Дождевой червь
| Дождевой червь Временной диапазон:
| |
|---|---|

| |
| Неопознанный вид дождевых червей с хорошо развитым клитором. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Аннелиды |
| Клад : | Плейстоаннелида |
| Клад : | Сидячий |
| Сорт: | Клителлатный |
| Заказ: | Опистопора |
| Подотряд: | Лумбричина |
червь Дождевой — в почве , обитающее наземное беспозвоночное животное и принадлежащее к типу Annelida . Термин — общее название крупнейших представителей класса ( или подкласса , в зависимости от автора) Oligochaeta . В классических системах они относились к отряду Opisthopora , поскольку мужские поры открывались позади женских, хотя внутренние мужские сегменты располагались впереди женских. Теоретические кладистические исследования отнесли их к подотряду Lumbricina отряда Haplotaxida , но ситуация может измениться. [ нужны разъяснения ] Другие сленговые названия дождевых червей включают «росовый червь», «дождевой червь», «ночной змей» и «угловой червь» (из-за его использования в качестве для рыбалки наживки ). Более крупных наземных червей также называют мегадрилами (что переводится как «большие черви») в отличие от микродрилов («маленьких червей») полуводных семейств Tubificidae , Lumbricidae и Enchytraeidae . Мегадрилы характеризуются отчетливым клитором (более обширным, чем у микродрилей) и сосудистой системой с настоящими капиллярами . [ 2 ]
Дождевые черви обычно обитают во влажной, богатой компостом почве, питаясь разнообразными органическими веществами . [ 3 ] в состав которых входят детрит , живые простейшие , коловратки , нематоды , бактерии , грибы и другие микроорганизмы . [ 4 ] Пищеварительная система дождевого червя проходит по всей длине его тела. [ 5 ] природы Они являются одними из наиболее важных детритофагов и копрофагов , а также служат пищей низкого уровня для многих потребителей в экосистемах.
«трубка внутри трубки» Дождевые черви имеют внешне сегментированное тело с соответствующими внутренними сегментациями и обычно имеют щетинки на всех сегментах. [ 6 ] Они имеют космополитическое распространение везде, где позволяют почвенные, водные и температурные условия. [ 7 ] У них есть двойная транспортная система, состоящая из целомической жидкости , которая движется внутри заполненного жидкостью целома , и простая замкнутая система кровообращения , и они дышат (дышат) посредством кожного дыхания . Как беспозвоночные с мягким телом, у них отсутствует настоящий скелет , но их структура поддерживается заполненными жидкостью камерами целома, которые функционируют как гидростатический скелет . [ нужна ссылка ]
У дождевых червей есть центральная нервная система, состоящая из двух ганглиев над ртом , по одному с каждой стороны, соединенных с осевым нервом , идущим по всей длине к двигательным нейронам и сенсорным клеткам в каждом сегменте. большое количество хеморецепторов Около рта концентрируется . Окружные и продольные мышцы, окаймляющие каждый сегмент, позволяют червю двигаться. Похожие наборы мышц выстилают кишечную трубку червя , и их действия продвигают переваренную пищу к анусу . [ 8 ]
Дождевые черви — гермафродиты : каждый червь несет мужские и женские репродуктивные органы и половые поры . При спаривании два отдельных дождевых червя обмениваются спермой друг друга и оплодотворяют яйцеклетки .
Анатомия
Форма и функция
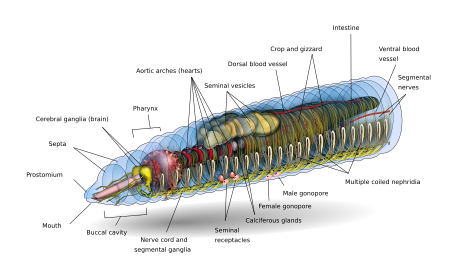
В зависимости от вида взрослый дождевой червь может иметь длину от 10 мм (0,39 дюйма) и ширину 1 мм (0,039 дюйма) до 3 м (9,8 фута) в длину и ширину более 25 мм (0,98 дюйма), но типичный Lumbricus terrestris растет примерно до 360 мм (14 дюймов) в длину. [ 9 ] Вероятно, самым длинным червем из подтвержденных записей является Amynthas mekongianus , длина которого достигает 3 м (10 футов). [ 10 ] в грязи вдоль берегов реки Меконг длиной 4350 км (2703 миль) в Юго-Восточной Азии.
Основная форма дождевого червя спереди назад представляет собой цилиндрическую трубку в трубке, разделенную на ряд сегментов (называемых метамерами ), которые разделяют тело на отдельные сегменты. Борозды, как правило, [ 11 ] внешне заметные на теле разграничения сегментов; спинные поры и нефридиопоры выделяют жидкость, которая увлажняет и защищает поверхность червя, позволяя ему дышать. За исключением ротового и анального сегментов, каждый сегмент несет щетинообразные волоски, называемые боковыми щетинками. [ 12 ] используется для фиксации частей тела во время движения; [ 13 ] виды могут иметь четыре пары щетинок на каждом сегменте или более восьми, иногда образуя полный круг щетинок на сегменте. [ 12 ] Специальные брюшные щетинки используются для закрепления спаривающихся дождевых червей при их проникновении в тела партнеров. [ 14 ]
Как правило, внутри вида количество обнаруженных сегментов одинаково у разных особей, и особи рождаются с тем количеством сегментов, которое у них будет на протяжении всей жизни. Первый сегмент тела (сегмент номер 1) включает в себя как рот дождевого червя, так и нависающую над ним мясистую долю, называемую простомиумом , которая закрывает вход, когда червь находится в состоянии покоя, но также используется для ощущения и химического ощущения окружения червя. . Некоторые виды дождевых червей могут даже использовать цепкую простомиум, чтобы хватать и тащить в свою нору такие предметы, как трава и листья.
У взрослого дождевого червя появляется ремневидная железистая опухоль, называемая клитором , которая покрывает несколько сегментов в передней части животного. Это часть репродуктивной системы, производящая яйцевые капсулы. Задняя часть чаще всего цилиндрическая, как и остальная часть тела, но в зависимости от вида она также может быть четырехугольной, восьмиугольной, трапециевидной или сплющенной. Последний сегмент называется перипроктом ; На этом сегменте находится анус дождевого червя — короткая вертикальная щель. [ 12 ]

Внешняя часть отдельного сегмента представляет собой тонкую кутикулу над кожей, обычно пигментированную от красного до коричневого цвета, которая имеет специализированные клетки, которые выделяют слизь поверх кутикулы, чтобы поддерживать тело влажным и облегчать движение по почве. Под кожей находится слой нервной ткани и два слоя мышц: тонкий внешний слой кольцевых мышц и гораздо более толстый внутренний слой продольных мышц. [ 15 ] Внутри мышечного слоя находится заполненная жидкостью камера, называемая целомом. [ 16 ] что за счет давления обеспечивает структуру бескостного тела червя. Сегменты отделены друг от друга перегородками (множественное число от слова «перегородка»). [ 17 ] которые представляют собой перфорированные поперечные стенки, позволяющие целомической жидкости проходить между сегментами. [ 18 ] Пара структур, называемых нефростомами, расположена в задней части каждой перегородки; от каждого нефростома через перегородку в следующий сегмент идет нефрический каналец. Затем этот каналец ведет к основному органу фильтрации жидкостей организма, нефридию или метанефридию, который удаляет метаболические отходы из целомической жидкости и вытесняет их через поры, называемые нефридиопорами, на боках червя; обычно в большинстве сегментов обнаруживаются две нефридии (иногда и больше). [ 19 ] В центре червя находится пищеварительный тракт , который проходит прямо ото рта до ануса, не скручиваясь, и окружен сверху и снизу кровеносными сосудами (дорсальным кровеносным сосудом и вентральным кровеносным сосудом, а также субневральным кровеносным сосудом) и вентральный нервный шнур и в каждом сегменте окружен парой паллиальных кровеносных сосудов, которые соединяют дорсальные и субневральные кровеносные сосуды.
Многие дождевые черви могут выбрасывать целомическую жидкость через поры на спине в ответ на стресс; Австралийский Didymogaster sylvaticus (известный как «синий дождевой червь») может выпрыскивать жидкость на высоту до 30 см (12 дюймов). [ 20 ] [ 18 ]
Нервная система

Центральная нервная система
ЦНС состоит из двудольного головного мозга (церебральных ганглиев , или надглоточных ганглиев), подглоточных ганглиев, окологлоточных коннектив и вентрального нервного тяжа .
Мозг дождевых червей состоит из пары мозговых ганглиев грушевидной формы. Они расположены на дорсальной стороне пищеварительного канала в третьем сегменте, в борозде между полостью рта и глоткой .
Пара окологлоточных связок головного мозга окружает глотку и затем соединяется с парой подглоточных ганглиев, расположенных ниже глотки в четвертом сегменте. Такое расположение означает, что мозг, подглоточные ганглии и окологлоточные соединительные ткани образуют нервное кольцо вокруг глотки.
Вентральный нервный тяж (образованный нервными клетками и нервными волокнами) начинается от подглоточных ганглиев и простирается ниже пищеварительного канала до самого заднего сегмента тела. Брюшной нервный тяж в каждом сегменте имеет вздутие, или ганглий, т. е. сегментарный ганглий, возникающий от пятого до последнего сегмента тела. также имеются три гигантских аксона На средне-дорсальной стороне вентрального нервного канатика , один медиальный гигантский аксон (MGA) и два латеральных гигантских аксона (LGA). MGA имеет диаметр 0,07 мм и передается в передне-заднем направлении со скоростью 32,2 м/с. LGA немного уже, их диаметр составляет 0,05 мм, а передача осуществляется в задне-переднем направлении со скоростью 12,6 м/с. Два LGA соединены вдоль тела через равные промежутки времени и поэтому считаются одним гигантским аксоном. [ 21 ] [ 22 ]
Периферическая нервная система
- От церебральных ганглиев отходят от восьми до десяти нервов, иннервирующих простомиум , щечную камеру и глотку .
- Три пары нервов отходят от подглоточных ганглиев и иннервируют второй, третий и четвертый сегменты.
- отходят три пары нервов, От каждого сегментарного ганглия иннервирующие различные структуры сегмента.
Симпатическая нервная система состоит из нервных сплетений в эпидермисе и пищеварительном тракте. (Сплетение представляет собой сеть связанных между собой нервных клеток.) Нервы, идущие вдоль стенки тела, проходят между наружным циркулярным и внутренним продольным мышечными слоями стенки. Они отдают ветви, образующие межмышечное и субэпидермальное сплетения. Эти нервы соединяются с перстнеглоточной соединительной тканью.
Движение

На первый взгляд, скорость ползания варьируется как внутри отдельных особей, так и между ними. Дождевые черви ползают быстрее, в первую очередь за счет более длинных «шагов» и большей частоты шагов. Более крупные черви Lumbricus terrestris ползают с большей абсолютной скоростью, чем более мелкие черви. Они достигают этого, делая немного более длинные шаги, но с немного более низкой частотой шагов. [ 23 ]
Прикосновение к дождевому червю, вызывающее реакцию «давления», а также (часто) реакцию на обезвоживающее действие соли на кожу человека (токсичной для дождевых червей), стимулирует субэпидермальное нервное сплетение, которое соединяется с межмышечным сплетением и вызывает продольное мышцы сокращаться. Это вызывает извивающиеся движения, наблюдаемые, когда человек берет в руки дождевого червя. Такое поведение является рефлексом и не требует участия ЦНС; это происходит, даже если нервный канатик удален. Каждый сегмент дождевого червя имеет собственное нервное сплетение. Сплетение одного сегмента не связано непосредственно со сплетением соседних сегментов. Нервный шнур необходим для соединения нервных систем сегментов. [ 24 ]
Гигантские аксоны передают самые быстрые сигналы по нервному канатику. Это сигналы чрезвычайной ситуации, которые инициируют рефлекторное поведение, связанное с бегством. Более крупный дорсальный гигантский аксон проводит сигналы быстрее всего: от задней части животного к передней. Если коснуться задней части червя, сигнал быстро посылается вперед, вызывая сокращение продольных мышц в каждом сегменте. Это приводит к тому, что червь очень быстро сокращается при попытке убежать от хищника или другой потенциальной угрозы. Два медиальных гигантских аксона соединяются друг с другом и посылают сигналы спереди назад. Их стимуляция заставляет дождевого червя очень быстро отступать (возможно, сжимаясь в норе, спасаясь от птицы).
Наличие нервной системы необходимо для того, чтобы животное могло испытывать ноцицепцию или боль . Однако необходимы и другие физиологические способности, такие как чувствительность к опиоидам и центральная модуляция ответов анальгетиками. [ 25 ] энкефалин и α-эндорфинподобные У дождевых червей обнаружены вещества. Инъекции налоксона (антагониста опиоидов) подавляют реакцию побега дождевых червей. Это указывает на то, что опиоидные вещества играют роль в сенсорной модуляции, аналогичной той, которая обнаружена у многих позвоночных. [ 26 ]
Сенсорный прием
Светочувствительность
Хотя у некоторых червей есть глаза , у дождевых червей их нет. Однако у них есть специализированные светочувствительные клетки, называемые «световыми клетками Гесса». Эти фоторецепторные клетки имеют центральную внутриклеточную полость ( фаосому ), заполненную микроворсинками . Помимо микроворсинок, в фаосоме имеется несколько сенсорных ресничек, структурно независимых от микроворсинок. [ 27 ] Фоторецепторы расположены на большей части эпидермиса, но более сконцентрированы на спине и боках червя. Относительно небольшое их количество встречается на вентральной поверхности первого сегмента. Они наиболее многочисленны в простомиуме и уменьшаются в плотности в первых трех сегментах; за пределами третьего сегмента их очень мало. [ 24 ]
Эпидермальный рецептор (орган чувств)
Эти рецепторы многочисленны и распределены по всему эпидермису . Каждый рецептор имеет слегка приподнятую кутикулу, которая покрывает группу высоких, тонких и столбчатых рецепторных клеток. Эти клетки несут на внешних концах небольшие волосообразные отростки, а внутренние концы связаны с нервными волокнами. Эпидермальные рецепторы выполняют тактильную функцию. Они также обеспокоены изменениями температуры и реагируют на химические раздражители. Дождевые черви чрезвычайно чувствительны к прикосновениям и механической вибрации.
