размножения в пределах ее европейского ареала, полученное на основе схем мониторинга гнездовых ящиков . Исследование птиц, 65(sup1), S85-S95.
- ^ Jump up to: а б с д и ж г час Казама, Т. (2005). Кольцевание и исследование рациона питания цыплят уральских сов Strix uralensis с использованием гнездовых ящиков . Журнал Японской ассоциации птиц, 2: 28-32.
- ^ Эскелинен О., Сулкава П. и Сулкава Р. (2004). Колебания численности лесного лемминга Myopus schisticolor в восточной и западной Финляндии . Acta theriologica, 49(2), 191-202.
- ^ Барта З. и Шеп Т. (1994). Поведение: Собирательство . Исследовательские заметки по биологии птиц 1994: Избранные материалы 21-го Международного орнитологического конгресса. Журнал für Ornithologie, 135 (1): 145–154.
- ^ Jump up to: а б с Нисимура, К. (1988). Кормовое поведение уральских сов (Strix uralensis) в неоднородной среде: важность полученной информации . Экологические исследования, 3(3), 319-332.
- ^ Jump up to: а б с д и ж Лундберг, А. (1980). Почему уральская неясыть Strix uralensis и неясыть S. aluco являются парапатрикальными в Скандинавии? Орнис Скандинавица, 116–120.
- ^ Jump up to: а б Нисимура, К. (1991). Использование участков различных типов добычи у уральской совы (Strix uralensis): хищника, сидящего и ожидающего . Поведенческая экология, 2(2), 99-105.
- ^ Нисимура, К., и Абэ, MT (1988). Восприимчивость добычи, использование добычи и различная эффективность атаки уральских сов . Экология, 77(3), 414-422.
- ^ Jump up to: а б с д Сузуки Т., Такацуку С., Хигучи А. и Сайто И. (2013). Пищевые привычки уральской совы (Strix uralensis) в период размножения в Центральной Японии . Журнал исследований хищников, 47(3), 304-311.
- ^ Аллен, М., Уорд, М., Юзник, Д. и Крофель, М. (2019). Совы-падальщики: глобальный обзор и новые наблюдения из Европы и Северной Америки . Журнал исследований хищников. 53.
- ^ Крофель, М. (2011). Мониторинг факультативных птиц-падальщиков туш крупных млекопитающих в динарском лесу Словении . Акроцефал, 32(148-149), 45-51.
- ^ Карлсен М., Лодал Дж., Лейрс Х. и Дженсен Т.С. (1999). Влияние риска нападения хищников на массу тела полевки Microtus agrestis . Ойкос, 277-285.
- ^ Коскела, Э., и Юленен, Х. (1995). Подавление размножения полевой полевки (Microtus agrestis): адаптация к циклически меняющемуся риску нападения хищников . Поведенческая экология, 6(3), 311-315.
- ^ Jump up to: а б Карелл П., Лехтосало Н., Пиетияйнен Х. и Броммер Дж. Э. (2010). Хищничество уральской совы на полевок и рыжих полевок по размеру, полу и репродуктивному состоянию . В Annales Zoologici Fennici (т. 47, № 2, стр. 90-99). Финское зоологическое и ботаническое издательство.
- ^ Юленен, Х., Витала, Дж., и Маппес, Т. (1991). Насколько сильно птичьи хищники влияют на циклические популяции рыжих полевок? Эксперимент в пиковый год . В Annales Zoologici Fennici (стр. 1–6). Совет финских зоологических издательств, созданный Академией наук Финляндии, Societas Biologica Fennica Vanamo, Societas pro Fauna et Flora Fennica и Societas Scientiarum Fennica.
- ^ Сайто, Т., Бьёрнстад, Онтарио, и Стенсет, Северная Каролина (1999). Зависимость плотности у полевок и мышей: сравнительное исследование . Экология, 80(2), 638-650.
- ^ Хаукисалми В., Хенттонен Х. и Пиетияйнен Х. (1994). Гельминтозный паразитизм не увеличивает уязвимость полевки Microtus agrestis к хищничеству уральской совы Strix uralensis . В Annales Zoologici Fennici (стр. 263–269). Совет финских зоологических издательств, созданный Академией наук Финляндии, Societas Biologica Fennica Vanamo, Societas pro Fauna et Flora Fennica и Societas Scientiarum Fennica.
- ^ Jump up to: а б с Тишечкин, А.К. (1997). Сравнительный анализ пищевой ниши сов Стрикс в Беларуси . В: Дункан, Джеймс Р.; Джонсон, Дэвид Х.; Николлс, Томас Х., ред. Биология и охрана сов Северного полушария: 2-й Международный симпозиум. Генерал Тех. Представитель NC-190. Сент-Пол, Миннесота: Министерство сельского хозяйства США, Лесная служба, Северо-Центральная лесная экспериментальная станция. 456-460. (Том 190).
- ^ Jump up to: а б с д и ж г час я Корпимяки Э. и Сулкава С. (1987). Рацион питания и репродуктивные качества уральских сов Strix uralensis . Орнис Фенника, 64, 57–66.
- ^ Jump up to: а б с Ядерхольм, К. (1987). Рацион Тенгмальмской совы Aegolius funereus и Уральской совы Strix uralensis в центральной Финляндии . Орнис Фенника, 64 (4), 149–153.
- ^ Jump up to: а б с д и ж г Корпимяки, Э. (1986). Нишевые отношения и тактика жизненного цикла трех симпатрических видов сов Strix в Финляндии . Орнис Скандинавица, 126–132.
- ^ Jump up to: а б с Лундберг, А. (1981). Популяционная экология уральской совы Strix uralensis в центральной Швеции . Орнис Скандинавица, 111–119.
- ^ Jump up to: а б с д и ж г час Моен, АГ (2015). Зависимость от еды: какую добычу приносят уральские совы (Strix uralensis) в гнездо в год с низкой численностью полевок и когда и что? (Магистерская диссертация, Норвежский университет естественных наук, Ос).
- ^ Jump up to: а б Лыхмус, А. (1999, январь). Регулярные колебания численности эстонской совы, вызванные полевками . В Annales Zoologici Fennici (стр. 167-178). Финское зоологическое и ботаническое издательство.
- ^ Jump up to: а б Сидорович В.Е., Шамович Д.И., Соловей И.А., Лаужель Г.О. (2003). Диетические вариации уральской совы Strix uralensis в переходных смешанных лесах северной Беларуси и их влияние на различия в распространении . Орнис Фенника, 80(4), 145–158.
- ^ Jump up to: а б Коцюба, М. (2012). Факторы, влияющие на состав рациона уральской совы Strix uralensis в предгорьях Центральных Бескид . Орнис Полоника, 53 (4), 283–292.
- ^ Jump up to: а б с д Штюрцер, С.Дж. (1998). Развитие популяции и экология питания уральской совы Strix uralensis и неясыти Strix aluco в национальном парке «Баварский лес» . Орн. Анз., 37: 109-119.
- ^ Врезец А. и Михелич Т. (2012). Уральская сова, Strix uralensis macroura, в Словении: обзор современных знаний об экологии видов . Итальянский журнал орнитологии.
- ^ Врезец, А. (2000). Вклад в знание диеты сорокопутов Strix uralensis macroura в Кочевско . Акроцефал, 21 (98-99): 77, 78.
- ^ Криштуфек, Б. (2010). Ухмыляющийся ухмылка (Rodentia: Gliridae) . Виды млекопитающих, 42(865), 195-206.
- ^ Jump up to: а б с Андрейчев А., Лапшин А. (2017). Количественный и качественный состав рациона уральской совы Strix Uralensi (Strigidae, Strigiformes) в центральной части Европейской России (на примере Республики Мордовия) . Вестник зоологии, 51(5), 421-428.
- ^ Jump up to: а б с д Карякин И. (1998). 2.1.3.2. Уральская неясыть — Strix uralensis Pall. Группа А, Категория 4 . Приложение II к Конвенции СИТЕС. Распространение птиц на Урале и сопредельных территориях в прошлом и настоящем по литературным источникам.
- ^ Шохрин, вице-президент (2016). Биология длиннохвостой совы Strix uralensis в зависимости от численности красно-серой полевки Clethrionomys rufocanus на юго-востоке Приморья . Рус. орнитол. Журнал, 1379.
- ^ Jump up to: а б с д и ж г час я дж Шибнев Ю.Б. (1989). К биологии длиннохвостой совы в Приморье . Бул. МОИП. Сентябрь биол. 94, 5:15-25.
- ^ Jump up to: а б с д и ж г Йонеда М., Абэ Х. и Накао Х. (1979). Зимние пищевые привычки йезской неясыти Strix uralensis japonica в ветрозащитной полосе . Журнал Института орнитологии имени Ямашина, 11(1), 49-53.
- ^ Врезец, А. (2001). Зимний рацион одной самки уральской совы (Strix uralensis) в Люблянском барье (центральная Словения) . Бутео, 12, 71–76.
- ^ Сайто, Х., Хашимото, Х., Хино, Т. и Мотокава, М. (2019). Как японская водяная землеройка Chimarrogale platycephalus пересекает бетонные стены заграждений? Исследование млекопитающих, 44 (1).
- ^ Jump up to: а б с Корпимяки Э., Хухтала К. и Сулкава С. (1990). Подтверждают ли межгодовые изменения в рационе филинов и уральских сов гипотезу об альтернативной добыче? Ойкос, 47-54.
- ^ Ёсихару, И. (1968). Анализ совиных погадок . Зоологический журнал, 77 (12), 402-404.
- ^ Линдстрём Э., Андрен Х., Ангелстам П. и Виден П. (1986). Влияние хищников на популяцию зайцев в Швеции: критический обзор . Обзор млекопитающих, 16(3-4), 151-156.
- ^ Ангербьёрн, А. (1995). Робкий кролик . Виды млекопитающих, (495), 1-11.
- ^ Хигучи А. и Абэ МТ (2001). Исследования энергетического баланса уральских сов, содержащихся в неволе, Strix uralensis . Японский журнал орнитологии, 50 (1), 25-30.
- ^ Рюзо, М. и Томоко, С. (1996). О содержании погадки уральской совы Strix uralensis hondoensis . Кагава Сейбуцу, 15:15-20.
- ^ Гриссер М. и Экман Дж. (2005). Непотистическое поведение сибирской сойки Perisoreus infaustus . Поведение животных, 69 (2), 345–352.
- ^ Бразилия, Массачусетс, и Ямамото, С. (1989). Статус и распространение сов в Японии . Хищники в современном мире. Берлин, Германия: Всемирная рабочая группа по хищным птицам, 389, 402.
- ^ Санига, М. (2002). Гибель гнезд и смертность птенцов глухаря (Tetrao urogallus) и рябчика (Bonasa bonasia) в Западных Карпатах . FOLIA ZOOLOGICA-PRAHA-, 51(3), 205-214.
- ^ Кук, МЫ (1987). Земноводные и рептилии: хищники и жертвы. Земноводные и птицы . Смитсоновская герпетологическая информационная служба.
- ^ Грибок, Ю. и Канеда, Х. (2009). Полевые наблюдения за хищничеством уральской совы Strix uralensis на японского рогатого жука Trypoxylus dichotomus septentrionalis . Ин-т Ж. Ямаскина Орнитология, 40(2), 90-95.
- ^ Сотеншек, Б. (2012). Диетические ниши симпатрических видов сипух (Strix uralensis) и лесных сов (Strix aluco) в период размножения . Дипломная работа, Любляна, Люблянский университет, 80.
- ^ Jump up to: а б с д Кайточ Л., Жмихорский М. и Вечорек П. (2015). Эффект смещения среды обитания между двумя конкурирующими видами сов во фрагментированных лесах . Популяционная экология, 57(3), 517-527.
- ^ Врх, П. и Врезец, А. (2006). Межвидовая территориальная вокальная активность уральской совы (Strix uralensis) по отношению к желтой неясыти (Strix aluco), конкуренту симпатрической совы: эксперимент по воспроизведению . Разправе, 47 (3): 99-105.
- ^ Солонен Т. и Кархунен Дж. (2002). Влияние переменных условий кормления на неясыть Strix aluco вблизи северной границы ее ареала. Орнис Фенника, 79(3), 121–131.
- ^ Солонен, Т. (2004). Влияют ли на сов-полёвок мягкие зимы на юге Финляндии? Орнис Фенника, 81(2), 65–74.
- ^ Jump up to: а б Врезец, Ал. (2003). Плотность гнездования и высотное распространение уральских, серых и бурых неясытей в Северо-Динарских Альпах (Центральная Словения) . Дж. Раптор Рес, 37 (1), 55-62.
- ^ Jump up to: а б с д Паченовский, С. (1995). К межвидовым отношениям Glaucidium passerinum, Strix uralensis и Strix aluco . Тиходрома, 8, 61-73.
- ^ Jump up to: а б Врезец А. и Томе Д. (2004). Выбор среды обитания и закономерности распределения в иерархической гильдии лесных сов . Орнис Фенника, 81 (3), 109–118.
- ^ Jump up to: а б с д и ж Кристин А., Михок Ю., Данко Ш., Караска Д., Паценовский С., Санига М., Бодова М., Балаж К., Шотнар К., Корнян Ю. и Олексак, М. (2007). Распространение, численность и сохранение уральской совы Strix uralensis в Словакии . Tagungsbericht des Nationalparks Bayerischer Wald, 8, 8-15.
- ^ Jump up to: а б Пентериани, В. и Дельгадо, MdM (2019). Филин . Монографии Пойзера.
- ^ Яношси Д. и Шмидт Э. (1970). Пища филина (Bubo bubo). Региональные и геологические изменения . Боннер зоол, ст. 21, 25-31.
- ^ Эндрюс, Питер (1990) Совы, пещеры и окаменелости: хищничество, сохранение и накопление мелких костей млекопитающих в пещерах, с анализом плейстоценовой пещерной фауны из Вестбери-суб-Мендип, Сомерсет, UK University of Chicago Press. 231 стр.
- ^ Донасар, Дж. А., Хиральдо, Ф., Делиб, М., и Эстрелла, Р. Р. (1989). Сравнительные пищевые привычки филина Bubo bubo и большой рогатой совы Bubo Virginianus в шести палеарктических и неарктических биомах . Орнис Скандинавица, 298–306.
- ^ Хаккарайнен Х., Корпимяки Э., Койвунен В. и Курки С. (1997). Реакция бореальной совы на управление лесами: обзор . Раптор Рес, 31 (2), 125–128.
- ^ Jump up to: а б с д и ж Миккола, Х. (1976). Совы убивают и убивают другие совы и хищники в Европе . Британские птицы, 69, 144–154.
- ^ Сулкава С., Хухтала К., Раджала П. и Торнберг Р. (1999). Изменения в питании беркута Aquila chrysaetos и популяций мелкой дичи в Финляндии в 1957-96 гг . Орнис Фенника, 76, 1–16.
- ^ Мошкин, А. (2009). Клептопаразитизм – один из способов охоты сапсана, получивший распространение в условиях увеличения его численности в горах Южного Урала, Россия . Сохранение хищников, (17).
- ^ Нечаев В.А. и Харченко В.А. (2012). Современное распространение и особенности биологии восточного орла (Spizaetus nipalensis orientalis Temmnick et Schelgel, 1844) в России . Русские Птицы, 571 (63): 238-244.
- ^ Крофель, М. (2012) Межвидовые взаимодействия евразийской рыси (Lynx lynx), связанные с хищничеством . Динариды. Доктор диссертация, Любляна, унив. в Любляне, факультет биотехнологии.
- ^ Дравецкий М., Данко Ш., Обух Ю., Кицко Дж., Мадерич Б., Караска Д., Врана Дж., Срейбр О., Шотнар К., Врлик П. и Богачик Л. (2008). Рацион малого подорлика (Aquila pomarina) в Словакии . Словацкий журнал Raptor, 2, 1-18.
- ^ Jump up to: а б Цинсмейстер, Д. (2012). Миграция молоди уральской совы (Strix uralensis) в биосферном парке Венский лес (Докторская диссертация, uniwien).
- ^ Jump up to: а б Ледицниг К. и Коль И. (2013). Реинтродукция уральской совы (Strix uralensis) в северные Известняковые Альпы . Сильва Фера - научные новости из дикой местности Дюрренштайн, 2, 78–93.
- ^ Книстаутас, AJV и Сибнев, JB (1987). Птичий мир Уссури . Гамбург-Берлин.
- ^ Jump up to: а б Солонен, Т. (1993). Расселение хищных птиц на юге Финляндии . Орнис Фенника, 70, 129–129.
- ^ Бышнев, И.И. (2002). Интересен случай агрессивного взаимодействия обыкновенного канюка (Buteobuteo) и уральской совы (Strix uralensis) . Субутео, 5: 46.
- ^ Важов С. и Бахтин Р. (2009). Некоторые сведения о биологии размножения уральской совы в окрестностях Бийска Алтайского края, Россия . Сохранение хищников, (17).
- ^ Моросинотто К., Томсон Р.Л., Ханнинен М. и Корпимяки Э. (2012). Более высокий риск нападения хищников на гнезда в сочетании с хищником высшего уровня: привлекательность мезохищников? Экология, 170(2), 507-515.
- ^ Jump up to: а б Хальме П., Хаккиля М. и Коскела Э. (2004). Защищают ли размножающиеся уральские совы Strix uralensis наземные гнезда птиц?: эксперимент с использованием фиктивных гнезд . Биология дикой природы, 10 (1), 145–149.
- ^ Саурола, П. (1987). Верность спариванию и месту гнездования у уральских неясытей и неясыти. Стр. 81–86 по биологии и охране северных лесных сов (Р.В. Неро, Р.Дж. Кларк, Р.Дж. Кнаптон и Р.Х. Хамре, ред.). Министерство сельского хозяйства США для. Серв. Генерал Тех. Реп.РМ-142.
- ^ Jump up to: а б Саурола, П. (1997). Мониторинг финских сов 1982-1996 гг.: методы и результаты . 2-й симпозиум по совам.
- ^ Корпимяки, Э. (1988). Факторы, способствующие полигинии у европейских хищных птиц — гипотеза . Экология, 77(2), 278-285.
- ^ Jump up to: а б Михелич Т., Врезец А., Перушек М. и Светличич Дж. (2000). Kozača strix uralensis в Словении . Акроцефал, 98(21), 9-22.
- ^ Jump up to: а б с д и ж г Врезец, А. (2016). Экология уральской совы на юго-западной границе ее распространения (Словения) . Рэпторс Консерв, 32, 8–20.
- ^ Ягелко Дж. и Вишневский М. (2012). Популяция уральской совы Strix uralensis в Силезских Бескидах . Давайте защитим Пржипр. Отец, 68(2), 83-90.
- ^ Сиах, М. (2005). Численность и закономерности распространения сов в Пенинском национальном парке, Южная Польша . Acta Zoologica Cracoviensia, 48(1-2), 21-33.
- ^ Jump up to: а б Матысек М., Фигарски Т., Вечорек П., Вика Дж. и Кайточ Л. (2015). Встречаемость евразийской совы Strix aluco и уральской совы S. uralensis в фрагментированных лесах центральной Малопольши . Орнис Полоника, 56(4).
- ^ Врезец, А. (2000). Инвентаризация коз Strix uralensis на Люблянском Врхе . Акроцефал, 21(98-99), 39-41.
- ^ Чапек, М. (1991). Дальнейшее гнездование уральской совы Strix uralensis в Моравско-Силезских Бескидах (северная Моравия, Чехословакия) . Время. Маллоу. Мужчина. Опава (А), 40:89, 90.
- ^ Башта А.-Т., Кучинская И. и Шидловский И. (2008). Уральская неясыть Strix uralensis Pall. экспансия в украинское Расточье . Институт экологии Карпат НАН Украины.
- ^ Jump up to: а б с Карякин И., Левашкин А. и Паженков А. (2009). Результаты привлечения уральской неясыти и неясыти в гнездовые ящики в Самарской области, Россия . Сохранение хищников, (16).
- ^ Балла, М. (2010). Уральская сова (Strix uralensis) гнездится в пойменном лесу на Восточно-Словацкой равнине . Словацкий журнал Raptor, 4, 105–108.
- ^ Андреевич Н.В., Константинович К.С. Редкий случай птенца длиннохвостой совы Strix uralensis в поселении на Южном Сахалине . Русский орнитологический журнал, 1351: 3920-3921.
- ^ Мистеруд И. и Хаген Ю. (1969). Питание уральской совы (Strix uralensis Pall.) в Норвегии . Новый. Маг. Зоол, 17, 165-167.
- ^ Андреенков О.В., Андреенкова Н.Г. и Жимулев И.Ф. (2010). Продолжение проекта по привлечению уральской совы в гнездовья в окрестностях Академгородка, Новосибирск, Россия . Сохранение хищников, (19).
- ^ Насу, Ю., Мурахама, С., Мацумуро, Х., Хасигути, Д., и Мурахама, К. (2007). Первая находка чешуекрылых из гнезд уральской совы в Японии . Прикладная энтомология и зоология, 42 (4), 607–612.
- ^ Ратайц У., Капла А. и Врезец А. Предварительная оценка жуков в гнездах сов, гнездящихся в норах: уральская неясыть (Strix uralensis) и неясыть серая (Strix aluco) .
- ^ Джалава, Дж. (1980). Tineidae (Lepidoptera) из гнезд уральской совы (Strix uralensis Pall.) . Энтомологические заметки, 60 (2), 96–100.
- ^ Jump up to: а б Лахти, Э. (1972). Места гнездования и особенности гнездования уральской совы Strix uralensis в Финляндии в период 1870-1969 гг . Орнис Фенника, 49, 91–97.
- ^ Мацуока С. и Шиина К. (2010). Пневые гнезда уральской совы Strix uralensis на Хоккайдо . Бюллетень Научно-исследовательского института лесного хозяйства и лесных товаров, Ибараки, (415), 69-74.
- ^ Сираиси, Х., и Китахара, М. (2007). Исследование экологии размножения уральской совы Strix uralensis, в которой использовалось искусственное гнездо, а также типов и видов ее добычи у северного подножия горы Фудзи в центральной Японии . Исследования горы Фудзи, 1, 17–23.
- ^ Jump up to: а б с Лундберг А. и Вестман Б. (1984). Репродуктивный успех, смертность и требования к месту гнездования уральской совы Strix uralensis в центральной Швеции . В Annales Zoologici Fennici (стр. 265–269). Академия наук Финляндии, Финское научное общество, Финское общество фауны и флоры и Финское биологическое общество Ванамо.
- ^ Сольхейм, Р., Беккен, Дж., Бьернстад, Р., Бай, Ф.Н., Хаген, Т.К., Исаксен, К., и Стрём, Х. (2009). Уральские совы Strix uralensis на границе: места гнездования не являются лимитирующим фактором . Ардеа, 97(4), 515-519.
- ^ Сольхейм, Р., и Бьорнстад, Р. (1987). Распространение и биология размножения маргинальной популяции уральской совы Strix uralensis на юго-востоке Норвегии . Акта Рег. Соц. наук. Литт. Готобургенсис, 14, 71–75.
- ^ Jump up to: а б с Пиетияйнен, Х. (1989). Сезонная и индивидуальная изменчивость приплода уральской совы Strix uralensis . Журнал экологии животных, 905–920.
- ^ Jump up to: а б с Пиетияйнен Х., Саурола П. и Вяйсянен Р.А. (1986). Родительский вклад в размер кладки и размера яиц уральской совы Strix uralensis . Орнис Скандинавица, 309–325.
- ^ Солонен, Т. (2010). Отражение крупномасштабных климатических явлений зимнего сезона и местных погодных условий в численности и частоте размножения хищных птиц-полёвок . Тенденции орнитологических исследований. Нова, Нью-Йорк, 95–119.
- ^ Врезец А. и Кохек К. (2002). Некоторые особенности размножения уральской совы Strix uralensis в Словении . Акроцефал, 23(115), 179-183.
- ^ Jump up to: а б Левашкин, АП (2009). Результаты привлечения длиннохвостой совы в искусственные гнезда в Нижегородской области, Россия . Русская консервация, 16.
- ^ Jump up to: а б Контиайнен П., Броммер Дж. Э., Карелл П. и Пиетийнен Х. (2008). Наследственность, пластичность и канализация размеров яиц уральской совы в циклической среде . Журнал эволюционной биологии, 21 (1), 88-96.
- ^ Jump up to: а б Контиайнен П., Пиетияйнен Х., Карелл П., Пихлая Т. и Броммер Дж. Э. (2010). Асинхронность высиживания — индивидуальное свойство самок уральских сов, повышающее выживаемость птенцов . Поведенческая экология, 21(4), 722-729.
- ^ Шерзингер, Ю.В. (1974). Успешное размножение уральских сов в национальном парке «Баварский лес» . Зоол. Сад, 44:59-61.
- ^ Пиетияйнен, Х. (1988). Качество сезона размножения, возраст и влияние опыта на репродуктивный успех уральской совы (Strix uralensis) . Аук, 105 (2), 316–324.
- ^ Пиетияйнен, Х., и Колюнен, Х. (1986). Определение возраста размножающихся уральских сов Strix uralensis . Орнис Фенн, 63, 26–27.
- ^ Jump up to: а б с д Контиайнен П., Пиетиянен Х., Хуттунен К., Карелл П., Колунен Х. и Броммер Дж. Э. (2009). Агрессивная уральская неясыть-мать набирает больше потомства . Поведенческая экология, 20(4), 789-796.
- ^ Jump up to: а б Броммер Дж. Э., Пиетийнен Х. и Колунен Х. (1998). Влияние возраста первого размножения на пожизненный репродуктивный успех и приспособленность уральской совы к циклическим условиям питания . Журнал экологии животных, 67 (3), 359–369.
- ^ Саурола, П. (1992). Популяционные исследования уральской совы Strix uralensis в Финляндии . Экология и охрана европейских сов, 28–31.
- ^ Броммер Дж. Э., Пихлаямяки О., Колунен Х. и Пиетияйнен Х. (2003). Последствия частичной асимметрии линьки для истории жизни . Журнал экологии животных, 72 (6), 1057–1063.
- ^ Jump up to: а б с Саурола П. и Фрэнсис К. (2018). К комплексному мониторингу популяций на основе полевых работ ринга-добровольцев: продуктивность, выживаемость и изменение популяции неясыти Strix aluco и уральской совы Strix uralensis в Финляндии . Исследование птиц, 65(sup1), S63-S76.
- ^ Броммер, Дж. Э., Пиетияйнен, Х., и Кокко, Х. (2002). Циклические изменения сезонного пополнения и эволюция сезонного снижения размера кладки уральской совы . Труды Лондонского королевского общества. Серия B: Биологические науки, 269(1491), 647-654.
- ^ Броммер, Дж. Э., Карелл, П., и Пиетияйнен, Х. (2004). Уральские совы, получающие дополнительное питание, увеличивают репродуктивную продуктивность с задержкой в один год . Экология, 139(3), 354-358.
- ^ Броммер, Дж. Э., Карелл, П., Пихлая, Т., Пейнтер, Дж. Н., Прайммер, Ч. Р. и Пиетиайнен, Х. (2003). Распределение пола уральской совы и родительские вложения в плохих условиях питания . Экология, 137(1), 140-147.
- ^ Коцюба, М. (2014). Места гнездования уральской совы Strix uralensis в западной части Предгорья Перемышля . Хроники Пшироде Ойчиста, 6(70).
- ^ Карелл, П. (2007). Краткосрочные и отдаленные последствия пищевых ресурсов на размножение уральской совы Strix uralensis . Отдел экологии птиц, факультет биологических и экологических наук. Факультет биологических наук Хельсинкского университета.
- ^ Ален, И. (1976). Лесное хозяйство и фауна позвоночных . Экологические бюллетени, 59-62.
- ^ Лыхмус, А. (2003). Страдают ли уральские совы (Strix uralensis) от отсутствия мест гнездования в управляемых лесах? Биологическая охрана, 110(1), 1-9.
- ^ Шандор, А.Д., и Домша, К. (2012). Специальные охраняемые территории для сохранения лесных птиц Румынии: оценка состояния и возможное расширение с использованием инструментов прогнозирования . Acta Zoologica Bulgarica, 64, 367–374.
- ^ Копий, Г. (2011). Расширение численности и ареала лесных бореальных сов (Glaucidium passerinum, Aegolius funereus, Strix uralensis, Strix nebulosa) в Восточно-Центральной Европе . Фогельвельт, 132, 207–214.
- ^ Вермузек З., Крженек Д. и Чернекова Б. (2004). Увеличение численности сорокопута (Strix uralensis) в Бескидах . Сильвия.–2004.–40.–С, 151–155.
- ^ Дворжак, Л. (2002). Доказано гнездование Strix uralensis на территории НОАК Бескиды в 2000 и 2001 годах .
- ^ Башта, АТ (2009). Динамика численности и расширение ареала уральской неясыти Strix uralensis на западе Украины . Ардеа, 97 (4), 483–488.
- ^ Саурола, П. (2009). Плохие и хорошие новости: изменение популяции финских сов в 1982–2007 гг . Ардеа, 97 (4), 469–483.
- ^ Саурола, П. (2012). Обзор мониторинга хищников в Финляндии . Акроцефал, 33 (154–155), 203–215.
- ^ Конторщиков В.В., Гринченко О.С., Иванов А.В., Петришева А.П., Севрюгин А.В., Челинцев Н.Г. (1996). Совы Московской области . Британские птицы, 89, 171–174.
- ^ Мацына А.И., Мацына Е.Л., Мацына А.А., Гришуткин Г.Ф., Спиридонов С.Н. (2010). Оценка эффективности мероприятий по охране птиц на линиях электропередачи 6--10 кВ в национальном парке «Смольный», Россия . Сохранение хищников, (20).
- ^ Мельников В.Н., Мельникова А.В. (2012). Первые результаты исследований случаев поражения птиц электрическим током в Ивановской области, Россия . Сохранение хищников, (24).
- ^ Франссон Т., Янссон Л., Колемайнен Т. и Веннингер Т. (2019). Столкновения с линиями электропередачи и поражение птиц электрическим током — анализ, основанный на данных о кольцевании в Швеции в 1990–2017 гг . Орнис Свечица, 29, 37–52.
- ^ Кухта А.Э. и Мацюра А.В. (2018). Анализ смертности птиц в результате транспортных происшествий В управлении безопасностью полетов . Украинский экологический журнал, 8(4), 351-356.
- ^ Юн, В., Пяо, З.Дж., Лэй, Г., Ван, XY, Конг, Ю.П. и Цзидинг, К. (2013). Смертность видов позвоночных на дорогах на горном шоссе Чанбайшань, провинция Цзилинь, Китай . Северо-Западный зоологический журнал, 9 (2).
- ^ Шумрада, Т. (2015). Столкновение птиц со стеклянным фасадом коммерческого здания в Любляне (центральная Словения) осенью 2012 года . Акроцефал, 36(164-165), 69-72.
- ^ Похья-Микра, М., Вуорисало, Т., и Микра, С. (2012). Организованное преследование хищных птиц в Финляндии: исторические и популяционно-биологические перспективы . Орнис Фенника, 89(1), 1.
- ^ Лыхмус, А. (2011). Трехлетняя периодичность исторических данных о преследовании хищников: показатель циклов полевок? Эстонский экологический журнал, 60(2), 155.
- ^ Воржишек П., Рейф Дж., Штястный К. и Бейчек В. (2008). Насколько эффективным может быть национальное законодательство в защите птиц? Пример из Чехии . Фолиа Зоол, 57(3), 221-230.
- ^ Мёрнер, Т., и Маттссон, Р. (1983). Туляремия у зимняка (Buteo lagopus) и уральской совы (Strix uralensis) . Журнал болезней дикой природы, 19 (4), 360–362.
- ^ Мейстер Т., Лусси Х., Баконьи Т., Шикутова С., Рудольф И., Фогль В., Винклер Х., Фрей Х., Хубалек З., Новотны Н. и Вайсенбёк, Х. (2008). Серологические доказательства сохраняющейся высокой активности вируса Усуту (Flaviviridae) и формирования коллективного иммунитета у диких птиц в Австрии . Ветеринарная микробиология, 127(3-4), 237-248.
- ^ Мурата, К. (2002). Распространенность кровяных паразитов у японских диких птиц . Журнал ветеринарной медицины, 64 (9), 785-790.
- ^ Коморова П., Шпакулова М., Гурникова З. и Ухрин М. (2015). Скребни рода Centrorhynchus (Palaeacanthocephala: Centrorhynchidae) хищных птиц (Falconiformes) и сов (Strigiformes) в Словакии . Паразитологические исследования, 114(6), 2273-2278.
- ^ Эллис, CJ (1971). Сравнительные измерения, хозяева и географическое распространение видов микротетрамеров (Nematoda: Tetrameridae) . Научный журнал штата Айова, 46 (1), 29–47.
- ^ Учида, С. (1948). Исследования мокрецов (Mallophaga) Японии и сопредельных территорий (подотряд Ischnocera Pt. I) . Японский медицинский журнал, 1 (4), 303–326.
- ^ Шерзингер, В. (1987). Реинтродукция уральской совы в Баварском национальном парке, Германия . В Биол. и Консерв. северных лесных сов. Симп. Учеб. Лесная служба Министерства сельского хозяйства США. Генерал Техн. Отчет. РМ-142 (стр. 75-80).
- ^ Энгедер, Т. (2003). Реинтродукция уральской совы (Strix uralensis) на австрийской стороне Богемского леса в 2001 году . Бутео, 13, 97–99.
- ^ Хорал Д., Хорт Л. и Клоубек Б. (1998). Доказано гнездование белого сорокопута (Strix uralensis) на Шумаве в 1998 году .
- ^ Шерзингер, В. (2014). Пересмотр разграничения подвидов среднеевропейских ястребов (Strix uralensis) . Трактаты Бременской ассоциации естественных наук 47 (2): 1-12.
- ^ Шеффер, Н. (1990). Наблюдения за выпущенными на волю уральскими совами (Strix uralensis) . Нет. Орн. Гес Бавария 29: 139, 154.
- ^ Цинк, Р. (2013). Реинтродукция уральских сов (Strix uralensis) на восточно-северной окраине Альп . Сильва Фера - научные новости из дикой местности Дюрренштайн, 2, 66-77.
Внешние ссылки
[ редактировать ]- Австрийский центр компетенции «Уральская сова» (на английском языке)
| Базы данных органов управления : Национальные |
|---|