Неконкурентное торможение
Неконкурентное ингибирование -это тип ингибирования фермента , когда ингибитор снижает активность фермента и в равной степени связывается с ферментом, независимо от того, уже связал ли он субстрат. [ 1 ] Это отличается от конкурентного ингибирования , где аффинность связывания к субстрату в ферменте снижается в присутствии ингибитора.
Ингибитор может связываться с ферментом, независимо от того, был ли субстрат уже связан, но если он имеет более высокое сродство к связыванию фермента в одном или другом состоянии, он называется смешанным ингибитором . [ 1 ]
История
[ редактировать ]За годы работы в качестве врача Леонора Михаэлиса и друга Питер Рона построили компактную лабораторию в больнице и в течение пяти лет - Михаэльс успешно стал опубликованным более чем 100 раз. Во время своего исследования в больнице он был первым, кто просматривал различные типы ингибирования; В частности, используя фруктозу и глюкозу в качестве ингибиторов малтазной активности. Малтаза разбивает мальтозу на две единицы глюкозы . Результаты этого эксперимента позволили осуществить расхождение неконкурентного и конкурентного ингибирования . Неконкурентное ингибирование влияет на значение k Cat (но не K M ) на любом данном графике; Этот ингибитор связывается с сайтом, который имеет специфичность для определенной молекулы. Михаэлис определил, что когда ингибитор связан, фермент станет инактивированным. [ 2 ]
Как и многие другие ученые своего времени, Леонор Михаэлис и Мод Менден работали над реакцией, которая использовалась для изменения состава сахарозы и превращения ее в два продукта - фруктоза и глюкоза. [ 2 ] Фермент, участвующий в этой реакции, называется инвертазой , и это фермент, кинетика которой была поддержана Михаэлисом и Менден, чтобы он революционер для кинетики других ферментов. При выражении скорости изучаемой реакции они вывели уравнение, которое описало скорость таким образом, что предполагало, что она в основном зависит от концентрации фермента, а также от присутствия субстрата, но только в определенной степени. [ 2 ] [ 3 ]
Адриан Джон Браун и Виктор Анри заложили основу для открытий в кинетике фермента, которыми известны Михаэлис и Менден. [ 4 ] Браун теоретически предполагал, что механизм, который теперь принимается для кинетики фермента, но не имел количественных данных для предъявления претензии. [ 4 ] Виктор Анри внес значительный вклад в кинетику фермента во время докторской диссертации, однако ему не хватало отмечения важности концентрации ионов водорода и мутаротации глюкозы. Целью тезиса Анри было сравнение его знаний о реакциях, катализируемых ферментами, на признанные законы физической химии. [ 2 ] Анри приписывается первым, кто написал уравнение, которое теперь известно как уравнение Михаэлис-Мюрмита. Используя глюкозу и фруктозу в каталитических реакциях, контролируемых Maltase и Invertase, Leonor Michaelis был первым ученым, который различает различные типы ингибирования, используя шкалу pH, которая не существовала во времена Анри. [ 2 ]
В частности, во время их работы по описанию скорости этой реакции они также тестировали и экстраполировали идею другого ученых, Виктора Анри , этого фермента, который они использовали, имел некоторое сродство к обоим продуктам этой реакции - фруктозы и глюкозы. [ 2 ] [ 3 ] Используя методы Анри, Михаэлис и Менден почти усовершенствовали эту концепцию метода начальной скорости для стационарных экспериментов. Они изучали ингибирование, когда обнаружили, что неконкурентное (смешанное) ингибирование характеризуется его влиянием на K CAT (скорость катализатора), в то время как конкуренция характеризуется его влиянием на скорость (V). [ 2 ] В экспериментах Michaelis and Menten они в значительной степени сосредоточились на эффектах инвертазы с использованием ионов водорода. [ 2 ] Инвертаза является ферментом, обнаруженным во внеклеточных дрожжах, и катализируемые реакции путем гидролиза или инвертирования сахарозы (смесь сахарозы и фруктозы) на « инверт сахар ». Основная причина использования инвертазы заключалась в том, что ее можно легко проанализировать, и эксперименты могут быть проведены более быстрым способом. Сахароза вращается в поляриметре как декстрорататор-D, тогда как инвертный сахар является леворотаторным-l . Это сделало отслеживание инверсии сахара относительно простой. Они также обнаружили, что α-D-глюкоза высвобождается в реакциях, катализируемой инвертазой, которая очень нестабильна и спонтанно изменяется в β-D-глюкозу . [ 4 ] Хотя они оба находятся в декстроротационной форме, именно здесь они отметили, что глюкоза может измениться спонтанно, также известная как мутаротация. Неспособность принять это во внимание было одной из главных причин, по которым эксперименты Анри не достигли. Используя инвертазу для катализации инверсии сахарозы, они могли видеть, насколько быстро фермент реагировал с помощью поляриметрии; Следовательно, было обнаружено, что неконкурентное ингибирование происходит в реакции, где сахарозу инвертировали инвертазой. [ 2 ]
Терминология
[ редактировать ]Важно отметить, что, хотя все неконкурентные ингибиторы связывают фермент в аллостерических сайтах (т.е. местоположения, кроме его активного сайта ),-не все ингибиторы, которые связываются в аллостерических сайтах, являются неконкурентными ингибиторами. [ 1 ] Фактически, аллостерические ингибиторы могут выступать в качестве конкурентных , неконкурентных или неконкуренторов . [ 1 ]
Многие источники продолжают объединять эти два термина, [ 5 ] или укажите определение аллостерического ингибирования как определение неконкурентного ингибирования.
Механизм
[ редактировать ]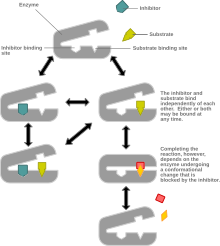
Неконкурентное ингибирование модели системы, в которой ингибитор и субстрат могут быть связаны с ферментом в любой момент времени. Когда как субстрат, так и ингибитор связаны, комплекс ингибитора фермента не может образовывать продукт и может быть преобразован только обратно в комплекс фермента-субстрата или комплекс ингибитора фермента. Неконкурентное ингибирование отличается от общего смешанного ингибирования в том смысле, что ингибитор обладает одинаковой аффинностью к ферменту и комплексу фермента-субстрата.
, катализируемых ферментами Например, в реакциях гликолиза , фосфоенол накопление катализируется пируваткиназой в пируват . Аланин представляет собой аминокислоту, которая синтезируется из пирувата, также ингибирует фермент пируваткиназу во время гликолиза. Аланин является неконкурентным ингибитором, поэтому он связывается с активным сайтом с субстратом, чтобы он все еще был конечным продуктом. [ 6 ]
Другой пример неконкурентного ингибирования дается глюкозо-6-фосфатом, ингибирующей гексокиназу в мозге. Углель 2 и 4 на глюкозе-6-фосфате содержат гидроксильные группы, которые прикрепляются вместе с фосфатом в углероде 6 к комплексу ингибитора фермента. Субстрат и фермент различаются в своих групповых комбинациях, к которым прикрепляется ингибитор. Способность глюкозо-6-фосфата связываться в разных местах одновременно делает его неконкурентным ингибитором. [ 7 ]
Наиболее распространенный механизм неконкурентного ингибирования включает обратимое связывание ингибитора с аллостерическим сайтом , но ингибитор может работать с другими способами, включая прямое связывание с активным сайтом. Он отличается от конкурентного ингибирования тем, что связывание ингибитора не предотвращает связывание субстрата и наоборот, но просто предотвращает образование продукта в течение ограниченного времени.

Этот тип ингибирования снижает максимальную скорость без химической реакции кажущейся аффинности связывания катализатора изменения для субстрата (K M приложение -См. Кинетику Михаэлиса-Мюмона ). Когда добавляется неконкурентный ингибитор, Vmax изменяется, а KM остается неизменным. Согласно графику Lineweaver-Burk, Vmax уменьшается во время добавления неконкурентного ингибитора, который показан на графике изменением как наклона, так и Y-перехвата, когда добавляется неконкурентный ингибитор. [ 8 ]
Основное различие между конкурентными и неконкурентными заключается в том, что конкурентное ингибирование влияет на способность субстрата связывать путем связывания ингибитора вместо субстрата, что снижает сродство фермента к субстрату. При неконкурентном ингибировании ингибитор связывается с аллостерическим сайтом и предотвращает выполнение химической реакции фермента-субстрата. Это не влияет на KM (аффинность) фермента (для субстрата). Неконкурентное ингибирование отличается от неконкурентоспособного ингибирования тем, что оно по-прежнему позволяет субстрату связываться с комплексом ингибитора фермента ингибитор через конформационные изменения при аллостерическом связывании.
Уравнение
[ редактировать ]В присутствии неконкурентного ингибитора кажущаяся аффинность фермента эквивалентна фактическому сродству. С точки зрения кинетики Михаэлиса-Мюртина , K M приложение = K m . Это можно рассматривать как следствие принципа Le Chatelier , потому что ингибитор связывается как с ферментом, так и с комплексом фермента-субстрата в равной степени, чтобы равновесие сохранялось. Однако, поскольку некоторый фермент всегда ингибируется от преобразования субстрата в продукт, эффективная концентрация фермента снижается.
Математически,
![{\displaystyle V_{max}^{app}={\frac {V_{max}}{1+{\frac {[I]}{K_{I}}}}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/1b945d1d3deff510520bdb20f9865068d4d12c4d)
![{\ displaystyle {ventent \ [e] _ {0}} = {\ frac {[e] _ {0}} {1+{\ frac {[i]} {k_ {i}}}}}}}}}}}}}}}}}}}}}}}}}}}}}}}}}}}}}}}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/b5dc3a4908e8409124b361d5e495f6d2f54f636e)
Пример: неконкурентные ингибиторы фермента CYP2C9
[ редактировать ]Неконкурентные ингибиторы CYP2C9 фермента включают нифедипин , транилсипромин , фенотилзотиоцианат и 6-гидроксифлавон. Заменимые мутанты компьютерной стыковки и построенные мутанты указывают, что неконкурентный сайт связывания 6-гидроксифлавона является зарегистрированным аллостерическим сайтом связывания CYP2C9 фермента . [ 9 ]
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в дюймовый Стрелоу Дж., Деве В., Айверсен П.В., Брукс Х.Б., Радинг Дж.А., МакГи Дж., Вейднер Дж. (2004). «Механизм действий анализа на ферменты». В Markossian S, Grossman A, Brimacombe K, Arkin M, Auld D, Austin CP, et al. (ред.). Руководство по анализу . Eli Lilly & Company и Национальный центр по продвижению трансляционных наук. PMID 22553872 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Корниш-Боуден А (2015-03-01). «Сто лет кинетики Михаэлис -Менден» . Перспективы в науке . Материалы симпозиума ESCEC Beilstein - празднование 100 -летия Михаэлиса Ментен -Конетика. 4 (Дополнение C): 3–9. doi : 10.1016/j.pisc.2014.12.002 .
- ^ Jump up to: а беременный Michaelis L, Menten MM (сентябрь 2013 г.). «Кинетика инвертина. 1913». Письма Febs . 587 (17): 2712–2720. doi : 10.1016/j.febslet.2013.07.015 . PMID 23867202 . S2CID 43226286 .
- ^ Jump up to: а беременный в Корниш-Боуден А (сентябрь 2013 г.). «Происхождение кинетики фермента» . Письма Febs . Столетие Михаэлиса - кинетика ментена. 587 (17): 2725–2730. doi : 10.1016/j.febslet.2013.06.009 . PMID 23791665 . S2CID 12573784 .
- ^ «Неконкурентное торможение и аллостерическое ингибирование» . Биология онлайн (форум). Архивировано с оригинала 25 апреля 2015 года . Получено 2 апреля 2012 года .
- ^ Berg JM, Tymoczko JL, Stryer L (2002). «Гликолитический путь плотно контролируется» . Биохимия (5 -е изд.).
- ^ Crane RK, Sols A (октябрь 1954 г.). «Неконкурентное ингибирование гексокиназы мозга глюкозо-6-фосфатом и родственными соединениями» (PDF) . Журнал биологической химии . 210 (2): 597–606. doi : 10.1016/s0021-9258 (18) 65385-2 . PMID 13211596 .
- ^ Waldrop GL (январь 2009 г.). «Качественный подход к ингибированию ферментов». Биохимия и молекулярное биологическое образование . 37 (1): 11–5. doi : 10.1002/bmb.20243 . PMID 21567682 . S2CID 205518237 .
- ^ Si D, Wang Y, Zhou YH, Guo Y, Wang J, Zhou H, et al. (Март 2009 г.). «Механизм ингибирования CYP2C9 флавонами и флавонолами» (PDF) . Метаболизм и расположение наркотиков . 37 (3). Американское общество фармакологии и экспериментальной терапии (ASPET): 629–634. doi : 10.1124/dmd.108.023416 . PMID 19074529 . S2CID 285706 .