Буккальный рецептор (орган чувств)
Эти рецепторы расположены только в эпителии щечной камеры. Эти рецепторы являются вкусовыми и обонятельными (связанными со вкусом и запахом). Они также реагируют на химические раздражители. (хеморецептор)
Пищеварительная система
Кишечник дождевого червя представляет собой прямую трубку, идущую от рта червя к его анусу . Он дифференцируется на пищеварительный канал и связанные с ним железы, которые встроены в стенку самого пищеварительного канала. Пищеварительный канал состоит из рта, ротовой полости (обычно проходит через один или два первых сегмента дождевого червя), глотки (обычно имеет длину около четырех сегментов), пищевода, зоба, желудка (обычно) и кишечника. [ 28 ]
Еда попадает через рот. Глотка ; действует как всасывающий насос его мускулистые стенки всасывают пищу. В глотке глоточные железы выделяют слизь . Пища попадает в пищевод , куда кальций закачивается (из крови и поступивший с предыдущим приемом пищи) для поддержания надлежащего уровня кальция в крови и pH пищи . Оттуда пища попадает в зоб и желудок. В желудке сильные мышечные сокращения перемалывают пищу с помощью минеральных частиц, попавших вместе с пищей. Пройдя через желудок, пища продолжает двигаться по кишечнику для переваривания. Кишечник секретирует пепсин для переваривания белков, амилазу для переваривания полисахаридов, целлюлазу для переваривания целлюлозы и липазу для переваривания жиров. [ 8 ] В дополнение к пищеварительным белкам дождевые черви используют класс поверхностно-активных соединений, называемых дрилодефензинами , которые помогают переваривать растительный материал. [ 29 ] Вместо того, чтобы быть свернутым, как кишечник млекопитающего, в кишечнике дождевого червя присутствует большая средне-спинная, похожая на язык складка, называемая тифлозолем , со множеством складок, идущих по ее длине, увеличивающих площадь ее поверхности для увеличения поглощения питательных веществ. Кишечник имеет свою пару мышечных слоев, как и тело, но в обратном порядке: внутренний циркулярный слой внутри внешнего продольного слоя. [ 30 ]
Кровеносная система
Дождевые черви имеют двойную систему кровообращения, в которой как целомическая жидкость, так и закрытая система кровообращения переносят пищу, отходы и дыхательные газы. Закрытая кровеносная система имеет пять основных кровеносных сосудов: спинной (верхний) сосуд, проходящий над пищеварительным трактом; вентральный (нижний) сосуд, проходящий ниже пищеварительного тракта; субневральный сосуд, проходящий ниже брюшного нервного канатика; и два латероневральных сосуда по обе стороны нервного канатика. [ 31 ]
Спинной сосуд представляет собой главным образом собирательную структуру в области кишечника. В каждом сегменте он получает парную комиссуральную и дорсальную кишки. В каждом сегменте вентральный сосуд разветвляется на пару вентро-тегументариев и вентро-кишечных костей. Субневральный сосуд также отдает пару комиссур, идущих по задней поверхности перегородки.
Насосное действие дорсального сосуда перемещает кровь вперед, в то время как другие четыре продольных сосуда несут кровь назад. В сегментах с седьмого по одиннадцатый пара дуг аорты окружает целом и действует как сердце, перекачивая кровь в вентральный сосуд, который действует как аорта. Кровь состоит из амебоидных клеток и гемоглобина, растворенного в плазме. Вторая система кровообращения происходит от клеток пищеварительной системы, выстилающих целом. Когда пищеварительные клетки наполняются, они выделяют неживые клетки жира в заполненный жидкостью целом, где они свободно плавают, но могут проходить сквозь стенки, разделяющие каждый сегмент, перемещая пищу в другие части и способствуя заживлению ран. [ 32 ]
Выделительная система
Выделительная система содержит по паре нефридиев в каждом сегменте, кроме первых трех и последнего. [ 33 ] Выделяют три типа нефридий: покровные, перегородочные и глоточные. Покровные нефридии прикрепляются к внутренней стороне стенки тела во всех сегментах, кроме первых двух. Перегородочные нефридии прикрепляются к обеим сторонам перегородки позади 15-го сегмента. Глоточные нефридии прикрепляются к четвертому, пятому и шестому сегментам. [ 33 ] переднего сегмента втягиваются за счет сокращения ресничек нефростома Отходы целомической жидкости из . Оттуда он проводится через перегородку (стенку) по трубке, которая образует ряд петель, оплетенных кровеносными капиллярами, которые также переносят отходы в каналец нефростома. Затем экскреторные отходы выводятся через пору на стороне червя. [ 34 ]
Дыхание
Специальных органов дыхания у дождевых червей нет. Газообмен осуществляется через влажную кожу и капилляры, где кислород подхватывается растворенным в плазме крови гемоглобином и выделяется углекислый газ. Вода, как и соли, также может перемещаться через кожу путем активного транспорта.
Жизнь и физиология
При рождении дождевые черви появляются маленькими, но полностью сформированными, у них отсутствуют только половые структуры, которые развиваются примерно через 60–90 дней. Полного размера они достигают примерно за год. Ученые прогнозируют, что средняя продолжительность жизни в полевых условиях составляет четыре-восемь лет, тогда как большинство садовых сортов живут всего один-два года.
Воспроизведение



Некоторые распространенные виды дождевых червей в основном являются партеногенетическими , что означает, что рост и развитие эмбрионов происходит без оплодотворения . У люмбрицидных дождевых червей партеногенез неоднократно возникал от половых родственников. [ 35 ] Партеногенез у некоторых линий Aporrectodea трапецоидес возник от 6,4 до 1,1 миллиона лет назад от половых предков. [ 36 ] У некоторых видов наблюдается псевдогамный партогенез, а это означает, что для стимуляции воспроизводства необходимо спаривание, даже если потомству не передается мужской генетический материал. [ 37 ]
Спаривание дождевых червей происходит на поверхности, чаще всего ночью. Дождевые черви — гермафродиты ; то есть у них есть как мужские, так и женские половые органы. Половые органы расположены в сегментах с 9 по 15. У дождевых червей есть одна или две пары семенников, находящихся внутри мешков. Две или четыре пары семенных пузырьков производят, хранят и высвобождают сперму через мужские поры. Яичники и яйцеводы в сегменте 13 выделяют яйцеклетки через женские поры в сегменте 14, а сперма выбрасывается из сегмента 15. одна или несколько пар сперматек В сегментах 9 и 10 (в зависимости от вида) присутствуют , которые представляют собой внутренние мешочки, которые принимают и хранят сперма другого червя во время совокупления. В результате 15-й сегмент одного червя выделяет сперму в сегменты 9 и 10 вместе с пузырьками-хранилищами своего партнера. Некоторые виды используют внешние сперматофоры для переноса спермы.
У Hormogaster samnitica и Hormogaster elisae транскриптомов были секвенированы библиотеки ДНК были обнаружены два половых феромона , и во всех образцах тканей обоих видов , аттрактин и темптин . [ 38 ] Половые феромоны, вероятно, важны для дождевых червей, поскольку они живут в среде, где химическая передача сигналов может играть решающую роль в привлечении партнера и облегчении ауткроссинга. Ауткроссинг позволит замаскировать проявление вредных рецессивных мутаций в потомстве. [ 39 ] (см . Дополнение ).
Копуляция и размножение — отдельные процессы у дождевых червей. Спаривающаяся пара перекрывает передние концы вентрально, и каждый обменивается спермой с другим. Клитор становится от красноватого до розоватого цвета. Через некоторое время после совокупления, спустя долгое время после разделения червей, клителлум (позади сперматек) выделяет материал, который образует кольцо вокруг червя. Затем червь выходит из кольца и при этом вводит в него свои собственные яйца и сперму другого червя. Таким образом, каждый червь становится генетическим отцом некоторых своих потомков (благодаря передаче своей собственной спермы другому дождевому червю) и генетической матерью (потомками из собственных яйцеклеток) остальных. Когда червь выскальзывает из кольца, концы кокона слипаются, образуя инкубатор ( кокон ) неопределенной луковичной формы, в котором развиваются эмбриональные черви. Следовательно, оплодотворение является внешним. Затем кокон помещают в почву. Через три недели вылупляется от 2 до 20 потомков, в среднем 4. Развитие прямое, т. е. без образования личинки.
восстановление ДНК
Воздействие дождевого червя Eisenia fetida на ионизирующего излучения вызвало разрывы цепей ДНК и окисление оснований ДНК . [ 40 ] Эти повреждения ДНК затем можно будет восстановить в соматических и сперматогенных клетках. [ 40 ] Клетки семенников дождевых червей также способны восстанавливать перекисью водорода . окислительные аддукты ДНК, индуцированные [ 41 ]
Передвижение

Дождевые черви перемещаются под землей посредством волн мышечных сокращений, которые поочередно укорачивают и удлиняют тело ( перистальтика ). Укороченная часть прикреплена к окружающей почве крошечными когтеобразными щетинками ( щетинками ), расположенными вдоль ее сегментированной длины. Во всех сегментах тела, кроме первого, последнего и клитора, в эпидермальной ямке каждого сегмента имеется кольцо S-образных щетинок (перихетин). Весь процесс рытья облегчается выделением смазывающей слизи. В результате перемещения по смазанным туннелям черви могут издавать под землей булькающие звуки, если их потревожить. Дождевые черви перемещаются по почве, с силой расширяя щели; Когда силы измеряются в зависимости от веса тела, только что вылупившиеся детеныши могут толкать вес, в 500 раз превышающий их собственный вес, тогда как крупные взрослые особи могут толкать только вес, в 10 раз превышающий их собственный вес. [ 42 ]
Регенерация
Дождевые черви обладают способностью регенерировать утраченные сегменты, но эта способность варьируется у разных видов и зависит от степени повреждения. Стефенсон (1930) посвятил этой теме главу своей монографии, а Дж. Гейтс в течение 20 лет изучал регенерацию у различных видов. Но «поскольку к этому вопросу было проявлено мало интереса», Гейтс (1972) опубликовал лишь некоторые из своих результатов. Тем не менее, они показывают, что у некоторых видов теоретически возможно вырастить двух целых червей из разрезанного пополам экземпляра.
Отчеты Гейтса включали:
- Eisenia fetida ( Savigny , 1826) с регенерацией головки в переднем направлении, возможной на каждом межсегментарном уровне вплоть до 23/24 включительно, тогда как хвосты регенерировали на любых уровнях после 20/21; таким образом, из одного могут вырасти два червя. [ 43 ]
- Lumbricus terrestris ( Linnaeus , 1758 ) заменял передние сегменты еще 13/14 и 16/17, но регенерации хвоста так и не было обнаружено.
- Perionyx excavatus (Perrier, 1872) легко регенерировал утраченные части тела в переднем направлении, начиная с 17/18, и в заднем направлении, начиная с 20/21.
- Lampito mauritii (Kinberg, 1867) с регенерацией в переднем направлении на всех уровнях до 25/26 и регенерацией хвоста от 30/31; Иногда считалось, что регенерация головы вызвана внутренней ампутацией, вызванной Sarcophaga sp. личиночная инвазия.
- Criodrilus lacuum (Hoffmeister, 1845) также обладает потрясающей регенеративной способностью: регенерация «головы» начинается уже с 40/41. [ 44 ]
Сообщалось о неопознанном тасманском дождевом черве, у которого выросла новая голова. [ 45 ]
Таксономия и распространение
В мире таксономии стабильная «классическая система» Микельсена (1900) и Стивенсона (1930) постепенно разрушалась из-за споров о том, как классифицировать дождевых червей, например, Фендер и Макки-Фендер (1990) зашли так далеко, что заявили: на уровне семейства , «Классификация дождевых червей мегаколецид находится в хаосе». [ 46 ] За прошедшие годы многие учёные разработали свои собственные системы классификации дождевых червей, что привело к путанице, и эти системы пересматривались и продолжают пересматриваться и обновляться. Используемая здесь система классификации, разработанная Блейкмором (2000), представляет собой современный возврат к классической системе, исторически проверенной и широко распространенной. [ 47 ]
Отнесение дождевого червя мегадрила к одному из таксономических семейств подотрядов Lumbricina и Moniligastrida основано на таких признаках, как строение клитора, расположение и расположение половых признаков (пор, предстательных желез и др.), количества желудков, и форму тела. [ 47 ] В настоящее время в базе данных названий видов названо более 6000 видов наземных дождевых червей. [ 48 ] но количество синонимов неизвестно.
Семейства с известным их распространением или происхождением: [ 47 ]
- Акантодрилиды
- Ailoscolecidae – Пиренеи и юго-восток США.
- Almidae – тропический экваториальный (Южная Америка, Африка, Индо-Азия).
- Benhamiinae - эфиопский, неотропический (возможное подсемейство Octochaetidae).
- Criodrilidae – юго-западная Палеарктика: Европа, Ближний Восток, Россия и Сибирь до побережья Тихого океана; Япония (Бивадрилус); в основном водный
- Diplocardiinae /-idae – Гондванский или Лавразийский? (подсемейство Acanthodrilidae)
- Enchytraeidae - космополиты, но редко встречаются в тропиках (обычно относят к Microdriles).
- Eudrilidae - Тропическая Африка к югу от Сахары.
- Exxidae - Неотропики: Центральная Америка и Карибский бассейн.
- Glossoscolecidae - Неотропические районы: Центральная и Южная Америка, Карибский бассейн.
- Haplotaxidae - космополитическое распространение (обычно отнесенное к Microdriles).
- Hormogastridae - Средиземноморье
- Kynotidae - малагасийский: Мадагаскар.
- Lumbricidae - Голарктика: Северная Америка, Европа, Ближний Восток, Центральная Азия и Япония.
- Lutodrilidae - Луизиана, юго-восток США.
- Мегасколециды
- Microchaetidae - наземные животные в Африке, особенно на лугах Южной Африки.
- Moniligastridae - Восточно-индийский субрегион.
- Ocnerodrilidae – Неотропы, Африка; Индия
- Octochaetidae - австралазийские, индийские, восточные, эфиопские, неотропические.
- Octochaetinae - австралазийские, индийские, восточные (подсемейство, если принято Benhamiinae)
- Sparganophilidae - Неарктика, Неотропика: Северная и Центральная Америка.
- Tumakidae - Колумбия, Южная Америка.
Как инвазивный вид
Из общего числа около 7000 видов только около 150 видов широко распространены по всему миру. Это сапсан или космополитический дождевой червь. [ 49 ] Из 182 таксонов дождевых червей, обнаруженных в США и Канаде, 60 (33%) являются интродуцированными видами.
Экология

Дождевые черви подразделяются на три основные экофизиологические категории: (1) черви, обитающие в листовой подстилке или компосте, которые не роют норы, живут на границе раздела почва-подстилка и питаются разлагающимися органическими веществами ( эпигейными ), например, Eisenia fetida ; (2) черви, обитающие в верхнем или подпочвенном слое почвы, которые питаются (почвой), роют норы и закапываются в почву, создавая горизонтальные норы в верхних 10–30 см почвы ( эндогейные ); и (3) черви, которые строят постоянные глубокие вертикальные норы, которые они используют для посещения поверхности в поисках растительного материала для еды, такого как листья ( anecic , что означает «достигать вверх»), например, Lumbricus terrestris . [ 50 ]
Популяции дождевых червей зависят как от физических, так и от химических свойств почвы, таких как температура, влажность, pH, соли, аэрация и текстура, а также от доступной пищи и способности вида к размножению и расселению. Одним из наиболее важных факторов окружающей среды является pH , но дождевые черви различаются в своих предпочтениях. Большинство предпочитают нейтральные или слегка кислые почвы. Lumbricus terrestris все еще присутствует при pH 5,4, Dendrobaena Octaedra при pH 4,3, а некоторые Megascolecidae присутствуют в чрезвычайно кислых гумусовых почвах. pH почвы также может влиять на количество червей, впадающих в диапаузу . Чем кислее почва, тем раньше черви переходят в диапаузу, и дольше всего остаются в диапаузе при рН 6,4.
На дождевых червей охотятся многие виды птиц (например, малиновки, скворцы , дрозды , чайки , вороны ), змеи, лесные черепахи, млекопитающие (например , медведи , кабаны, лисы , ежи , свиньи , кроты) . [ 51 ] ) и беспозвоночных (например , муравьев , [ 52 ] плоские черви , жужелицы и другие жуки , улитки , пауки и слизни ). У дождевых червей имеется множество внутренних паразитов , включая простейших , платихельминтов, клещей и нематод ; червей их можно обнаружить в крови , семенных пузырьках , целоме или кишечнике , а также в их коконах (например, клещ Histiostoma murchiei является паразитом коконов дождевых червей). [ 53 ] ).
Деятельность дождевых червей аэрирует и перемешивает почву, способствует минерализации питательных веществ и их поглощению растительностью. Некоторые виды дождевых червей выходят на поверхность и питаются присутствующими там более высокими концентрациями органических веществ, смешивая их с минеральной почвой. Поскольку высокий уровень смешивания органических веществ связан с плодородием почвы , фермеры и садоводы обычно считают обилие дождевых червей полезным. [ 54 ] [ 55 ] Еще в 1881 году Чарльз Дарвин писал: «Можно сомневаться, есть ли много других животных, сыгравших столь важную роль в мировой истории, как эти низкоорганизованные существа». [ 56 ]

Кроме того, хотя, как следует из названия, основная среда обитания дождевых червей находится в почве, они не ограничиваются этой средой обитания. Червь-клейка Eisenia fetida живет в разлагающихся растительных веществах и навозе. Arctiostrotus vancouverensis с острова Ванкувер и Олимпийского полуострова обычно встречается в гниющих бревнах хвойных пород. Aporrectodea limicola , Sparganophilus spp. и некоторые другие встречаются в иле ручьев. Некоторые виды ведут древесный образ жизни. [ 57 ] некоторые водные, некоторые эвригалинные (толерантные к соленой воде) и прибрежные (живущие на берегу моря, например, Pontodriluslitoralis ). [ 58 ] Даже среди почвенных видов особые места обитания, такие как почвы, полученные из серпентина , имеют собственную фауну дождевых червей.
Вермикомпостирование органических «отходов» и добавление этого органического вещества в почву, предпочтительно в виде поверхностной мульчи , обеспечит несколько видов дождевых червей необходимыми пищевыми и питательными веществами, а также создаст оптимальные условия температуры и влажности, которые будут стимулировать их активность. .
Дождевые черви являются экологическими индикаторами здоровья почвы . Дождевые черви питаются разлагающимися веществами в почве, и анализ содержимого их пищеварительного тракта дает представление об общем состоянии почвы. Кишечник дождевого червя накапливает химические вещества, в том числе тяжелые металлы, такие как кадмий , ртуть , цинк и медь . Размер популяции дождевых червей указывает на качество почвы, поскольку в здоровой почве обитает большее количество дождевых червей. [ 59 ]
Воздействие на окружающую среду
Основные преимущества деятельности дождевых червей для плодородия почвы в сельском хозяйстве можно резюмировать следующим образом:
- Биологический : во многих почвах дождевые черви играют важную роль в преобразовании крупных кусков органического вещества в богатый гумус , тем самым улучшая плодородие почвы. Это достигается за счет того, что червь вытягивает под поверхность отложившиеся органические вещества, такие как листопад или навоз, либо в пищу, либо для закупорки норы. Попав в нору, червь измельчит лист, частично переварит его и смешает с землей. Червевые слепки (см. внизу справа) могут содержать на 40 процентов больше гумуса, чем верхние 9 дюймов (230 мм) почвы, в которой живет червь. [ 60 ]

- Химические вещества : помимо мертвого органического вещества , дождевой червь также заглатывает любые другие достаточно мелкие частицы почвы, включая песчинки размером до 1 ⁄ 20 дюйма (1,3 мм) — в желудок, где эти мельчайшие фрагменты песка измельчают все в мелкую пасту, которая затем переваривается в кишечнике. Когда червь выделяет их в виде слепков, откладывающихся на поверхности или глубже в почве, минералы и питательные вещества для растений переходят в доступную для использования растениями форму. Исследования, проведенные в Соединенных Штатах, показывают, что свежие отбросы дождевых червей в пять раз богаче доступным азотом , в семь раз богаче доступными фосфатами и в 11 раз богаче доступным калием , чем окружающие верхние 6 дюймов (150 мм) почвы. В условиях обилия гумуса вес полученных слепков может превышать 4,5 кг (9,9 фунта) на червя в год. [ 60 ]
- Физический : роющие дождевые черви создают множество каналов в почве и имеют большое значение для поддержания структуры почвы , обеспечивая процессы аэрации и дренажа. [ 61 ] пермакультуры Соучредитель Билл Моллисон отмечает, что, скользя в своих туннелях, дождевые черви «действуют как бесчисленная армия поршней, вкачивающих воздух в почву и из нее в 24-часовом цикле (ночью быстрее)». [ 62 ] Таким образом, дождевой червь не только создает проходы для воздуха и воды, которые проходят через почву, но также модифицирует жизненно важный органический компонент, который делает почву здоровой (см. Биотурбация ). Дождевые черви способствуют образованию богатых питательными веществами отливок (шариков почвы, устойчивых в почвенной слизи), обладающих высокой почвенной агрегацией, плодородием и качеством почвы. [ 60 ] В подзолистых почвах дождевые черви могут стирать характерный полосатый вид почвенного профиля, смешивая органический (LFH), элювиальный (E) и верхний иллювиальный (B) горизонты, создавая единый темный горизонт Ap. [ 63 ] [ 64 ]
Дождевые черви ускоряют круговорот питательных веществ в системе почва-растение за счет фрагментации и смешивания растительных остатков – физического измельчения и химического переваривания. [ 60 ] Существование дождевого червя нельзя воспринимать как нечто само собой разумеющееся. Доктор У. Э. Шевелл-Купер наблюдал «огромные количественные различия между соседними садами», а на популяции червей влияет множество факторов окружающей среды, на многие из которых могут влиять хорошие методы управления со стороны садовника или фермера. [ 65 ]
Дарвин подсчитал, что пахотная земля содержит до 53 000 червей на акр (130 000/га), но более поздние исследования дали цифры, предполагающие, что даже бедная почва может содержать 250 000 червей на акр (620 000/га), в то время как на богатых плодородных сельскохозяйственных угодьях может быть до 1 750 000 на акр (4 300 000 на га), а это означает, что вес дождевых червей под землей фермера может быть больше, чем вес домашнего скота на ее поверхности. Популяции дождевых червей в богатом органическими веществами верхнем слое почвы намного выше – в среднем 500 на квадратный метр (46/кв. фут) и до 400 г2. [ сомнительно – обсудить ] – так что из 7 миллиардов из нас на каждого живущего сегодня человека приходится 7 миллионов дождевых червей. [ 66 ]
Способность расщеплять органические материалы и выделять концентрированные питательные вещества делает дождевых червей функциональным участником проектов восстановления. В ответ на нарушения экосистемы некоторые участки использовали дождевых червей для подготовки почвы к возвращению местной флоры. Исследования Тропической экологической станции Ламто утверждают, что дождевые черви положительно влияют на скорость образования макроагрегатов, что является важной характеристикой структуры почвы. [ 67 ] Также было обнаружено, что устойчивость агрегатов к воздействию воды улучшается, когда они создаются дождевыми червями. [ 67 ]
, хотя и не полностью подсчитаны, Выбросы дождевых червей парниковых газов вероятно, способствуют глобальному потеплению, особенно с учетом того, что обитающие на верхушках дождевые черви увеличивают скорость углеродных циклов и были распространены людьми во многих новых географических регионах. [ 68 ]
Угрозы
Азотные удобрения имеют тенденцию создавать кислую среду , губительную для червей, а мертвые особи часто обнаруживаются на поверхности после применения таких веществ, как ДДТ , известковая сера и арсенат свинца . В Австралии изменения в методах ведения сельского хозяйства, такие как применение суперфосфатов на пастбищах и переход от пастбищного земледелия к пахотному земледелию, оказали разрушительное воздействие на популяции гигантского дождевого червя Гиппсленда , что привело к их классификации как охраняемого вида . Во всем мире некоторые популяции дождевых червей были уничтожены из-за отклонения от органического производства и распыления синтетических удобрений и биоцидов, при этом по крайней мере три вида в настоящее время числятся вымершими, но многие другие находятся под угрозой исчезновения. [ 69 ]
Экономический эффект

Различные виды червей используются в вермикультуре — практике скармливания дождевым червям органических отходов для разложения пищевых отходов. Обычно это Eisenia fetida (или ее близкий родственник Eisenia andrei ) или клейменный червь, широко известный как тигровый червь или красный вигглер. Они отличаются от почвенных дождевых червей. В тропиках африканский ночной змей Eudrilus eugeniae. [ 70 ] и индийский синий Perionyx excavatus .
Дождевые черви продаются по всему миру; рынок огромен. По словам Дуга Колликатта, «в 1980 году из Канады было экспортировано 370 миллионов червей, при этом стоимость канадского экспорта составила 13 миллионов долларов, а розничная стоимость в Америке - 54 миллиона долларов». [ 71 ]
Дождевые черви являются отличным источником белка для рыбы, птицы и свиней, но также традиционно используются в пищу человеком. Ноке — кулинарный термин, используемый маори Новой Зеландии для обозначения дождевых червей, которых они считают деликатесом для своих вождей.
См. также
- Дрилосфера , часть почвы, находящаяся под воздействием выделений и отбросов дождевых червей.
- «Формирование растительной плесени под действием червей» , книга Чарльза Дарвина 1881 года.
- Почвенная жизнь
- Вермикомпост
- Вермифильтр
- Вермифильтр для туалета
- Червь очаровательный
Ссылки
- ^ Андерсон, Фрэнк; Джеймс, Сэмюэл (июнь 2017 г.). «Эволюция дождевых червей» . БМК . Проверено 3 февраля 2024 г.
- ^ Омодео, Пьетро (2000). «Эволюция и биогеография мегадрилей (Annelida, Clitellata)» . Итальянский журнал зоологии . 67–2 (2): 179–201. дои : 10.1080/11250000009356313 . S2CID 86293273 .
- ^ Бонковски, Майкл; Гриффитс, Брайан С.; Ритц, Карл (ноябрь 2000 г.). «Пищевые предпочтения дождевых червей почвенным грибам». Педобиология . 44–6 (6):667. Бибкод : 2000Педоб..44..666Б . дои : 10.1078/S0031-4056(04)70080-3 .
- ^ Лофти, Клайв А.; Лофти, младший (1977). Биология дождевых червей . Лондон: Чепмен и Холл. п. 80. ИСБН 0-412-14950-8 .
- ^ Эдвардс, Клайв А.; Лофти, младший (1977). Биология дождевых червей . Лондон: Чепмен и Холл. п. 19. ISBN 0-412-14950-8 .
- ^ Эдвардс, Клайв А.; Лофти, младший (1977). Биология дождевых червей . Лондон: Чепмен и Холл. стр. предисловие. ISBN 0-412-14950-8 .
- ^ Коулман, Дэвид С.; Кроссли, округ Колумбия; Хендрикс, Пол Ф. (2004). Основы экологии почв . Амстердам; Бостон: Elsevier Academic Press. п. 170. ИСБН 0-12-179726-0 .
- ^ Перейти обратно: а б Кливленд П. Хикман-младший; Ларри С. Робертс; Фрэнсис М. Хикман (1984). Комплексные принципы зоологии (7-е изд.). Times Mirror/Издательство колледжа Мосби. п. 344 . ISBN 978-0-8016-2173-4 .
- ^ Блейкмор 2012 , с. хл.
- ^ Блейкмор, Р.Дж.; и др. (2007). «Megascolex (Promegascolex) mekongianus Cognetti, 1922 - его размеры, экология и отнесение к Amynthas (Clitellata/Oligochaeta: Megascolecidae)» (PDF) . Зоологические брошюры.
- ^ Эдвардс и Болен 1996 , с. 11.
- ^ Перейти обратно: а б с Симс и Джерард 1985 , стр. 3–6.
- ^ Эдвардс и Болен 1996 , с. 3.
- ^ Фельдкамп, Дж. (1924). «Фельдкамп, Дж. «Исследования половых признаков и спаривания дождевых червей. Зоологические ежегодники». Анатомия . 46 : 609–632.
- ^ Эдвардс и Болен 1996 , с. 8-9.
- ^ Эдвардс и Болен 1996 , с. 1.
- ^ Симс и Джерард 1985 , с. 8.
- ^ Перейти обратно: а б Эдвардс и Болен 1996 , с. 12.
- ^ Эдвардс и Болен 1996 , с. 6.
- ^ Мирмекофурмис (2018). «Знакомьтесь: брызгающий дождевой червь» . Ютуб. Архивировано из оригинала ( видео ) 30 октября 2021 г.
- ^ «Эксперимент: сравнение скоростей нервных волокон двух размеров» . Задний двор Мозги . Проверено 4 апреля 2015 г.
- ^ Древес, CD; Ланда, КБ; Макфолл, Дж.Л. (1978). «Активность гигантских нервных волокон у неповрежденных, свободно перемещающихся дождевых червей» . Журнал экспериментальной биологии . 72 : 217–227. дои : 10.1242/jeb.72.1.217 . ПМИД 624897 .
- ^ Куиллин, К.Дж. (1999). «Кинематическое масштабирование передвижения гидростатических животных: онтогенез перистальтического ползания дождевого червя lumbricus terrestris». Журнал экспериментальной биологии . 202 (6): 661–674. дои : 10.1242/jeb.202.6.661 . ПМИД 10021320 .
- ^ Перейти обратно: а б «Дождевой червь-нервная система» . Кронодон . Проверено 3 апреля 2015 г.
- ^ Элвуд, RW (2011). «Боль и страдания у беспозвоночных?» . Журнал ИЛАР . 52 (2): 175–84. дои : 10.1093/ilar.52.2.175 . ПМИД 21709310 .
- ^ Смит, Дж. А. (1991). «Вопрос о боли у беспозвоночных» . Журнал ИЛАР . 33 (1–2): 25–31. дои : 10.1093/ilar.33.1-2.25 .
- ^ Релих, П.; Арос, Б.; Вираг, Сз. (1970). «Тонкая структура фоторецепторных клеток дождевого червя Lumbricus Terrestris ». Журнал клеточных исследований и микроскопической анатомии . 104 (3): 345–357. дои : 10.1007/BF00335687 . ПМИД 4193489 . S2CID 22771585 .
- ^ Эдвардс и Болен 1996 , с. 13.
- ^ Либеке, Мануэль; Стриттматтер, Николь; Фирн, Сара; Морган, А. Джон; Килле, Питер; Фухсер, Йенс; Уоллис, Дэвид; Пальчиков, Виталий; Робертсон, Джереми (4 августа 2015 г.). «Уникальные метаболиты защищают дождевых червей от растительных полифенолов» . Природные коммуникации . 6 : 7869. Бибкод : 2015NatCo...6.7869L . дои : 10.1038/ncomms8869 . ПМЦ 4532835 . ПМИД 26241769 .
- ^ Эдвардс и Болен 1996 , стр. 13–15.
- ^ Симс и Джерард 1985 , с. 10.
- ^ Кливленд П. Хикман младший; Ларри С. Робертс; Фрэнсис М. Хикман (1984). Комплексные принципы зоологии (7-е изд.). Times Mirror/Издательство колледжа Мосби. стр. 344–345 . ISBN 978-0-8016-2173-4 .
- ^ Перейти обратно: а б Фараби, Г.Дж. «Выделительная система» . Архивировано из оригинала 30 июля 2012 года . Проверено 29 июля 2012 г.
- ^ Кливленд П. Хикман младший; Ларри С. Робертс; Фрэнсис М. Хикман (1984). Комплексные принципы зоологии (7-е изд.). Times Mirror/Издательство колледжа Мосби. стр. 345–346 . ISBN 978-0-8016-2173-4 .
- ^ Домингес Х., Айра М., Брейнхольт Дж.В., Стоянович М., Джеймс С.В., Перес-Лосада М. (2015). «Подземная эволюция: новые корни старого дерева люмбрицидных дождевых червей» . Мол. Филогенет. Эвол . 83 : 7–19. Бибкод : 2015МОЛПЭ..83....7Д . дои : 10.1016/j.ympev.2014.10.024 . ПМК 4766815 . ПМИД 25463017 .
- ^ Фернандес Р., Альмодовар А., Ново М., Симанкас Б., Диас Косин DJ (2012). «Усложнение комплекса: новое понимание филогении, разнообразия и происхождения партеногенеза в комплексе видов Aporrectodea caliginosa (Oligochaeta, Lumbricidae)». Мол. Филогенет. Эвол . 64 (2): 368–79. Бибкод : 2012МОЛПЭ..64..368Ф . дои : 10.1016/j.ympev.2012.04.011 . ПМИД 22542691 .
- ^ Косин DJD, Ново М., Фернандес Р. (2011) Размножение дождевых червей: половой отбор и партеногенез. В: Караджа А. (ред.) Биология дождевых червей. Биология почвы, том 24. Springer, Berlin, Heidelberg, стр. 76 и далее. [1]
- ^ Ново М, Рисго А, Фернандес-Герра А, Гирибет Г (2013). «Эволюция феромонов, репродуктивные гены и сравнительная транскриптомика средиземноморских дождевых червей (аннелиды, олигохеты, hormogastridae)» . Мол. Биол. Эвол . 30 (7): 1614–29. дои : 10.1093/molbev/mst074 . hdl : 10261/94159 . ПМИД 23596327 .
- ^ Бернштейн Х., Хопф Ф.А., Мишод Р.Э. (1987). «Молекулярная основа эволюции пола». Молекулярная генетика развития . Достижения генетики. Том. 24. С. 323–70. дои : 10.1016/S0065-2660(08)60012-7 . ISBN 978-0-12-017624-3 . ПМИД 3324702 .
- ^ Перейти обратно: а б Хертель-Аас Т., Оутон Д.Х., Яворска А., Брунборг Г. (ноябрь 2011 г.). «Индукция и восстановление разрывов цепей ДНК и окисленных оснований в соматических и сперматогенных клетках дождевого червя Eisenia fetida после воздействия ионизирующего излучения». Мутагенез . 26 (6): 783–93. дои : 10.1093/mutage/ger048 . ПМИД 21825113 .
- ^ Чанг В.С., Цай CW, Линь CC, Линь CH, Шен WC, Линь СС, Бау Д.Т. (2011). «Дождевые черви восстанавливают индуцированные H2O2 окислительные аддукты ДНК, не удаляя димеры пиримидина, индуцированные УФ-излучением». В Виво . 25 (6): 977–81. ПМИД 22021692 .
- ^ Куиллан, Кей Джей (2000). «Онтогенетическое масштабирование роющих сил дождевого червя Lumbricus terrestris» . Журнал экспериментальной биологии . 203 (Часть 18): 2757–2770. дои : 10.1242/jeb.203.18.2757 . ПМИД 10952876 . Проверено 4 апреля 2015 г.
- ^ Гейтс, GE (апрель 1949 г.). «Регенерация дождевого червя Eisenia Foetida (Савиньи) 1826. I. Передняя регенерация» (PDF) . Биологический вестник . 96 (2): 129–139. дои : 10.2307/1538195 . JSTOR 1538195 . ПМИД 18120625 . Архивировано из оригинала (PDF) 1 апреля 2007 г.
- ^ Гейтс, GE (1 января 1953 г.). «О регенеративной способности дождевых червей семейства Lumbricidae». Американский натуралист из Мидленда . 50 (2): 414–419. дои : 10.2307/2422100 . JSTOR 2422100 .
- ^ «Предметы Беспозвоночные 20а» . Архивировано из оригинала 22 июня 2006 г. Проверено 20 ноября 2006 г.
- ^ Фендер и Макки-Фендер (1990). Руководство по биологии почвы . Уайли-Интерсайенс. ISBN 978-0-471-04551-9 .
- ^ Перейти обратно: а б с Блейкмор, Р.Дж. (2006) (март 2006 г.). «Пересмотренный справочник по всемирным семействам дождевых червей от Блейкмора (2000) плюс обзоры Criodrilidae (включая Biwadrilidae) и Octochaetidae» (PDF) . Серия текстов с возможностью поиска по биоразнообразию, экологии и систематике дождевых червей из различных регионов мира . annelida.net . Проверено 15 мая 2012 г.
{{cite web}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ «Дождевые черви» . Earthworms.elte.hu . Проверено 19 марта 2022 г.
- ^ [ссылка на Блейкмора (2016) Cosmopolitan Earthworms]
- ^ Дождевые черви: обновлятели агроэкосистем (SA Fall, 1990 (v3n1)) Архивировано 13 июля 2007 г. в Wayback Machine.
- ^ Гулд, Эдвин; Макши, Уильям; Гранд, Теодор (1993). «Функция звезды у звездоноса, Condylura cristata». Журнал маммологии . 74 (1): 108–116. дои : 10.2307/1381909 . ISSN 0022-2372 . JSTOR 1381909 .
- ^ Дежан, А.; Шаткс, Б. (1999). «Поведение Psalidomyrmex procerus (Formicidae; Ponerinae), специализированного хищника дождевых червей (Annelida)» при захвате добычи. Социобиология : 545–554. ISSN 0361-6525 .
- ^ Оливер, Джеймс Х. (1962). «Клещ-паразит в коконах дождевых червей» . Журнал паразитологии . 48 (1): 120–123. дои : 10.2307/3275424 . ISSN 0022-3395 . JSTOR 3275424 . ПМИД 14481811 .
- ^ Департамент первичной промышленности Нового Южного Уэльса, Как дождевые черви могут помочь вашей почве. Архивировано 7 августа 2017 г. в Wayback Machine.
- ^ Ассоциация главных садовников округа Галвестон, Благотворители в саду: # 38 Дождевые черви
- ^ Дарвин, Чарльз (1881). Образование растительной плесени под действием червей и наблюдения за их повадками . Джон Мюррей . Найдено в проекте Гутенберга Etext «Формирование растительной формы», Дарвин.
- ^ Гаум, Лоуренс; Шеной, Мега; Захариас, Мерри; Борхес, Рене М. (май 2006 г.). «Сосуществование муравьев и древесного дождевого червя в мирмекофите Индийских Западных Гат: противохищнический эффект слизи дождевого червя» . Журнал тропической экологии . 22 (3): 341–344. дои : 10.1017/S0266467405003111 . ISSN 1469-7831 .
- ^ Блейкмор, Р.Дж. (2007). «Происхождение и пути распространения космополита Pontodrilus Litoralis (Oligochaeta: Megascolecidae)» . Европейский журнал почвенной биологии.
{{cite web}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Фрюнд, Хайнц-Кристиан; Грефе, Ульферт; Тишер, Сабина (2011), Карача, Айтен (редактор), «Дождевые черви как биоиндикаторы качества почвы» , Биология дождевых червей , Биология почвы, том. 24, Берлин, Гейдельберг: Springer Berlin Heidelberg, стр. 261–278, doi : 10.1007/978-3-642-14636-7_16 , ISBN 978-3-642-14635-0 , получено 18 февраля 2021 г.
- ^ Перейти обратно: а б с д Найл С. Брэди; Рэй Р. Вейл (2009). Элементы природы и свойства почв (3-е изд.). Прентис Холл. ISBN 978-0-13-501433-2 .
- ^ Лайнс-Келли, