Зрение у рыб

Зрение — важная сенсорная система для большинства видов рыб . рыб Глаза похожи на глаза наземных позвоночных, таких как птицы и млекопитающие, но имеют более сферическую линзу . Птицы и млекопитающие (включая человека) обычно регулируют фокус , изменяя форму хрусталика, а рыбы обычно регулируют фокус, перемещая хрусталик ближе к сетчатке или дальше от нее . Сетчатка рыб обычно имеет как палочки, так и колбочки (для скотопического и фотопического зрения ), и большинство видов обладают цветовым зрением . Некоторые рыбы видят ультрафиолет , а некоторые чувствительны к поляризованному свету .
Среди рыб минога бесчелюстных [ 1 ] имеет хорошо развитые глаза, а у миксины лишь примитивные глазные пятна . [ 2 ] Предки современных миксин, считающихся протопозвоночными, [ 3 ] очевидно, были вытеснены в очень глубокие темные воды, где они были менее уязвимы для зрячих хищников и где выгодно иметь выпуклое глазное пятно, собирающее больше света, чем плоское или вогнутое. Зрение рыб демонстрирует эволюционную адаптацию к зрительной среде: например, глаза глубоководных рыб приспособлены к темноте.
Вода как визуальная среда
[ редактировать ]

Рыбы и другие водные животные живут в другой световой среде, чем наземные виды. Вода поглощает свет , поэтому с увеличением глубины количество доступного света быстро уменьшается. Оптические свойства воды также приводят к тому, что свет разной длины волны поглощается в разной степени. Например, видимый свет с длинными волнами (например, красный, оранжевый) поглощается водой больше, чем свет с более короткими длинами волн (зеленый, синий). Ультрафиолетовый свет (длина волны которого даже короче, чем у фиолетового) может проникать глубже, чем зрительные спектры. [ 5 ] Помимо этих универсальных качеств воды, разные водоемы могут поглощать свет разной длины волны из-за различного присутствия солей и/или химических веществ в воде.
Вода очень эффективно поглощает падающий свет, поэтому количество света, проникающего в океан, быстро уменьшается (ослабляется) с глубиной. В прозрачной океанской воде на глубине одного метра сохраняется лишь 45% солнечной энергии, попадающей на поверхность океана. На глубине 10 метров все еще присутствует только 16% света, и только 1% исходного света остается на глубине 100 метров. Ни один свет не проникает дальше 1000 метров. [ 6 ]
Помимо общего ослабления, океаны поглощают свет разных длин волн с разной скоростью. Длины волн на крайних концах видимого спектра ослабляются быстрее, чем длины волн в середине. В первую очередь поглощаются более длинные волны. В чистых океанских водах красный цвет поглощается на глубине 10 метров, оранжевый — примерно на 40 метрах, а желтый исчезает на глубине до 100 метров. Более короткие волны проникают дальше, а синий и зеленый свет достигают самых глубоких глубин. [ 6 ] Вот почему под водой предметы кажутся синими: то, как цвета воспринимаются глазом, зависит от длины волны света, воспринимаемого глазом. Объект кажется глазу красным, потому что он отражает красный свет и поглощает другие цвета. Таким образом, единственный цвет, который попадает в глаза, — красный. Синий — единственный цвет света, доступный на глубине под водой, поэтому это единственный цвет, который может отражаться обратно в глаз, а под водой все имеет синий оттенок. Красный объект на глубине не будет казаться красным, потому что красный свет не может отразиться от объекта. Объекты в воде будут выглядеть в своих реальных цветах только вблизи поверхности, где все длины волн света все еще доступны, или если другие длины волн света обеспечиваются искусственно, например, путем освещения объекта подводным светом. [ 6 ]
Структура и функции
[ редактировать ]

Глаза рыб во многом похожи на глаза других позвоночных, особенно четвероногих (амфибий, рептилий, птиц и млекопитающих, которые произошли от предков-рыб). Свет попадает в глаз через роговицу , проходит через зрачок и достигает хрусталика . Большинство видов рыб, по-видимому, имеют фиксированный размер зрачка, но пластиножаберные (например, акулы и скаты) имеют мускулистую радужку , которая позволяет регулировать диаметр зрачка. Форма зрачков варьируется и может быть, например, круглой или щелевидной. [ 5 ]
Линзы обычно имеют сферическую форму, но у некоторых видов могут быть слегка эллиптическими. По сравнению с наземными позвоночными линзы рыб обычно более плотные и сферические. В водной среде нет большой разницы в показателе преломления роговицы и окружающей воды (по сравнению с воздухом на суше), поэтому большую часть преломления приходится выполнять линзе. [ 7 ] Из-за « градиента показателя преломления внутри линзы - точно так, как и следовало ожидать из оптической теории», [ 8 ] Сферические линзы рыб способны формировать четкие изображения без сферических аберраций . [ 7 ]
Когда свет проходит через хрусталик, он передается через прозрачную жидкую среду, пока не достигнет сетчатки, содержащей фоторецепторы . Как и у других позвоночных, фоторецепторы находятся на внутреннем слое, поэтому свет должен пройти через слои других нейронов, прежде чем достичь их. Сетчатка содержит клетки-палочки и колбочки. [ 5 ] Глаза рыб имеют сходство с глазами других позвоночных. Обычно свет проникает через «рыбий глаз» в роговицу и проходит через зрачок, чтобы достичь хрусталика. У большинства видов рыб зрачок фиксированного размера, а у некоторых видов имеется мускулистая радужная оболочка, позволяющая регулировать диаметр зрачка.
Глаза рыб имеют более сферическую линзу, чем у других наземных позвоночных. Регулировка фокуса у млекопитающих и птиц обычно осуществляется путем изменения формы глазного хрусталика, а у рыб — путем перемещения хрусталика дальше или ближе к сетчатке. Сетчатка рыбы обычно содержит как палочки, так и колбочки, которые отвечают за скотопическое и фотопическое зрение. Большинство видов рыб обладают цветовым зрением. Некоторые виды способны видеть ультрафиолет, а некоторые чувствительны к поляризованному свету. [ 9 ]
Сетчатка рыб имеет палочки, которые обеспечивают высокую зрительную чувствительность в условиях низкой освещенности, и колбочки, которые обеспечивают более высокое временное и пространственное разрешение, чем способны палочки. Они обеспечивают возможность цветового зрения за счет сравнения оптической плотности колбочек разных типов. [ 10 ] По данным Маршалла и др. Большинство животных морской среды обитания не обладают цветовым зрением или обладают относительно простым цветовым зрением. Однако в океане цветовое зрение больше, чем на суше. В основном это связано с крайностями в световой среде обитания и цветовом поведении. [ 11 ]
Сетчатка
[ редактировать ]В сетчатке палочки обеспечивают высокую зрительную чувствительность (за счет остроты зрения ) и используются в условиях низкой освещенности. Колбочки обеспечивают более высокое пространственное и временное разрешение, чем палочки, и обеспечивают возможность цветового зрения путем сравнения оптической плотности колбочек разных типов, которые более чувствительны к разным длинам волн. Соотношение палочек и колбочек зависит от экологии рассматриваемых видов рыб: например , те рыбы, которые в основном активны в течение дня в прозрачной воде, будут иметь больше колбочек, чем те, которые живут в условиях низкой освещенности. Цветовое зрение более полезно в средах с более широким диапазоном доступных длин волн, например , у поверхности в чистой воде, а не в более глубокой воде, где сохраняется только узкий диапазон длин волн. [ 5 ]
Распределение фоторецепторов по сетчатке неравномерно. Например, в некоторых областях плотность колбочек выше (см. ямку ) . У рыбы могут быть две или три области, отвечающие за высокую остроту зрения (например, при поимке добычи) или чувствительность (например, от тусклого света, исходящего снизу). Распределение фоторецепторов также может меняться со временем в ходе развития человека. Это особенно актуально, когда вид в течение своего жизненного цикла обычно перемещается между различными световыми средами (например, от мелководья к глубоководью или от пресной воды к океану). [ 5 ] или когда изменения пищевого спектра сопровождают рост рыбы, как это видно на примере антарктической ледяной рыбы Champsocephalus Gunnari . [ 12 ]
У некоторых видов есть тапетум — отражающий слой, который отражает свет, проходящий через сетчатку, обратно через нее. Это повышает чувствительность в условиях низкой освещенности, например, у ночных и глубоководных видов, давая фотонам второй шанс захватить фоторецепторы. [ 7 ] Однако за это приходится платить снижением разрешения. Некоторые виды способны эффективно отключать свой тапетум в ярких условиях, при необходимости покрывая его слоем темного пигмента. [ 5 ]
Сетчатка потребляет много кислорода по сравнению с большинством других тканей и снабжается обильным количеством крови, насыщенной кислородом, для обеспечения оптимальной работы. [ 5 ]
Размещение
[ редактировать ]Аккомодация — это процесс, посредством которого глаз позвоночного настраивает фокус на объекте по мере его приближения или удаления. В то время как птицы и млекопитающие достигают аккомодации за счет деформации хрусталика глаза, рыбы и амфибии обычно регулируют фокус , перемещая хрусталик ближе или дальше от сетчатки. [ 5 ] Они используют специальную мышцу, которая изменяет расстояние хрусталика от сетчатки. У костных рыб эта мышца называется ретрактором чечевицы и расслабляется при зрении вблизи, тогда как у хрящевых рыб мышца называется ретрактором чечевицы и расслабляется при зрении вдаль. Таким образом, костные рыбы приспосабливаются к зрению вдаль, перемещая хрусталик ближе к сетчатке, а хрящевые рыбы приспосабливаются к зрению вблизи, перемещая хрусталик дальше от сетчатки. [ 13 ] [ 14 ] [ 15 ]
Стабилизация изображений
[ редактировать ]
Существует необходимость в каком-то механизме, который стабилизирует изображение при быстрых движениях головы. Это достигается за счет вестибулоокулярного рефлекса , который представляет собой рефлекторное движение глаз , стабилизирующее изображение на сетчатке , производя движения глаз в направлении, противоположном движениям головы, сохраняя таким образом изображение в центре поля зрения. Например, когда голова движется вправо, глаза движутся влево, и наоборот. Вестибулоокулярный рефлекс человека — это рефлекторное движение глаз , которое стабилизирует изображение на сетчатке во время движения головы, вызывая движение глаз в направлении, противоположном движению головы, сохраняя таким образом изображение в центре поля зрения. Подобным образом у рыб есть вестибулоокулярный рефлекс, который стабилизирует зрительные изображения на сетчатке, когда она двигает хвостом. [ 16 ] У многих животных, включая человека, внутреннее ухо функционирует как биологический аналог акселерометра в системах стабилизации изображения камеры, стабилизируя изображение путем перемещения глаз . При обнаружении вращения головы тормозной сигнал посылается экстраокулярным мышцам с одной стороны и возбуждающий сигнал мышцам с другой стороны. Результатом является компенсаторное движение глаз. Типичные движения глаз человека отстают от движений головы менее чем на 10 мс. [ 17 ]
На схеме справа показана схема горизонтального вестибулоокулярного рефлекса у костных и хрящевых рыб .
- «Золотая рыбка» демонстрирует основной трехнейрональный вестибулоокулярный рефлекс, связывающий горизонтальный полукружный канал с контралатеральными отводящими (ABD) и ипсилатеральными MR- мотонейронами . [ 18 ]
- «Камбалка» показывает, что после смещения вестибулярного аппарата на 90° относительно зрительной оси (метаморфоза) компенсаторные движения глаз производятся за счет перенаправления сигналов горизонтальных каналов на вертикальные и косые мотонейроны. [ 19 ] [ 20 ]
- В «Акуле» горизонтальный канал/ нейроны второго порядка проецируются на контралатеральные мотонейроны ABD и MR, включая ипсилатеральные нейроны AI . 1° — вестибулярный нейрон первого порядка; АТД — восходящий путь Дейтера . [ 20 ]
Ультрафиолетовый
[ редактировать ]Зрение рыб осуществляется четырьмя зрительными пигментами, которые поглощают свет различной длины. Каждый пигмент состоит из хромофора и трансмембранного белка, известного как опсин . Мутации опсина позволили добиться визуального разнообразия, включая изменение поглощения длины волны. [ 21 ] Мутация опсина пигмента SWS-1 позволяет некоторым позвоночным поглощать ультрафиолетовый свет (≈360 нм), поэтому они могут видеть объекты, отражающие ультрафиолетовый свет. [ 22 ] Многие виды рыб развили и сохранили эту визуальную особенность на протяжении всей эволюции, что позволяет предположить ее преимущество. УФ-зрение может быть связано с поиском пищи, общением и выбором партнера.
Ведущая теория эволюционного отбора ультрафиолетового зрения у некоторых видов рыб обусловлена его важной ролью в выборе партнера. Поведенческие эксперименты показывают, что африканские цихлиды используют визуальные сигналы при выборе партнера. Их места размножения обычно находятся на мелководье с высокой прозрачностью и проникновением ультрафиолетового света. Самцы африканских цихлид в основном имеют синий цвет, отражающий ультрафиолетовый свет. Самки могут правильно выбрать партнера своего вида, когда присутствуют эти отражающие визуальные сигналы. Это говорит о том, что обнаружение ультрафиолетового излучения имеет решающее значение для правильного выбора партнера. [ 23 ] Цветовые узоры, отражающие УФ-излучение, также повышают привлекательность самцов гуппи и трехиглой колюшки. В экспериментальных условиях самки гуппи тратили значительно больше времени на осмотр самцов с УФ-отражающей окраской, чем те, у которых УФ-отражение было заблокировано. [ 24 ] Аналогичным образом, самки трехиглой колюшки предпочитали смотреть на самцов в полном спектре, а не на тех, которые просматривались через фильтры, блокирующие УФ-излучение. [ 25 ] Эти результаты убедительно свидетельствуют о роли обнаружения УФ-излучения в половом отборе и, следовательно, в репродуктивной приспособленности. Выдающаяся роль обнаружения ультрафиолетового света в выборе партнера по рыбе позволила сохранить этот признак с течением времени. УФ-зрение также может быть связано с поиском пищи и другим коммуникативным поведением.
Многие виды рыб способны видеть ультрафиолетовый конец спектра, выходящий за пределы фиолетового. [ 26 ]
Ультрафиолетовое зрение иногда используется только в течение части жизненного цикла рыбы. Например, молодь кумжи обитает на мелководье, где они используют ультрафиолетовое зрение, чтобы улучшить свою способность обнаруживать зоопланктон . По мере взросления они перемещаются в более глубокие воды, где мало ультрафиолета. [ 22 ]
Двухполосая стрекоза , Dascyllus reticulatus отражающую ультрафиолет , имеет окраску, , которую они, по-видимому, используют в качестве сигнала тревоги для других рыб своего вида. [ 27 ] Хищные виды не смогут этого увидеть, если их зрение не чувствительно к ультрафиолету. Есть еще одно подтверждение этой точки зрения: некоторые рыбы используют ультрафиолет как «высокоточный секретный канал связи, скрытый от хищников», в то время как другие виды используют ультрафиолет для передачи социальных или сексуальных сигналов. [ 5 ] [ 28 ]
Поляризованный свет
[ редактировать ]Нелегко установить, чувствительна ли рыба к поляризованному свету , хотя это вполне вероятно у ряда таксонов. Это было однозначно продемонстрировано на анчоусах . [ 29 ] Способность обнаруживать поляризованный свет может обеспечить лучшую контрастность и/или информацию о направлении мигрирующих видов. Поляризованный свет наиболее распространен на рассвете и в сумерках. [ 5 ] Поляризованный свет, отраженный от чешуи рыбы, может помочь другим рыбам лучше обнаружить его на рассеянном фоне. [ 30 ] и могут предоставить стайным рыбам полезную информацию об их близости и ориентации относительно соседних рыб. [ 31 ] Некоторые эксперименты показывают, что, используя поляризацию, некоторые рыбы могут настроить свое зрение, чтобы увеличить расстояние наблюдения добычи вдвое по сравнению с обычным. [ 9 ]
Двойные конусы
[ редактировать ]У большинства рыб есть двойные конусы — пара конусных клеток, соединенных друг с другом. Каждый член двойного конуса может иметь разный пик поглощения, и поведенческие данные подтверждают идею о том, что каждый тип отдельных колбочек в двойном конусе может предоставлять отдельную информацию (т. е. сигналы от отдельных членов двойного конуса не обязательно суммируются). . [ 32 ]
Адаптация к среде обитания
[ редактировать ]
Четырехглазая рыба питается у поверхности воды, глаза позволяют ей видеть одновременно и над, и под поверхностью.

1 Подводная сетчатка 2) Хрусталик 3) Воздушный зрачок
4) Тканевая полоска 5) Радужная оболочка 6) Подводный зрачок
7) Воздушная сетчатка 8) Зрительный нерв



Рыбы, обитающие в поверхностных водах на глубине примерно 200 метров, эпипелагические рыбы , обитают в освещенной солнцем зоне, где зрительные хищники используют зрительные системы, устроенные примерно так, как и можно было ожидать. Но даже в этом случае могут быть необычные адаптации. У четырехглазых рыб глаза подняты над макушкой и разделены на две разные части, так что они могут одновременно видеть под и над поверхностью воды. У четырехглазых рыб на самом деле всего два глаза, но их глаза специально приспособлены к образу жизни, обитающему на поверхности. Глаза расположены на макушке головы, и рыба плавает на поверхности воды, оставляя под водой только нижнюю половину каждого глаза. Две половины разделены полосой ткани, и глаз имеет два зрачка , соединенных частью радужной оболочки . Верхняя половина глаза приспособлена для зрения на воздухе, нижняя — для зрения в воде. [ 35 ] Толщина хрусталика глаза меняется сверху вниз, чтобы учесть разницу в показателях преломления воздуха и воды. Большую часть времени эти рыбы проводят у поверхности воды. Их рацион в основном состоит из наземных насекомых, обитающих на поверхности. [ 36 ]
Мезопелагические рыбы обитают в более глубоких водах, в сумеречной зоне до глубины 1000 метров, где количества доступного солнечного света недостаточно для поддержания фотосинтеза . Эти рыбы приспособлены к активной жизни в условиях недостаточной освещенности. Большинство из них — зрительные хищники с большими глазами. У некоторых глубоководных рыб глаза трубчатые с большими линзами и только палочковые клетки смотрят вверх. Они обеспечивают бинокулярное зрение и высокую чувствительность к слабым световым сигналам. [ 37 ] Эта адаптация дает улучшенное конечное зрение за счет бокового зрения и позволяет хищнику различать кальмаров , каракатиц и более мелкую рыбу, силуэты которых вырисовываются на фоне мрака над ними. Для более чувствительного зрения при слабом освещении имеется ретрорефлектор у некоторых рыб за сетчаткой . Рыбы-фонарики имеют этот плюс фотофоры , которые они используют в сочетании, чтобы обнаружить блеск глаз у других рыб. [ 38 ] [ 39 ] [ 40 ]
Еще глубже в толще воды , ниже 1000 метров, водятся батипелагические рыбы . На этой глубине океан угольно-черный, а рыбы ведут оседлый образ жизни, приспособленные к выработке минимума энергии в среде обитания с очень небольшим количеством пищи и отсутствием солнечного света. Биолюминесценция — единственный свет, доступный на этих глубинах. Отсутствие света означает, что организмам приходится полагаться не только на зрение, но и на другие чувства. Их глаза маленькие и могут вообще не функционировать. [ 41 ] [ 42 ]
На самом дне океана камбалу можно встретить . Камбала — это донная рыба с отрицательной плавучестью, поэтому она может отдыхать на морском дне. Хотя камбалы и являются донными обитателями, они обычно не являются глубоководными рыбами, а встречаются главным образом в эстуариях и на континентальном шельфе. Когда личинки камбалы вылупляются, они имеют удлиненную и симметричную форму типичной костной рыбы . Личинки не обитают на дне, а плавают в море в виде планктона . Со временем они начинают превращаться во взрослую форму. Один из глаз мигрирует через макушку головы на другую сторону тела, оставляя рыбу слепой с одной стороны. Личинка теряет плавательный пузырь и шипы и опускается на дно, кладя слепую сторону на подстилающую поверхность. [ 43 ] Ричард Докинз объясняет это как пример эволюционной адаптации.
...костные рыбы, как правило, имеют выраженную тенденцию сплющиваться в вертикальном направлении.... Поэтому было естественно, что, когда предки [камбалы] опустились на морское дно, они должны были лежать на одном боку .... Но при этом возникла проблема: один глаз всегда смотрел в песок и был фактически бесполезен. В ходе эволюции эта проблема была решена за счет «перемещения» нижнего глаза на верхнюю сторону. [ 44 ]
-
![Большинство глубоководных рыб не видят красного света. Глубоководный стоп-сигнал «Свободная челюсть» излучает красную биолюминесценцию, поэтому он может охотиться с помощью практически невидимого луча света.[45]](//upload.wikimedia.org/wikipedia/commons/thumb/b/b2/Malacosteus.JPG/179px-Malacosteus.JPG) Большинство глубоководных рыб не видят красного света. Глубоководный стоп-сигнал «Свободная челюсть» излучает красную биолюминесценцию, поэтому он может охотиться с помощью практически невидимого луча света. [ 45 ]
Большинство глубоководных рыб не видят красного света. Глубоководный стоп-сигнал «Свободная челюсть» излучает красную биолюминесценцию, поэтому он может охотиться с помощью практически невидимого луча света. [ 45 ] -
 Когда личинки камбалы растут , глаз с одной стороны поворачивается в другую сторону, чтобы рыба могла отдыхать на морском дне.
Когда личинки камбалы растут , глаз с одной стороны поворачивается в другую сторону, чтобы рыба могла отдыхать на морском дне. -
 Европейская камбала — это камбала с приподнятыми глазами, поэтому, когда она в целях маскировки зарывается в песок, она все еще может видеть.
Европейская камбала — это камбала с приподнятыми глазами, поэтому, когда она в целях маскировки зарывается в песок, она все еще может видеть.
![Большинство глубоководных рыб не видят красного света. Глубоководный стоп-сигнал «Свободная челюсть» излучает красную биолюминесценцию, поэтому он может охотиться с помощью практически невидимого луча света.[45]](http://upload.wikimedia.org/wikipedia/commons/thumb/b/b2/Malacosteus.JPG/179px-Malacosteus.JPG)


Глаза у добычи обычно расположены по бокам головы, поэтому у нее есть большое поле зрения, позволяющее избегать хищников. У хищников глаза обычно расположены впереди головы, поэтому они лучше воспринимают глубину . [ 46 ] [ 47 ] У донных хищников, таких как камбалы , глаза устроены таким образом, что они могут бинокулярно видеть то, что находится над ними, когда они лежат на дне.
Цвет
[ редактировать ]Рыбы развили сложные способы использования окраски . Например, хищные рыбы используют окраску, чтобы хищникам было труднее их увидеть. У пелагических рыб эти адаптации в основном связаны с уменьшением силуэта, формой камуфляжа . Один из способов добиться этого — уменьшить площадь их тени путем бокового сжатия тела. Другой метод, также являющийся формой камуфляжа, заключается в затенении в случае эпипелагических рыб и встречном освещении в случае мезопелагических рыб . Противозатенение достигается путем окрашивания рыбы более темными пигментами вверху и более светлыми пигментами внизу таким образом, чтобы окраска соответствовала фону. Если смотреть сверху, более темная спинная область животного сливается с темнотой воды внизу, а если смотреть снизу, более светлая брюшная область сливается с солнечным светом с поверхности. Противоосвещение достигается за счет биолюминесценции за счет образования света от вентральной части тела. фотофоры , направленные на согласование интенсивности света, исходящего от нижней части рыбы, с интенсивностью света, исходящего от фона. [ 48 ]
Бентические рыбы , покоящиеся на морском дне, физически прячутся, зарываясь в песок или прячась в укромные уголки и щели, или маскируют себя, сливаясь с фоном или выглядя как камень или кусок морской водоросли. [ 49 ]
Хотя эти инструменты могут быть эффективными в качестве механизмов избегания хищников, они также служат не менее эффективными инструментами для самих хищников. Например, глубоководная бархатная акула-фонарь использует встречное освещение, чтобы спрятаться от добычи. [ 50 ]
-
 Эпипелагические рыбы, такие как атлантический голубой тунец , обычно окрашены в серебристый цвет.
Эпипелагические рыбы, такие как атлантический голубой тунец , обычно окрашены в серебристый цвет. -
 У четырехглазой рыбы-бабочки на заднем конце есть ложные глаза, что сбивает с толку хищников, относительно которых находится передняя часть рыбы.
У четырехглазой рыбы-бабочки на заднем конце есть ложные глаза, что сбивает с толку хищников, относительно которых находится передняя часть рыбы. -
 У Джона Дори есть большое глазное пятно посередине тела, которое сбивает с толку добычу.
У Джона Дори есть большое глазное пятно посередине тела, которое сбивает с толку добычу.



Некоторые виды рыб также имеют ложные глазные пятна . Четырехглазая рыба-бабочка получила свое название из-за большого темного пятна на задней части каждой стороны тела. Это пятно окружено блестящим белым кольцом, напоминающим глазное пятно. Черная вертикальная полоса на голове проходит через настоящий глаз, из-за чего его трудно увидеть. [ 51 ] Это может привести к тому, что хищник подумает, что рыба больше, чем она есть на самом деле, и перепутает заднюю часть с передней. Первый инстинкт рыбы-бабочки при угрозе — бежать, помещая ложное глазное пятно ближе к хищнику, чем к голове. Большинство хищников целятся в глаза, и это ложное пятно заставляет хищника поверить, что рыба сначала убежит от хвоста.
Джон Дори — бентопелагическая прибрежная рыба с высоким сжатым с боков телом. Его тело настолько тонкое, что его едва можно увидеть спереди. У него также есть большое темное пятно с обеих сторон, которое используется для «сглаза», если приближается опасность. Большие глаза в передней части головы обеспечивают бифокальное зрение и восприятие глубины, необходимые для ловли добычи. Джона Дори Глазное пятно на боковой стороне тела также сбивает с толку добычу, которую затем засасывает в рот. [ 52 ]
Бочковые глаза
[ редактировать ]
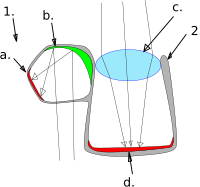
обычно направлен вверх, но может быть также повернут вперед
---------------------------------------------------------------------
Справа: Коричневорыла рыба-призрак — единственное известное позвоночное животное.
использовать зеркальный глаз (а также линзу):
(1) дивертикул (2) главный глаз
а) сетчатка (б) светоотражающие кристаллы (в) хрусталик (г) сетчатка
| Внешние видео | |
|---|---|
Баррелеи — это семейство маленьких, необычно выглядящих мезопелагических рыб, названных в честь бочкообразных трубчатых глаз, которые обычно направлены вверх, чтобы обнаружить силуэты доступной добычи. [ 53 ] [ 54 ] У баррелиев большие телескопические глаза, которые доминируют и выступают из черепа . Эти глаза обычно смотрят вверх, но у некоторых видов они также могут быть повернуты вперед. Их глаза имеют большой хрусталик и сетчатку с исключительным количеством палочек и высокой плотностью родопсина («зрительного пурпурного» пигмента); нет колбочек . [ 53 ]
Вид бочонка, Macropinna microstoma , имеет прозрачный защитный купол над макушкой, чем-то похожий на купол над кабиной самолета, через который можно увидеть линзы его глаз. Купол жесткий и гибкий и, по-видимому, защищает глаза от нематоцист (стрекательных клеток) сифонофоров, из которых, как полагают, бочонок крадет пищу. [ 53 ] [ 54 ] [ 55 ]
Другой вид бочкоглазых, коричнерылая рыба-призрак , является единственным известным позвоночным, которое использует зеркало, а не линзу, чтобы сфокусировать изображение в своих глазах. [ 56 ] [ 57 ] Он необычен тем, что для зрения использует как преломляющую , так и отражающую оптику. Главный трубчатый глаз содержит боковое овоидное вздутие, называемое дивертикулом , в значительной степени отделенное от глаза перегородкой . Сетчатка выстилает большую часть внутренней части глаза, и в роговице есть два отверстия, одно направлено вверх, а другое вниз, через которые свет попадает в главный глаз и дивертикул соответственно. В главном глазу, как и у других рыб, используется линза для фокусировки изображения. Однако внутри дивертикула свет отражается и фокусируется на сетчатке с помощью изогнутого составного зеркала, полученного из тапетума сетчатки , состоящего из множества слоев небольших отражающих пластинок, возможно, состоящих из гуанина кристаллов . Раздвоенная структура глаза буромордой рыбы-призрака позволяет рыбе видеть одновременно и вверх, и вниз. Кроме того, зеркальная система превосходит линзу по сбору света. Вероятно, главный глаз служит для обнаружения силуэтов объектов на фоне солнечного света, а дивертикул — для обнаружения биолюминесцентные вспышки с боков и снизу. [ 56 ]
Акулы
[ редактировать ]
акул Глаза похожи на глаза других позвоночных животных , включая аналогичные хрусталики , роговицу и сетчатку , хотя их зрение хорошо адаптировано к морской среде с помощью ткани, называемой Tapetum lucidum . Эта ткань находится за сетчаткой и отражает свет обратно к ней, тем самым увеличивая видимость в темной воде. Эффективность ткани варьируется: у некоторых акул более сильная адаптация к ночному образу жизни . Многие акулы могут сжимать и расширять свои зрачки , как и люди, чего не может сделать ни одна костистая рыба . У акул есть веки, но они не моргают, потому что окружающая вода очищает им глаза. Для защиты глаз у некоторых видов имеются мигательные перепонки . Эта мембрана закрывает глаза во время охоты и при нападении акулы. Однако некоторые виды, в том числе большая белая акула ( Carcharodon carcharias ), не имеют этой перепонки, а вместо этого закатывают глаза назад, чтобы защититься при ударе по добыче. Важность зрения в охотничьем поведении акул обсуждается. Некоторые считают, что электро- и хеморецепция более значимы, в то время как другие указывают на мигательную перепонку как на доказательство важности зрения. Предположительно, акула не стала бы защищать свои глаза, если бы они были неважны. Использование зрения, вероятно, зависит от вида и условий воды. Поле зрения акулы может переключаться между монокулярным и стереоскопическим . в любой момент [ 58 ] Микроспектрофотометрическое что исследование 17 видов акул показало, что у 10 из них были только палочковидные фоторецепторы и не было колбочек, в сетчатке обеспечивало им хорошее ночное зрение и в то же время делало их дальтониками . Остальные семь видов имели в дополнение к палочкам один тип колбочек-фоторецепторов, чувствительных к зеленому цвету, и, видя только оттенки серого и зеленого, считается, что они фактически страдают дальтонизмом. Исследование показывает, что контраст объекта на фоне, а не цвет, может быть более важным для обнаружения объекта. [ 59 ] [ 60 ] [ 61 ]
Другие примеры
[ редактировать ]


Мелкие рыбы часто собираются вместе в целях безопасности. Это может иметь визуальные преимущества, как за счет того, что хищные рыбы визуально сбивают с толку , так и за счет того, что косяк, рассматриваемый как тело, получает множество глаз. «Эффект замешательства хищника» основан на идее о том, что хищникам становится трудно выделять отдельную добычу из групп, поскольку множество движущихся целей создают сенсорную перегрузку зрительного канала хищника. [ 62 ] «Стайные рыбы одинакового размера и серебристого цвета, поэтому визуально ориентированному хищнику сложно выделить особь из массы извивающихся, сверкающих рыб и затем успеть схватить добычу до того, как она исчезнет в стае». [ 63 ] «Эффект многих глаз» основан на идее о том, что по мере увеличения размера группы задача сканирования окружающей среды на наличие хищников может быть распределена между многими особями, причем массовое сотрудничество, предположительно, обеспечивает более высокий уровень бдительности. [ 64 ] [ 65 ]
Рыбы обычно хладнокровны, их температура тела такая же, как и температура окружающей воды. Однако некоторые океанические хищные рыбы , такие как рыба-меч и некоторые виды акул и тунцов , могут согревать части своего тела, когда охотятся за добычей в глубокой и холодной воде. Рыба-меч с отличным зрением использует систему обогрева, задействующую ее мышцы, которая повышает температуру ее глаз и мозга на 15 °C. Нагревание сетчатки увеличивает скорость, с которой глаза реагируют на быстрые движения жертвы, почти в десять раз. [ 66 ] [ 67 ] [ 68 ]
У некоторых рыб есть блеск для глаз . [ 69 ] Блеск глаз является результатом работы светособирающего слоя в глазах, называемого тапетум люцидум , который отражает белый свет. У людей оно не встречается, но его можно увидеть у других видов, например, у оленя в свете фар. Блеск глаз позволяет рыбам хорошо видеть в условиях слабой освещенности, а также в мутной (с пятнами или бурной, бурной) воде, что дает им преимущество перед добычей. Это улучшенное зрение позволяет рыбе заселять более глубокие районы океана или озера. В частности, пресноводный судак назван так из-за блеска глаз. [ 70 ]
Многие виды Loricariidae , семейства сомовых , имеют видоизмененную радужку, называемую омега-ирисом . Верхняя часть радужной оболочки опускается, образуя петлю, которая может расширяться и сжиматься, называемую жаберной крышкой радужной оболочки; когда уровень освещенности высок, диаметр зрачка уменьшается, а петля расширяется, закрывая центр зрачка, образуя полумесяца . светопередающую часть в форме [ 71 ] Эта особенность получила свое название из-за сходства с перевернутой греческой буквой омега (Ом). Происхождение этой структуры неизвестно, но было высказано предположение, что разрушение контура хорошо видимого глаза помогает замаскировать животных, которые часто бывают сильно пестрыми. [ 71 ]
Дистанционные сенсорные системы
[ редактировать ]
Визуальные системы — это системы восприятия расстояния, которые предоставляют рыбе данные о местонахождении или объектах на расстоянии без необходимости непосредственного прикосновения к ним. Такие системы дистанционного зондирования важны, поскольку они позволяют общаться с другими рыбами и предоставляют информацию о местонахождении пищи и хищников, а также о том, как избегать препятствий или сохранять положение в косяках рыб . Например, у некоторых стайных видов на боках есть «стайные знаки», такие как визуально заметные полосы, которые служат ориентирами и помогают соседним рыбам судить об их относительном положении. [ 73 ] Но зрительная система – не единственная, способная выполнять подобные функции. У некоторых стайных рыб также есть боковая линия, проходящая по всей длине тела. Эта боковая линия позволяет рыбе чувствовать изменения давления воды и турбулентность вблизи ее тела. Используя эту информацию, стайная рыба может регулировать дистанцию до соседней рыбы, если она подойдет слишком близко или отойдет слишком далеко. [ 73 ]
Зрительная система рыб дополняется другими сенсорными системами с сопоставимыми или взаимодополняющими функциями. Некоторые рыбы слепы и должны полностью полагаться на альтернативные сенсорные системы. [ 74 ] Другие чувства, которые также могут предоставлять данные о местоположении или удаленных объектах, включают слух и эхолокацию , электрорецепцию , магнитоцепцию и хеморецепцию ( обоняние и вкус ). Например, у сомов хеморецепторы расположены по всему телу, что означает, что они «чувствуют вкус» всего, к чему прикасаются, и «нюхают» любые химические вещества в воде. «У сома вкус играет первостепенную роль в ориентации и расположении пищи». [ 75 ]
Хрящевые рыбы (акулы, скаты и химеры) используют магнитоцепцию. У них есть специальные электрорецепторы, называемые ампулами Лоренцини , которые улавливают небольшие изменения электрического потенциала. Эти рецепторы, расположенные вдоль рта и носа рыбы, действуют по принципу, согласно которому изменяющееся во времени магнитное поле, проходящее через проводник, индуцирует электрический потенциал на концах проводника. Ампулы также могут позволить рыбе обнаруживать изменения температуры воды. [ 76 ] [ 77 ] Как и у птиц, магнитоцепция может предоставлять информацию, которая помогает рыбам картировать маршруты миграции. [ 78 ]
См. также
[ редактировать ]- Глаз членистоногого
- соотношение Маттиссена
- Глаз моллюска
- Теменной глаз
- Простой глаз у беспозвоночных
- Визуальная система
Примечания
[ редактировать ]- ^ Мейер-Рохов, В. Бенно; Стюарт, Дункан (1996). «Обзор личиночной и постличиночной ультраструктуры глаз миноги (круглоротых) с особым акцентом на Geotria australis (серый)». Микроскопические исследования и техника . 35 (6): 431–444. doi : 10.1002/(SICI)1097-0029(19961215)35:6<431::AID-JEMT3>3.0.CO;2-L . ПМИД 9016447 . S2CID 22940203 .
- ^ Лэмб, Тревор Д.; Коллин, Шон П.; Пью, Эдвард Н. (2007). «Эволюция глаза позвоночных: опсины, фоторецепторы, сетчатка и глазная чашечка» . Обзоры природы Неврология . 8 (12): 960–976. дои : 10.1038/nrn2283 . ISSN 1471-003X . ПМК 3143066 . ПМИД 18026166 . См. также статью Ламба и др. «Происхождение глаза позвоночных», 2008 г.
- ^ Тревор Д. Лэмб; Шон П. Коллин; Эдвард Н. Пью-младший (2007). «Эволюция глаза позвоночных: опсины, фоторецепторы, сетчатка и глазная чашечка» . Обзоры природы Неврология . 8 (12): 960–976. дои : 10.1038/nrn2283 . ПМК 3143066 . ПМИД 18026166 .
- ^ Исследователь океана NOAA. Обновлено: 26 августа 2010 г.
- ^ Перейти обратно: а б с д и ж г час я дж к Хелфман и др. 2009 , стр. 84–87.
- ^ Перейти обратно: а б с Уэбб, Пол (2019) Введение в океанографию , глава 6.5 Свет, Сообщество Ребуса, Университет Роджера Уильямса, открытый учебник.
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Перейти обратно: а б с д Земля, МФ; Нильссон, Д. (2012). Глаза животных . Издательство Оксфордского университета. ISBN 9780199581146 .
- ^ Венер, Р. (2005). «Сенсорная физиология: безмозглые глаза» (PDF) . Природа . 435 (7039): 157–159. Бибкод : 2005Natur.435..157W . дои : 10.1038/435157a . ПМИД 15889076 . S2CID 4408533 .
- ^ Перейти обратно: а б Новалес Фламарик, Иньиго (2019). «Плавательное поведение рыбы настраивает поляризационное зрение рыбы, увеличивая вдвое дистанцию наблюдения за добычей» . Научные отчеты . 9 (1): 944. Бибкод : 2019НатСР...9..944Н . дои : 10.1038/s41598-018-37632-1 . ПМК 6353921 . ПМИД 30700806 .
- ^ Ингрэм, Норианн Т.; Сампат, Алапаккам П.; Фейн, Гордон Л. (2016). «Почему палочки более чувствительны, чем колбочки?» . Журнал физиологии . 594 (19): 5415–5426. дои : 10.1113/JP272556 . ПМК 5043029 . ПМИД 27218707 .
- ^ Маршалл, Джастин; Карлтон, Карен Л.; Кронин, Томас (2015). «Цветовое зрение морских организмов». Современное мнение в нейробиологии . 34 : 86–94. дои : 10.1016/j.conb.2015.02.002 . ПМИД 25725325 . S2CID 20978931 .
- ^ Миядзаки, Т; Иваму, Т; Мейер-Рохов, В.Б. (2011). «Положение центральной области сетчатки меняется с возрастом у Champsocephalus Gunnari (Channichthyidae), хищной рыбы из прибрежных вод Антарктики». Полярная биология . 34 (8): 1117–1123. дои : 10.1007/s00300-011-0969-2 . S2CID 19066809 .
- ^ Шваб, ИК; Харт, Н. (2006). «Больше, чем черно-белое» . Британский журнал офтальмологии . 90 (4): 406. doi : 10.1136/bjo.2005.085571 . ПМК 1857009 . ПМИД 16572506 .
- ^ Шваб, Иван Р. (2012) Свидетель эволюции: как развивались глаза, стр. 82. Oxford University Press. ISBN 9780195369748 .
- ^ Хоррамшахия, О; Шартауа, Дж. М.; Крегера, RHH (2008). «В глазах костистых рыб (костистых рыб) хрусталик удерживается на месте сложной системой связок и мышц» . Исследование зрения . 48 (13): 1503–1508. дои : 10.1016/j.visres.2008.03.017 . ПМИД 18471852 . S2CID 17757889 .
- ^ Сингх Х.Р. и Ханна С.С. (1994) Достижения в биологии рыб , стр. 235, паб «Хиндустан». ISBN 978-81-7075-029-1 .
- ^ Барнс, Греция (1 февраля 1979 г.). «Вестибулоокулярная функция при скоординированных движениях головы и глаз для захвата визуальных целей» . Журнал физиологии . 287 (1): 127–147. doi : 10.1113/jphysicalol.1979.sp012650 . ПМЦ 1281486 . ПМИД 311828 .
- ^ Граф, Вернер; Спенсер, Роберт; Бейкер, Харриет; Бейкер, Роберт (1 мая 1997 г.). «Возбуждающие и тормозные вестибулярные пути к экстраокулярным моторным ядрам золотых рыбок». Журнал нейрофизиологии . 77 (5): 2765–2779. дои : 10.1152/jn.1997.77.5.2765 . ПМИД 9163391 . S2CID 13004673 .
- ^ Граф, В.; Бейкер, Р. (1 октября 1985 г.). «Вестибулоокулярный рефлекс взрослой камбалы. II. Вестибулоокулярная связь». Журнал нейрофизиологии . 54 (4): 900–916. дои : 10.1152/jn.1985.54.4.900 . ПМИД 4067626 .
- ^ Перейти обратно: а б Граф, Вернер; Спенсер, Роберт; Бейкер, Харриет; Бейкер, Роберт (1 сентября 2001 г.). «Вестибулоокулярный рефлекс взрослой камбалы. III. Видоспецифическая реципрокная закономерность возбуждения и торможения». Журнал нейрофизиологии . 86 (3): 1376–1388. дои : 10.1152/jn.2001.86.3.1376 . ПМИД 11535684 .
- ^ Ёкояма, Сёдзо; Ёкояма, Рут (ноябрь 1996 г.). «Адаптивная эволюция фоторецепторов и зрительных пигментов у позвоночных». Ежегодный обзор экологии и систематики . 27 (1): 543–567. doi : 10.1146/annurev.ecolsys.27.1.543 .
- ^ Перейти обратно: а б Ши, Юншэн; Ёкояма, Сёдзо (8 июля 2003 г.). «Молекулярный анализ эволюционного значения ультрафиолетового зрения у позвоночных» . Труды Национальной академии наук . 100 (14): 8308–8313. Бибкод : 2003PNAS..100.8308S . дои : 10.1073/pnas.1532535100 . ПМК 166225 . ПМИД 12824471 .
- ^ Карлтон, Карен Л; Хароши, Ференц I; Кохер, Томас Д. (апрель 2000 г.). «Зрительные пигменты африканских цихлид: доказательства ультрафиолетового зрения на основе микроспектрофотометрии и последовательностей ДНК» . Исследование зрения . 40 (8): 879–890. дои : 10.1016/S0042-6989(99)00238-2 . ПМИД 10720660 . S2CID 5420659 .
- ^ Кодрик-Браун, Астрид; Джонсон, Салли К. (1 февраля 2002 г.). «Способы отражения ультрафиолета самцами гуппи повышают их привлекательность для самок». Поведение животных . 63 (2): 391–396. дои : 10.1006/anbe.2001.1917 . S2CID 53172856 .
- ^ Рик, Ингольф П.; Модарресси, Рикарда; Баккер, Тео CM (февраль 2006 г.). «Длина волны УФ-излучения влияет на выбор самки трехиглой колюшки». Поведение животных . 71 (2): 307–313. дои : 10.1016/j.anbehav.2005.03.039 . S2CID 937644 .
- ^ Джейкобс, Джеральд Х. (август 1992 г.). «Ультрафиолетовое зрение у позвоночных» . Американский зоолог . 32 (4): 544–554. дои : 10.1093/icb/32.4.544 .
- ^ Лоузи, Джордж С. (1 августа 2003 г.). «Крипсис и коммуникационные функции УФ-видимой окраски у двух коралловых рифовых стрекоз, Dascyllus aruanus и D.reticulatus *». Поведение животных . 66 (2): 299–307. дои : 10.1006/anbe.2003.2214 . S2CID 140204848 .
- ^ Зибек, Ульрике Э.; Паркер, Амира Н.; Шпренгер, Деннис; Метгер, Лидия М.; Уоллис, Гай (март 2010 г.). «Вид рифовых рыб, использующих ультрафиолетовые лучи для скрытого распознавания лиц» . Современная биология . 20 (5): 407–410. дои : 10.1016/j.cub.2009.12.047 . ПМИД 20188557 . S2CID 3743161 .
- ^ Хорват Г и Варью Д (2004) Поляризованный свет в зрении животных: закономерности поляризации в природе с. 294, Спрингер. ISBN 978-3-540-40457-6 .
- ^ Дентон, EJ ; Никол, JAC (1965). «Поляризация света, отраженного от серебристой внешности унылого Alburnus alburnus » (PDF) . Ж. Мар. Биол. доц. Великобритания 150 : 78–94.
- ^ Роу, DM; Дентон, Э.Дж. (1997). «Физическая основа рефлексивного общения между рыбами, в частности, ставрида Trachurus trachurus » . Фил. Пер. Р. Сок. Лонд. Б. 352 (1353): 531–549. Бибкод : 1997РСТБ.352..531Р . дои : 10.1098/rstb.1997.0037 . ПМК 1691948 .
- ^ Пиньятелли, В.; Чемп, К.; Маршалл, Дж.; Воробьев, М. (2010). «Двойные колбочки используются для распознавания цвета у рифовой рыбы Rhinecanthus aculeatus » . Письма по биологии . 6 (4): 537–539. дои : 10.1098/rsbl.2009.1010 . ПМК 2936199 . ПМИД 20129950 .
- ^ Фрёзе, Райнер; Поли, Дэниел (ред.) (2010). « Гигантура чуни » в FishBase . Версия за октябрь 2010 г.
- ^ Фрёзе, Райнер; Поли, Дэниел (ред.) (2009). « Dissostichus mawsoni » в FishBase . Версия за август 2009 года.
- ^ Нельсон, Джозеф, С. (2006). Рыбы мира . Джон Уайли и сыновья, Inc. ISBN 978-0-471-25031-9 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Фрёзе, Райнер; Поли, Дэниел (ред.) (2007). « Анаблепс анаблепс » в FishBase . Версия от марта 2007 г.
- ^ Мойл и Чех 2004 , с. 585.
- ^ Морин, Джеймс Г.; Харрингтон, Энн; Нилсон, Кеннет; Кригер, Нил; Болдуин, Томас О.; Гастингс, JW (1975). «Свет для всех причин: универсальность поведенческого репертуара рыбы-фонарика». Наука . 190 (4209): 74–76. Бибкод : 1975Науч...190...74М . дои : 10.1126/science.190.4209.74 . S2CID 83905458 .
- ^ Маккоскер, Джон Э. (март 1977 г.). «Рыбки-фонарики». Научный американец . 236 (3): 106–114. Бибкод : 1977SciAm.236c.106M . doi : 10.1038/scientificamerican0377-106 . JSTOR 24953941 . ПМИД 841297 .
- ^ Пакстон, Джон Р. (1998). Пакстон, младший; Эшмейер, WN (ред.). Энциклопедия рыб . Сан-Диего: Академическая пресса. п. 162. ИСБН 978-0-12-547665-2 .
- ^ Райан П. «Глубоководные существа: Батипелагическая зона» Те Ара - Энциклопедия Новой Зеландии . Обновлено 21 сентября 2007 г.
- ^ Мойл и Чех 2004 , с. 587.
- ^ Шапло, Франсуа и Амаока, Кунио (1998). Пакстон, младший и Эшмейер, WN (ред.). Энциклопедия рыб Сан-Диего: Академическая пресса. ххх. ISBN 978-0-12-547665-2 .
- ^ Докинз, Ричард (1991). Слепой часовщик . Лондон: Книги Пингвина. п. 92. ИСБН 978-0-14-014481-9 .
- ^ Кенали, CP (2007). «Пересмотр рода Stoplight Loosejaw Malacosteus (Teleostei: Stomiidae: Malacosteinae) с описанием нового вида из умеренного южного полушария и Индийского океана». Копейя . 2007 (4): 886–900. doi : 10.1643/0045-8511(2007)7[886:ROTSLG]2.0.CO;2 . S2CID 1038874 .
- ^ «Хищники» . Министерство внутренних дел США, Бюро землеустройства. 14 декабря 2009 года. Архивировано из оригинала 14 июня 2011 года . Проверено 28 марта 2011 г.
- ^ Бородицкий, Лера (24 июня 1999 г.). «Свет и глаза: конспект лекций» . Конспекты лекций . Стэнфорд . Архивировано из оригинала 5 июля 2010 года . Проверено 11 мая 2010 г.
- ^ Countershading BBC: Наука и природа . Проверено 28 сентября 2011 г.
- ^ Рыбные друзья и рыбные враги. Руководство по подготовке, Морской институт Лонг-Бич .
- ^ Клаас, Жюльен М.; Акснес, Даг Л.; Маллефет, Жером (май 2010 г.). «Охотник-призрак фьордов: камуфляж с помощью контриллюминации у акулы (Etmopterus spinax)». Журнал экспериментальной морской биологии и экологии . 388 (1–2): 28–32. дои : 10.1016/j.jembe.2010.03.009 .
- ^ FishBase Фрёзе, Райнер; Поли, Дэниел (ред.) (2009). « Chaetodon capistratus » в FishBase . Версия за июль 2009 года.
- ^ Уолронд, Карл (2006) Прибрежные рыбы — Рыбы дна открытого моря , Те Ара: Энциклопедия Новой Зеландии . По состоянию на 28 мая 2019 г.
- ^ Перейти обратно: а б с Робисон, Брюс Х.; Райзенбихлер, Ким Р. (18 декабря 2008 г.). «Микростома Macropinna и парадокс ее трубчатых глаз». Копейя . 2008 (4): 780–784. дои : 10.1643/CG-07-082 . S2CID 85768623 .
- ^ Перейти обратно: а б Робисон, Брюс Х.; Райзенбихлер, Ким Р. (18 декабря 2008 г.). «Микростома Macropinna и парадокс ее трубчатых глаз». Копейя . 2008 (4): 780–784. дои : 10.1643/CG-07-082 . JSTOR 25512162 . S2CID 85768623 . ПроКвест 207224476 .
- «Исследователи разгадали тайну глубоководной рыбы с трубчатыми глазами и прозрачной головой» . Научно-исследовательский институт аквариумов Монтерей-Бэй (пресс-релиз). 23 февраля 2009 г.
- ^ Фрёзе, Райнер; Поли, Дэниел (ред.) (2011). « Макропинна микростома » в FishBase . Версия за сентябрь 2011 года.
- ^ Перейти обратно: а б Вагнер, Ханс-Иоахим; Дуглас, Рон Х.; Франк, Тамара М.; Робертс, Николас В.; Партридж, Джулиан К. (январь 2009 г.). «Новый глаз позвоночных, использующий как преломляющую, так и отражающую оптику» . Современная биология . 19 (2): 108–114. дои : 10.1016/j.cub.2008.11.061 . ПМИД 19110427 . S2CID 18680315 .
- ^ Репортер Льюис Смит, «Окружающая среда» (8 января 2009 г.). «Рыба с четырьмя глазами может видеть сквозь глубокий морской мрак» . Таймс .
{{cite news}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Мартин, Р. Эйдан. «Видение и ковер света» . Центр исследований акул ReefQuest . Проверено 22 августа 2009 г.
- ^ Харт, Натан Скотт; Тайсс, Сьюзен Мишель; Харахуш, Блейк Кристин; Коллин, Шон Патрик (март 2011 г.). «Микроспектрофотометрические доказательства монохромности конуса у акул». Naturwissenschaften . 98 (3): 193–201. Бибкод : 2011NW.....98..193H . дои : 10.1007/s00114-010-0758-8 . ПМИД 21212930 . S2CID 30148811 .
- «Новые исследования показали, что акулы не различают цвета» . Австралийское географическое издание . 19 января 2011 г.
- ^ Гилл, Виктория (18 января 2011 г.). «Акулы, вероятно, дальтоники» . Новости Би-би-си . Проверено 19 января 2011 г.
- ^ Харт, Натан Скотт; Тайсс, Сьюзен Мишель; Харахуш, Блейк Кристин; Коллин, Шон Патрик (март 2011 г.). «Микроспектрофотометрические доказательства монохромности конуса у акул». Naturwissenschaften . 98 (3): 193–201. Бибкод : 2011NW.....98..193H . дои : 10.1007/s00114-010-0758-8 . ПМИД 21212930 . S2CID 30148811 .
- ^ Милински, Манфред; Хеллер, Рольф (октябрь 1978 г.). «Влияние хищника на оптимальное пищевое поведение колюшки (Gasterosteus aculeatus L.)». Природа . 275 (5681): 642–644. Бибкод : 1978Natur.275..642M . дои : 10.1038/275642a0 . S2CID 4184043 .
- ^ Мойл и Чех 2004 , с. [ нужна страница ] .
- ^ Робертс, Гилберт (май 1996 г.). «Почему индивидуальная бдительность снижается по мере увеличения размера группы». Поведение животных . 51 (5): 1077–1086. дои : 10.1006/anbe.1996.0109 . S2CID 53202810 .
- ^ Лима, Стивен Л. (январь 1995 г.). «Назад к основам бдительности против хищников: эффект размера группы». Поведение животных . 49 (1): 11–20. дои : 10.1016/0003-3472(95)80149-9 . S2CID 53205760 .
- ^ Фричес, Керстин А.; Брилл, Ричард В.; Ордер, Эрик Дж. (январь 2005 г.). «Теплые глаза обеспечивают превосходное зрение у рыб-меч» . Современная биология . 15 (1): 55–58. дои : 10.1016/j.cub.2004.12.064 . ПМИД 15649365 . S2CID 14070646 .
- ^ Хопкин, Майкл (10 января 2005 г.). «Рыба-меч нагревает глаза для лучшего зрения». Природа : news050110–2. дои : 10.1038/news050110-2 .
- ^ Хелфман и др. 2009 , стр. 95–97.
- ^ Сомия, Х (1980). «Рыбы с блеском глаз: функциональная морфология гуанинового типа Tapetum Lucidum» . Серия «Прогресс в области морской экологии» . 2 : 9–26. Бибкод : 1980MEPS....2....9S . дои : 10.3354/meps002009 .
- ^ Джонсон Дж.А. и Эссер Р. (2009) « http://www.fishculturalsection.org/Aquanotes/pdf/Aq_App_Note_1_April_2009.pdf Разведение судака – привычка кормиться в темноте» Американское общество рыболовства , Рекомендации по применению аквакультуры.
- ^ Перейти обратно: а б Дуглас, Рон Х.; Коллин, Шон П.; Корриган, Джули (15 ноября 2002 г.). «Глаза панцирного сома-присоски (Loricariidae, подсемейство Hypostomus): реакция зрачка, двояковыпуклая продольная сферическая аберрация и топография сетчатки» . Журнал экспериментальной биологии . 205 (22): 3425–3433. дои : 10.1242/jeb.205.22.3425 . ПМИД 12364396 .
- ^ Ёсидзава, Масато; Ямамото, Ёсиюки; О'Куин, Келли Э; Джеффри, Уильям Р. (декабрь 2012 г.). «Эволюция адаптивного поведения и его сенсорных рецепторов способствует регрессии глаз у слепых пещерных рыб» . БМК Биология . 10 (1): 108. дои : 10.1186/1741-7007-10-108 . ПМЦ 3565949 . ПМИД 23270452 .
- ^ Перейти обратно: а б Боун и Мур 2008 , стр. 418–422.
- ^ Боун и Мур 2008 , с. 311.
- ^ Атема, Джелле (1980) «Химические чувства, химические сигналы и пищевое поведение рыб» с. 57–101. В: Бардах, Дж. Э. Поведение рыбы и его использование в отлове и выращивании рыб, The WorldFish Center, ISBN 978-971-02-0003-0 .
- ^ Филдс, Р. Дуглас; Филдс, Кайл Д.; Филдс, Мелани К. (октябрь 2007 г.). «Полупроводниковый гель в органах чувств акулы?» . Письма по неврологии . 426 (3): 166–170. дои : 10.1016/j.neulet.2007.08.064 . ПМК 2211453 . ПМИД 17904741 .
- ^ Браун, Брэндон Р. (март 2010 г.). «Температурный отклик электросенсоров и тепловые напряжения в электролитах» . Журнал биологической физики . 36 (2): 121–134. дои : 10.1007/s10867-009-9174-8 . ПМЦ 2825305 . ПМИД 19760113 .
- ^ Йонсен, Зёнке; Ломанн, Кеннет Дж. (сентябрь 2005 г.). «Физика и нейробиология магниторецепции». Обзоры природы Неврология . 6 (9): 703–712. дои : 10.1038/nrn1745 . ПМИД 16100517 . S2CID 13996233 .
Ссылки
[ редактировать ]- Боун, Квентин; Мур, Ричард (2008). Биология рыб . Гирляндная наука. ISBN 978-0-203-88522-2 .
- Хельфман, Джин; Коллетт, Брюс Б.; Фейси, Дуглас Э.; Боуэн, Брайан В. (2009). Разнообразие рыб: биология, эволюция и экология . Джон Уайли и сыновья. ISBN 978-1-4443-1190-7 .
- Мойл, Питер Б.; Чех, Джозеф Дж. (2004). Рыбы: введение в ихтиологию . Пирсон Прентис Холл. ISBN 978-0-13-100847-2 .
Дальнейшее чтение
[ редактировать ]- Артур, Джозеф; Никол, Колин; Сомия, Хироаки (1989). Глаза рыб . Кларендон Пресс. ISBN 978-0-19-857195-7 .
- Дуглас Р.Х. и Джамгоз М. (редакторы) (1990) Зрительная система рыб . Чепмен и Холл, 526 стр.
- Лэмб, Тревор Д. (14 июня 2011 г.). «Эволюция глаза». Научный американец . 305 (1): 64–69. Бибкод : 2011SciAm.305f..64L . doi : 10.1038/scientificamerican0711-64 .
- Лэнд, Майкл Ф. и Нильссон, Дэн-Эрик (2012) Animal Eyes Oxford University Press. ISBN 9780199581146 .
- Лэмб, Тревор Д.; Коллин, Шон П.; Пью, Эдвард Н. (декабрь 2007 г.). «Эволюция глаза позвоночных: опсины, фоторецепторы, сетчатка и глазная чашечка» . Обзоры природы. Нейронаука . 8 (12): 960–976. дои : 10.1038/nrn2283 . ПМК 3143066 . ПМИД 18026166 .
- «Следить за эволюцией» . Физика.орг . 3 декабря 2007 г.
- Нильссон, Делавэр; Пелгер, С. (22 апреля 1994 г.). «Пессимистическая оценка времени, необходимого для развития глаза». Труды Лондонского королевского общества. Серия Б: Биологические науки . 256 (1345): 53–58. Бибкод : 1994РСПСБ.256...53Н . дои : 10.1098/rspb.1994.0048 . ПМИД 8008757 . S2CID 13061351 .
- Берлински, Дэвид (2002) Встретил ли Дарвин свою пару? Страница 34, Досадный глаз (Письмо). Комментарий , 1 декабря 2002 г.
- Нильссон, Дэн-Э. «Остерегайтесь псевдонауки: ответ на нападки Дэвида Берлински на мои расчеты того, сколько времени требуется глазу, чтобы развиться» . Разговор о причине .
- Мейер-Рохов, Виктор Бенно; Коддингтон, Пол Эдвард (2003). «Глаза и зрение новозеландской торрент-рыбы Cheimarrichthys способствуют ФОН ХААСТУ (1874 г.): Гистология, фотохимия и электрофизиология». В Вале — Адальберто Луис; Капур, Б.Г. (ред.). Адаптации рыб . Научные издательства. стр. 337–383. ISBN 978-1-57808-249-0 .
- «Эволюция глаза» - видео о модели Нильссона-Пельгера (прокрутите вниз)
Библиография
[ редактировать ]- Маршалл, Джастин; Карлтон, Карен Л; Кронин, Томас (октябрь 2015 г.). «Цветовое зрение морских организмов». Современное мнение в нейробиологии . 34 : 86–94. дои : 10.1016/j.conb.2015.02.002 . ПМИД 25725325 . S2CID 20978931 .
- Камидзё, Макико; Кавамура, Маюко; Фукамати, Сёдзи (май 2018 г.). «Потеря генов красного опсина ослабляет сексуальную изоляцию между вариантами медаки цвета кожи». Поведенческие процессы . 150 : 25–28. дои : 10.1016/j.beproc.2018.02.006 . ПМИД 29447852 . S2CID 4239046 .

