Аккомодация (глаз позвоночного)


Аккомодация — это процесс, посредством которого позвоночного глаз изменяет оптическую силу , чтобы сохранять четкое изображение или фокусироваться на объекте при изменении расстояния до него. При этом расстояния у разных людей различаются от дальней точки — максимального расстояния от глаза, на котором можно увидеть четкое изображение объекта, до ближней точки — минимального расстояния для четкого изображения.Аккомодация обычно действует как рефлекс , включая часть рефлекса аккомодации-конвергенции , но ею также можно сознательно управлять.
Основные способы, с помощью которых животные могут менять фокус:
- Изменение формы линзы.
- Изменение положения хрусталика относительно сетчатки.
- Изменение осевой длины глазного яблока.
- Изменение формы роговицы.
Механизмы фокусировки
[ редактировать ]
Чтобы сфокусировать свет, рассеянный объектами в трехмерной среде, в двумерную совокупность отдельных ярких точек света, необходимо изогнуть свет. Чтобы получить хорошее изображение этих точек света на определенной области, требуется точное систематическое искривление света, называемое рефракцией . Настоящее изображение , сформированное из миллионов этих световых точек, — это то, что животные видят с помощью сетчатки . Очень равномерное систематическое искривление частей роговицы и хрусталика приводит к систематическому преломлению света на сетчатку.

Из-за особенностей оптики сфокусированное изображение на сетчатке всегда перевернуто относительно объекта.Разные животные живут в разных средах, имеющих разные показатели преломления, включая воду, воздух, а часто и то, и другое. Поэтому глазам необходимо преломлять свет в разной степени, что приводит к использованию разных механизмов фокусировки в разных средах. Граница раздела воздух/роговица имеет большую разницу в показателе преломления, чем гидратированные структуры внутри глаза. В результате у животных, живущих в воздухе, большая часть отклонения света достигается на границе раздела воздух/роговица, при этом хрусталик участвует в более точной фокусировке изображения.
Обычно млекопитающие, птицы и рептилии, живущие в воздухе, изменяют оптическую силу своих глаз, тонко и точно изменяя форму эластичного хрусталика с помощью цилиарного тела .
Небольшая разница в показателе преломления воды и гидратированной роговицы означает, что рыбам и амфибиям необходимо сильнее преломлять свет, используя внутренние структуры глаза. Следовательно, глаза, развившиеся в воде, имеют механизм, включающий изменение расстояния между жесткой, округлой и более преломляющей линзой и сетчаткой с использованием менее однородных мышц , а не тонкое изменение формы самой линзы с помощью циркулярно расположенных мышц. [1]
Наземные животные и линза, меняющая форму
[ редактировать ]Различные формы прямых экспериментальных доказательств, изложенные в этой статье, показывают, что большинство неводных позвоночных достигают фокусировки, по крайней мере частично, за счет изменения формы своих хрусталиков.
Менее понятно, как происходят тонкие, точные и очень быстрые изменения формы линзы. Прямое экспериментальное доказательство любой модели хрусталика обязательно затруднено, поскольку хрусталик позвоночных прозрачен и хорошо функционирует только у живых животных. При рассмотрении позвоночных аспекты всех моделей могут играть разную роль в фокусе линзы. Модели можно условно разделить на два лагеря. Те модели, которые подчеркивают важность внешних сил, действующих на более пассивно-эластичную линзу, и другие модели, которые включают силы, которые могут создаваться внутри линзы.
Внешние силы
[ редактировать ]Модель человеческой линзы, меняющей форму, была предложена Янгом в лекции 27 ноября 1800 года. [2] Другие, такие как Гельмгольц и Хаксли, усовершенствовали модель в середине 1800-х годов, объяснив, как сокращается цилиарная мышца, округляющая хрусталик, чтобы фокусироваться вблизи. [3] и эта модель была популяризирована Гельмгольцем в 1909 году. [4] [5] Модель можно резюмировать следующим образом. Обычно хрусталик удерживается под напряжением за счет подвешивающих связок , а капсула туго натягивается под давлением глазного яблока. На коротком фокусном расстоянии цилиарная мышца сокращается, растягивая цилиарное тело и снимая часть напряжения с поддерживающих связок, что позволяет хрусталику немного упруго скруглиться, увеличивая преломляющую силу. Для изменения фокуса на объекте, находящемся на большем расстоянии, требуется более тонкая и менее изогнутая линза. Это достигается за счет расслабления некоторых сфинктеров, таких как цилиарные мышцы, позволяя цилиарному телу пружинить назад, сильнее натягивая хрусталик, делая его менее изогнутым и тоньше, тем самым увеличивая фокусное расстояние . Проблема с моделью Гельмгольца заключается в том, что, несмотря на опробование математических моделей, ни одна из них не приблизилась к работе с использованием только механизмов Гельмгольца. [6]

Шахар предложил модель наземных позвоночных, которая не получила одобрения. [7] Теория позволяет математическому моделированию более точно отразить способ фокусировки хрусталика, принимая во внимание сложность поддерживающих связок и наличие радиальных, а также круговых мышц в цилиарном теле. [8] [9] В этой модели связки могут в разной степени натягивать хрусталик по экватору с помощью радиальных мышц, в то время как связки смещаются от экватора вперед и назад. [10] расслабляются в разной степени за счет сокращения круговых мышц. [11] Эти многочисленные действия [12] Работа с эластичной линзой позволяет более тонко менять форму линзы спереди. Не только изменение фокуса, но и коррекция аберраций объектива, которые в противном случае могли бы возникнуть из-за изменения формы, и при этом лучше подходящее для математического моделирования. [6]
« Цепная » модель фокуса линзы, предложенная Коулманом. [13] требует меньшего напряжения связок, удерживающих хрусталик. Вместо того, чтобы хрусталик в целом растягивался тоньше для зрения вдаль и позволял расслабиться для фокусировки вблизи, сокращение круговых цилиарных мышц приводит к тому, что хрусталик испытывает меньшее гидростатическое давление на свою переднюю часть. Передняя часть хрусталика может затем изменить свою форму между поддерживающими связками, подобно тому, как провисшая цепь, висящая между двумя полюсами, может изменить свою кривизну, когда полюса сближаются. Эта модель требует точного плавного движения только передней части линзы, а не пытается изменить форму линзы в целом. Хотя эта концепция может быть задействована в фокусировке, фотография Шаймпфлюга показала, что задняя часть хрусталика также меняет форму в живом глазу. [14]
Внутренние силы
[ редактировать ]

Когда в 1801 году Томас Янг предложил изменение формы хрусталика человека в качестве механизма фокальной аккомодации, он думал, что хрусталик может представлять собой мышцу, способную сокращаться. Этот тип модели называется внутрикапсулярной аккомодацией, поскольку он основан на активности внутри хрусталика. В Нобелевской лекции 1911 года Аллвар Гулстранд говорил на тему «Как я нашел внутрикапсулярный механизм аккомодации», и этот аспект фокусировки хрусталика продолжает исследоваться. [15] [16] [17] Янг потратил время на поиск нервов, которые могли бы стимулировать сокращение хрусталика, но безуспешно. С тех пор стало ясно, что хрусталик — это не простая мышца, стимулируемая нервом, поэтому модель Гельмгольца 1909 года взяла верх. Исследователи до двадцатого века не имели возможности воспользоваться многими более поздними открытиями и методами. Мембранные белки, такие как аквапорины , которые позволяют воде проникать в клетки и выходить из них, являются наиболее распространенным мембранным белком в хрусталике. [18] [19] Также широко распространены коннексины , которые обеспечивают электрическое соединение клеток. Электронная и иммунофлуоресцентная микроскопия показывают, что волокнистые клетки сильно различаются по структуре и составу. [20] [21] [22] Магнитно-резонансная томография подтверждает наличие слоев в хрусталике, которые могут обеспечивать различные планы рефракции внутри него. [23] Показатель преломления хрусталика человека варьируется примерно от 1,406 в центральных слоях до 1,386 в менее плотных слоях хрусталика. [24] Этот градиент индекса увеличивает оптическую силу линзы. становится известно о структуре хрусталика млекопитающих in situ По мере того, как благодаря фотографиям Шаймпфлюга , МРТ [14] [25] В ходе физиологических исследований становится очевидным, что хрусталик сам по себе не реагирует полностью пассивно на окружающую цилиарную мышцу, но может быть способен изменять свой общий показатель преломления посредством механизмов, включающих динамику воды в хрусталике, которые еще предстоит выяснить. [26] [27] [28] На прилагаемой микрофотографии показаны сморщенные волокна расслабленной овечьей линзы после ее удаления из животного, что указывает на укорочение волокон хрусталика во время аккомодации вблизи фокуса. Возрастные изменения хрусталика человека также могут быть связаны с изменениями динамики воды в хрусталике. [29] [30]
Человеческие глаза
[ редактировать ]Было предложено Амплиту аккомодации» объединить в этот раздел « . ( Обсудить ) Предлагается с июля 2024 г. |

Глаз молодого человека может менять фокус от расстояния (бесконечности) до расстояния до 6,5 см от глаза. [32] [33] Это резкое изменение фокусной силы глаза примерно на 15 диоптрий (обратная величина фокусному расстоянию в метрах) происходит в результате уменьшения напряжения связок , вызванного сокращением цилиарной мышцы . Этот процесс может произойти всего за 224 ± 30 миллисекунд при ярком свете. [34] Амплитуда аккомодации снижается с возрастом. К пятому десятилетию жизни амплитуда аккомодации может снизиться настолько, что ближняя точка глаза окажется более удаленной, чем расстояние чтения. Когда это происходит, пациент страдает пресбиопией . При возникновении пресбиопии людям с эмметропией (т. е. не нуждающимся в оптической коррекции зрения вдаль) потребуется оптическое приспособление для зрения вблизи ; те, кто близорук (близорук и нуждается в оптической коррекции для зрения вдаль или вдаль), обнаружат, что они лучше видят вблизи без коррекции вдаль; а те, кто страдает дальнозоркостью (дальнозоркостью), обнаружат, что им может потребоваться коррекция зрения как вдаль, так и вблизи. Обратите внимание, что эти эффекты наиболее заметны, когда зрачок большой; то есть при тусклом свете. Возрастное снижение аккомодации происходит почти повсеместно и составляет менее 2 диоптрий к тому времени, когда человек достигает 45–50 лет, и к этому времени большая часть населения заметит снижение своей способности фокусироваться на близких объектах и, следовательно, потребует очков. для чтения или бифокальные линзы. Аккомодация снижается примерно до 1 диоптрии в возрасте 70 лет. Зависимость амплитуды аккомодации от возраста графически суммируется выражением . Классические кривые Дуэйна [31]
Теории о том, как люди сосредотачиваются
[ редактировать ]- Гельмгольца — Наиболее распространенный [35] Теория аккомодации - это теория, предложенная Германом фон Гельмгольцем в 1855 году. При взгляде на далекий объект циркулярно расположенная цилиарная мышца расслабляется, позволяя хрусталика связкам и поддерживающим связкам натягивать хрусталик, уплощая его. Источником напряжения является давление, которое стекловидное тело и водянистая жидкость оказывают на склеру наружу . При взгляде на ближний объект цилиарные мышцы сокращаются (сопротивляясь внешнему давлению на склеру), вызывая ослабление связок хрусталика, что позволяет хрусталику снова принять более толстую и выпуклую форму.
- Шачар - Рональд А. Шачар в 1992 году предложил то, что было названо «довольно причудливой геометрической теорией». [36] который утверждает, что фокусировка хрусталика человека связана с повышенным напряжением хрусталика через экваториальные пояски; что при сокращении цилиарной мышцы увеличивается экваториальное зональное напряжение, в результате чего центральные поверхности хрусталика утолщаются, центральная толщина хрусталика увеличивается (передне-задний диаметр), а периферические поверхности хрусталика уплощаются. В то время как напряжение экваториальных поясков увеличивается во время аккомодации, передние и задние пояски одновременно расслабляются. Повышенное экваториальное зональное напряжение сохраняет стабильность хрусталика и выравнивает периферическую поверхность хрусталика во время аккомодации. Как следствие, сила тяжести не влияет на амплитуду аккомодации, и первичная сферическая аберрация смещается в отрицательную сторону во время аккомодации. [37] [38] Теория не нашла значительной независимой поддержки. [ нужна ссылка ]
- Цепная линия - Д. Джексон Коулман предполагает, что хрусталик, связка и переднее стекловидное тело составляют диафрагму между передней и стекловидной камерами глаза. [39] Сокращение цилиарной мышцы инициирует градиент давления между стекловидным телом и водным пространством, поддерживающим переднюю форму хрусталика. Именно в этой форме линзы возникает механически воспроизводимое состояние крутого радиуса кривизны в центре линзы с небольшим уплощением периферической передней линзы, т. е. форма в поперечном сечении цепной линии . Передняя капсула и связка образуют поверхность в форме батута или гамака, которая полностью воспроизводится в зависимости от круговых размеров, т. е. диаметра цилиарного тела (мышцы Мюллера). Таким образом, цилиарное тело направляет форму, как пилоны подвесного моста, но ему не требуется поддерживать экваториальную силу тяги, чтобы сплющить хрусталик. [40] [41]
Индуцированные эффекты аккомодации
[ редактировать ]Когда люди приспосабливаются к близкому объекту, они также сужают глаза и сужают зрачки . Сочетание этих трех движений (аккомодации, конвергенции и миоза ) находится под контролем ядра Эдингера-Вестфаля и называется ближней триадой , или рефлексом аккомодации . [42] необходима правильная конвергенция Хотя хорошо известно, что для предотвращения диплопии , функциональная роль сужения зрачков остается менее ясной. Возможно, это может увеличить глубину резкости за счет уменьшения апертуры глаза и, таким образом, уменьшить количество аккомодации, необходимой для фокусировки изображения на сетчатке. [43]
Существует измеримое соотношение ( коэффициент Маттиссена ) между тем, насколько конвергенция происходит из-за аккомодации (отношение AC/A, соотношение CA/C). Нарушения этого могут привести к проблемам с бинокулярным зрением . [44]
Описанные у человека аномалии аккомодации
[ редактировать ]Существует много типов аномалий аккомодации. Его можно разделить на две группы: пониженное размещение и повышенное размещение. [45] Снижение аккомодации может возникать вследствие физиологического (пресбиопия), фармакологического (циклоплегия) или патологического. [45] Чрезмерная аккомодация и спазм аккомодации являются типами повышенной аккомодации. [ нужна ссылка ]
Пресбиопия
[ редактировать ]Пресбиопия , физиологическая недостаточность аккомодации, обусловленная возрастными изменениями хрусталика (снижение эластичности и повышение твердости) и силы цилиарной мышцы, является наиболее распространенной формой дисфункции аккомодации. [45] Это приведет к постепенному снижению зрения вблизи.
Аккомодационная недостаточность
[ редактировать ]Аккомодационная недостаточность – состояние, при котором амплитуда аккомодации человека меньше физиологических пределов для его возраста. [45] Преждевременный склероз хрусталика или слабость цилиарной мышцы вследствие системных или местных случаев могут вызвать недостаточность аккомодации. [45] Недостаточность аккомодации подразделяется на различные категории. [ нужна ссылка ]
Плохое проживание
[ редактировать ]Плохая аккомодация – это состояние, похожее на аккомодационную недостаточность. При этом диапазон аккомодации будет нормальным, но после чрезмерной работы вблизи аккомодационная способность снизится. [46]
Паралич аккомодации
[ редактировать ]При параличе аккомодации амплитуда аккомодации либо заметно снижена, либо полностью отсутствует ( циклоплегия ). [47] Это может произойти из-за паралича цилиарной мышцы или паралича глазодвигательного нерва. [45] Парасимпатолитические препараты, такие как атропин, также вызывают паралич аккомодации. [46]
Неравное размещение
[ редактировать ]Если амплитуда аккомодации между глазами различается на 0,5 диоптрии и более, ее считают неравной. [47] органические заболевания, травма головы или функциональная амблиопия . Причиной неравной аккомодации могут быть [47]
Адаптационные условия
[ редактировать ]Неспособность к аккомодации также известна как инерция аккомодации. [47] В этом случае возникнут трудности с переездом из одной точки в другую. Могут возникнуть трудности с регулировкой фокуса на расстоянии и вблизи. [46] Это сравнительно редкое состояние.
Спазм аккомодации
[ редактировать ]Спазм аккомодации, также известный как цилиарный спазм, представляет собой состояние аномально чрезмерной аккомодации, которое находится вне произвольного контроля человека. [45] Зрение может быть нечетким из-за индуцированной псевдомиопии .
Адаптационный избыток
[ редактировать ]Избыток аккомодации возникает, когда человек использует больше приспособлений, чем обычно, для выполнения определенной работы вблизи. Современные определения просто рассматривают это как неспособность легко расслабиться. [47]
Водные животные
[ редактировать ]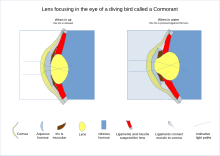

Среди водных животных есть те, которые прекрасно себя чувствуют и в воздухе, поэтому механизмы фокусировки различаются сильнее, чем у тех, которые обитают только на суше. Некоторые киты и тюлени способны фокусироваться над и под водой, имея две области сетчатки с большим количеством палочек и колбочек. [48] а не один, как у людей. Наличие двух областей сетчатки с высоким разрешением предположительно обеспечивает две оси зрения: одну над водой, другую под водой. У рептилий и птиц цилиарное тело, поддерживающее хрусталик посредством поддерживающих связок, также соприкасается с хрусталиком множеством подушечек на его внутренней поверхности. Эти подушечки сжимают и отпускают линзу, изменяя ее форму при фокусировке на объектах на разных расстояниях; эту функцию обычно выполняют поддерживающие связки у млекопитающих . При зрении у рыб и амфибий форма хрусталика фиксируется, и вместо этого фокусировка достигается за счет перемещения хрусталика вперед или назад внутри глаза с помощью мышцы, называемой ретрактором линзы. [49]
У хрящевых рыб поддерживающие связки заменены мембраной, включающей небольшую мышцу на нижней стороне хрусталика. Эта мышца вытягивает хрусталик вперед из расслабленного положения при фокусировке на близлежащих объектах. У костистых , напротив, из сосудистой структуры на дне глаза выступает мышца, называемая серповидным отростком , и служит для оттягивания хрусталика назад из расслабленного положения для фокусировки на удаленных объектах. Хотя амфибии, как и хрящевые рыбы, перемещают хрусталик вперед, задействованные мышцы не одинаковы ни у одного из типов животных. У лягушек имеются две мышцы: одна над хрусталиком и одна под хрусталиком, тогда как у других земноводных имеется только нижняя мышца. [49]
У простейших позвоночных — миног и миксин — хрусталик вообще не прикрепляется к внешней поверхности глазного яблока. Водянистой влаги у этих рыб нет, и стекловидное тело просто прижимает хрусталик к поверхности роговицы. Чтобы сфокусировать взгляд, минога сглаживает роговицу с помощью мышц снаружи глаза и отталкивает хрусталик назад. [49]
Хотя это и не позвоночные, здесь кратко упоминается конвергентная эволюция глаз позвоночных и моллюсков . Самый сложный глаз моллюсков - это глаз головоногих моллюсков , который внешне похож по структуре и функциям на глаз позвоночных, включая аккомодацию, но отличается в основных чертах, таких как наличие двухчастной линзы и отсутствие роговицы. [50] [51] Фундаментальные требования оптики должны быть выполнены всеми глазами с помощью линз с использованием имеющихся в их распоряжении тканей, поэтому на первый взгляд все глаза имеют тенденцию выглядеть одинаково. Это способ удовлетворения оптических потребностей с использованием различных типов клеток и структурных механизмов, который варьируется у разных животных.
См. также
[ редактировать ]Расстройства аккомодации и связанные с ней
[ редактировать ]Другой
[ редактировать ]- Размещение в рыбе
- Адаптация (глаз)
- Амплитуда размещения
- Циклоплегия
- Циклоспазм
- Ядро Эдингера-Вестфаля
- Эффект Мандельбаума
- Отрицательная относительная аккомодация
- Положительное относительное размещение
Ссылки
[ редактировать ]- ^ «Оген» . www.bio.vobs.at (на немецком языке) . Проверено 11 сентября 2020 г.
- ^ Лэнд, М (19 апреля 2015 г.). «Фокусировка путем изменения формы хрусталика глаза: комментарий к книге Янга (1801) «О механизме глаза» » . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 370 (1666). дои : 10.1098/rstb.2014.0308 . ПМК 4360117 . ПМИД 25750232 .
- ^ Хаксли, Томас Х (1871). Уроки элементарной физиологии (5-е изд.). Лондон и Нью-Йорк: МАКМИЛЛАН И КО., стр. 256–258.
- ^ Гельмгольц, Х. фон (1962). Трактат по физиологической оптике (английский перевод под редакцией JPC Southall. Оптическое общество Америки. Из третьего немецкого издания Handbuch der Physiologischen Optik (1909), Леопольд Восс, Лейпциг. Переиздание Дувра). Нью-Йорк, штат Нью-Йорк: Dover Publications Inc.
- ^ Фишер, РФ (1 августа 1977 г.). «Сила сокращения цилиарной мышцы человека при аккомодации» . Журнал физиологии . 270 (1): 51–74. дои : 10.1113/jphysicalol.1977.sp011938 . ПМЦ 1353417 . ПМИД 915798 .
- ^ Перейти обратно: а б Шачар, Рональд А.; Бакс, Эндрю Дж. (июнь 2001 г.). «Механизм аккомодации человека, анализируемый с помощью нелинейного анализа методом конечных элементов». Комплексная терапия . 27 (2): 122–132. дои : 10.1007/s12019-996-0006-5 . ПМИД 11430259 . S2CID 71369369 .
- ^ Атчисон, Д.А. (июль 1995 г.). «Аккомодация и пресбиопия». Офтальмологическая и физиологическая оптика . 15 (4): 255–72. doi : 10.1046/j.1475-1313.1995.9500020e.x . ПМИД 7667018 . S2CID 24282106 .
- ^ Мэйсяо; Сюй, Чжэ; Карп, Ван, Цзяньхуа (1 июня 2015 г.). Шао, Илей; Тао, Цзян, Хун; Чжун, Цзянгуан , Сопутствующие изменения в биометрии переднего сегмента во время аккомодации» . & Visual Science . 56 (6): 3522–3530. doi : 10.1167/iovs.15-16825 . PMC 4464043. . PMID 26030106 Investigative Ophthalmology
- ^ Шачар, Рональд А. (22 сентября 2015 г.). «Изменения конфигурации аккомодационных цилиарных мышц человека соответствуют механизму аккомодации Шахара». Исследовательская офтальмология и визуальные науки . 56 (10): 6075. doi : 10.1167/iovs.15-17452 . ПМИД 26393665 .
- ^ Стритен, BW (апрель 1977 г.). «Поясная вставка: исследование сканирующей электронной микроскопии» . Исследовательская офтальмология и визуальные науки . 16 (4): 364–375. ISSN 0146-0404 . ПМИД 844993 .
- ^ Шачар, РА (март 1994 г.). «Зонулярная функция: новая гипотеза с клиническим значением». Анналы офтальмологии . 26 (2): 36–8. ПМИД 8010701 .
- ^ Кнаус, Кэтрин Р.; Хипсли, ЭннМари; Блемкер, Сильвия С. (июнь 2021 г.). «Влияние сокращения цилиарной мышцы на аккомодацию хрусталика, исследованное с помощью 3D-модели». Биомеханика и моделирование в механобиологии . 20 (3): 879–894. дои : 10.1007/s10237-021-01417-9 . ПМИД 33491156 . S2CID 231704221 .
- ^ Коулман, Д. Джексон (июнь 1970 г.). «Единая модель аккомодационного механизма». Американский журнал офтальмологии . 69 (6): 1063–1079. дои : 10.1016/0002-9394(70)91057-3 . ПМИД 5423772 .
- ^ Перейти обратно: а б Херманс, Эрик А.; Пауэлс, Петра Дж.В.; Дуббельман, Мишель; Куйер, Йост, Пенсильвания; ван дер Хейде, Роб Г.Л.; Хитаар, Роб М. (1 января 2009 г.). «Постоянный объем хрусталика человека и уменьшение площади поверхности капсульного мешка во время аккомодации: исследование МРТ и Шаймпфлюга» . Исследовательская офтальмология и визуальные науки . 50 (1): 281–289. дои : 10.1167/iovs.08-2124 . ПМИД 18676625 .
- ^ ПАУ, Ч (1952). «[Аккомодационное смещение ядра хрусталика при внутрикапсулярной аккомодации]». Клинические ежемесячные информационные бюллетени по офтальмологии и офтальмологическому обучению . 121 (2): 224–6. ПМИД 14955961 .
- ^ Хуггерт, Арне (27 мая 2009 г.). «Внутрикапсулярный механизм аккомодации». Акта офтальмологическая . 42 (2): 389–397. дои : 10.1111/j.1755-3768.1964.tb03627.x . ПМИД 14213923 . S2CID 37325357 .
- ^ Лопес-Хиль, Норберто (3 марта 2022 г.). «Пересмотр механизма внутрикапсульной аккомодации Галлстранда» . Фотоника . 9 (3): 152. Бибкод : 2022Фото...9..152Л . doi : 10.3390/photonics9030152 .
- ^ Броехейсе, РМ; Кульманн, Эд; Столс, А.Л. (сентябрь 1976 г.). «Мембраны хрусталика II. Выделение и характеристика основного внутреннего полипептида (MIP) мембран бычьих волокон хрусталика». Экспериментальное исследование глаз . 23 (3): 365–371. дои : 10.1016/0014-4835(76)90135-4 . ПМИД 976377 .
- ^ Малдерс, С.М.; Престон, генеральный менеджер; Дин, премьер-министр; Гуджино, ВБ; ван Ос, Швейцария; Агре, П. (14 апреля 1995 г.). «Свойства водного канала основного внутреннего белка хрусталика» . Журнал биологической химии . 270 (15): 9010–16. дои : 10.1074/jbc.270.15.9010 . hdl : 2066/216433 . ПМИД 7536742 .
- ^ Кушак, Дж; Алькала, Дж; Мейзел, Х. (декабрь 1980 г.). «Поверхностная морфология клеток хрусталика эмбрионов и взрослых кур». Американский журнал анатомии . 159 (4): 395–410. дои : 10.1002/aja.1001590406 . ПМИД 7223675 .
- ^ Грюйтерс, WT; Кистлер, Дж; Булливант, С. (октябрь 1987 г.). «Формирование, распределение и диссоциация межклеточных соединений в хрусталике». Журнал клеточной науки . 88 (Часть 3) (3): 351–9. дои : 10.1242/jcs.88.3.351 . ПМИД 3448099 .
- ^ Грюйтерс, WT (июль 1989 г.). «Белок, не являющийся коннексоном (MIP), участвует в формировании щелевых соединений хрусталика». Журнал клеточной науки . 93 (Часть 3) (3): 509–13. дои : 10.1242/jcs.93.3.509 . ПМИД 2691517 .
- ^ Германс, Э.А.; Дуббельман, М; Ван дер Хейде, Р.; Хитаар, РМ (декабрь 2008 г.). «Эквивалентный показатель преломления хрусталика человека при аккомодационной реакции». Оптометрия и наука о зрении . 85 (12): 1179–84. дои : 10.1097/OPX.0b013e31818e8d57 . ПМИД 19050472 . S2CID 205907383 .
- ^ Хехт, Юджин. Оптика , 2-е изд. (1987), Эддисон Уэсли, ISBN 0-201-11609-X . п. 178.
- ^ Станке, Т.; Хадлич, С.; Ври, А.; Гутхофф, Р.; Стахс, О.; Лангнер, С. (16 декабря 2016 г.). «Магнитно-резонансная микроскопия аппарата аккомодации». Клинические ежемесячные бюллетени по офтальмологии . 233 (12): 1320–1323. дои : 10.1055/s-0042-118599 . ПМИД 27984837 . S2CID 78808282 .
- ^ Вагефи, Э; Понтре, BP; Джейкобс, доктор медицины; Дональдсон, П.Дж. (август 2011 г.). «Визуализация динамики жидкости в хрусталике глаза с помощью МРТ: манипулирование стабильным содержанием воды и потоками воды». Американский журнал физиологии. Регуляторная, интегративная и сравнительная физиология . 301 (2): Р335-42. дои : 10.1152/ajpregu.00173.2011 . ПМИД 21593426 . S2CID 9525037 .
- ^ Дональдсон, Пол Дж.; Чен, Яди; Петрова, Росица С.; Грей, Ангус К.; Лим, Джули К. (декабрь 2022 г.). «Регулирование содержания воды в хрусталике: влияние на физиологическую оптику хрусталика». Прогресс в исследованиях сетчатки и глаз . 95 : 101152. doi : 10.1016/j.preteyeres.2022.101152 . ПМИД 36470825 . S2CID 254243790 .
- ^ Сузуки-Керр, Х; Уокер, КЛ; Хан, Миннесота; Лим, Джей Си; Дональдсон, Пи Джей (2022). «Гипосмотический стресс вызывает высвобождение АТФ в отдельной зоне внешней коры хрусталика крысы» . Молекулярное видение . 28 : 245–256. ПМЦ 9514545 . ПМИД 36284672 .
- ^ Моффат, бакалавр; Ландман, штат Калифорния; Траскотт, Р.Дж.; Суини, Миннесота; Поуп, Дж. М. (декабрь 1999 г.). «Возрастные изменения кинетики транспорта воды в хрусталике нормального человека». Экспериментальное исследование глаз . 69 (6): 663–9. дои : 10.1006/exer.1999.0747 . ПМИД 10620395 .
- ^ Джонс, CE; Атчисон, округ Колумбия; Медер, Р.; Поуп, Дж. М. (август 2005 г.). «Распределение показателя преломления и оптические свойства изолированного хрусталика человека, измеренные с помощью магнитно-резонансной томографии (МРТ)». Исследование зрения . 45 (18): 2352–2366. дои : 10.1016/j.visres.2005.03.008 . ПМИД 15979462 . S2CID 8894700 .
- ^ Перейти обратно: а б Дуэйн, Александр (1922). «Исследования монокулярной и бинокулярной аккомодации и их клиническое применение» . Американский журнал офтальмологии . 5 (11): 865–877. дои : 10.1016/s0002-9394(22)90793-7 . S2CID 43172462 .
- ^ Чен, Ай Хун; О'Лири, Дэниел Дж.; Хауэлл, Эдвин Р. (2000). «Функция зрения вблизи у детей раннего возраста. Часть I: точка конвергенции вблизи. Часть II: амплитуда аккомодации. Часть III: гетерофория вблизи». Офтальмологическая и физиологическая оптика . 20 (3): 185–198. дои : 10.1046/j.1475-1313.2000.00498.x . ISSN 0275-5408 .
- ^ Это значение следует из максимальной аккомодационной способности и может быть рассчитано как 100 см/15 дпт.
- ^ Локхарт, TE; Ши, В. (2010). «Влияние возраста на динамическую аккомодацию» . Эргономика . 53 (7): 892–903. дои : 10.1080/00140139.2010.489968 . ПМК 2908311 . ПМИД 20582770 .
- ^ Баумайстер, М.; Конен, Т. (июнь 2008 г.). «Аккомодация и пресбиопия: Часть 1: Физиология аккомодации и развития пресбиопии». Офтальмолог (на немецком языке). 105 (6): 597–610. дои : 10.1007/s00347-008-1761-8 . ПМИД 18594896 .
- ^ Атчисон, Дэвид А. (1995). «Аккомодация и пресбиопия». Офтальмологическая и физиологическая оптика . 15 (4): 255–212. doi : 10.1046/j.1475-1313.1995.9500020e.x . ПМИД 7667018 . S2CID 24282106 .
- ^ Шачар, РА (2012). Механизм аккомодации и пресбиопия . Гаага: Публикации Куглера. ISBN 978-90-6299-233-1 .
- ^ Чжоу, XY; Ван, Л; Чжоу, ХТ; Ю, ZQ (24 октября 2014 г.). «Изменения аберрации волнового фронта, вызванные градиентом увеличения стимулов аккомодации» . Глаз . 29 (1): 115–121. дои : 10.1038/eye.2014.244 . ISSN 0950-222X . ПМЦ 4289835 . ПМИД 25341432 .
- ^ Коулман, Д. Джексон (июнь 1970 г.). «Единая модель аккомодационного механизма». Американский журнал офтальмологии . 69 (6): 1063–1079. дои : 10.1016/0002-9394(70)91057-3 . ISSN 0002-9394 . ПМИД 5423772 .
- ^ Коулман, диджей (1986). «К теории аккомодации гидравлической подвески» . Trans Am Ophthalmol Soc . 84 : 846–68. ПМЦ 1298753 . ПМИД 3590482 .
- ^ Коулман, диджей; Фиш, СК (сентябрь 2001 г.). «Пресбиопия, аккомодация и зрелая цепная связь». Офтальмология . 108 (9): 1544–51. дои : 10.1016/s0161-6420(01)00691-1 . ПМИД 11535447 .
- ^ Бхола, Рахул (23 января 2006 г.). «EyeRounds.org: Учебное пособие: бинокулярное зрение» . webeye.ophth.uiowa.edu . Университет Айовы . Проверено 11 сентября 2020 г.
- ^ Ван, Б.; Чуффреда, К.Дж. (2006). «Глубина резкости человеческого глаза: теория и клинические последствия». Обзор офтальмологии . 51 (1): 75–85. doi : 10.1016/j.survophthal.2005.11.003 . ПМИД 16414364 .
- ^ Уильям Дж., Бенджамин (2006). «Слияние и бинокулярность». Клиническая рефракция Бориша (2-е изд.). Сент-Луис, Миссури: Баттерворт Хайнеманн/Эльзевир. ISBN 978-0-7506-7524-6 .
- ^ Перейти обратно: а б с д и ж г Хурана, АК (сентябрь 2008 г.). «Астенопия, аномалии аккомодации и конвергенции». Теория и практика оптики и рефракции (2-е изд.). Эльзевир. стр. 100–107. ISBN 978-81-312-1132-8 .
- ^ Перейти обратно: а б с Герцог, Старейшина (1969). «Аномалии аккомодации». Практика рефракции (8-е изд.). Лондон: Черчилль. ISBN 0-7000-1410-1 .
- ^ Перейти обратно: а б с д и Уильям Дж., Бенджамин (2006). «Аккомодация, ученик и пресбиопия». Клиническая рефракция Бориша (2-е изд.). Сент-Луис, Миссури: Баттерворт Хайнеманн/Эльзевир. п. 112. ИСБН 978-0-7506-7524-6 .
- ^ Масс, Алла М.; Супин, Александр Я. (июнь 2007 г.). «Адаптивные особенности глаза водных млекопитающих» . Анатомические записи: достижения в интегративной анатомии и эволюционной биологии . 290 (6): 701–715. дои : 10.1002/ar.20529 . ПМИД 17516421 . S2CID 39925190 .
- ^ Перейти обратно: а б с Ромер, Альфред Шервуд; Парсонс, Томас С. (1977). Тело позвоночного . Филадельфия, Пенсильвания: Холт-Сондерс Интернэшнл. стр. 463–464. ISBN 978-0-03-910284-5 .
- ^ Джаггер, WS; Сэндс, П.Дж. (1 августа 1999 г.). «Широкоугольная градиентно-индексная оптическая модель хрусталика и глаза осьминога». Исследование зрения . 39 (17): 2841–2852. дои : 10.1016/S0042-6989(99)00012-7 . ПМИД 10492814 . S2CID 17808919 .
- ^ Шеффель, Ф.; Мерфи, CJ; Хауленд, ХК (15 ноября 1999 г.). «Размещение у каракатицы (Sepia officinalis)» . Журнал экспериментальной биологии . 202 (22): 3127–3134. дои : 10.1242/jeb.202.22.3127 . ПМИД 10539961 .
Внешние ссылки
[ редактировать ]- oph/723 в eMedicine — «Пресбиопия: причина и лечение»
- Окуляр + проживание в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
| Иллюзии |
| |
|---|---|---|
| Популярная культура |
| |
| Связанный | ||
| Базы данных органов управления : Национальные |
|---|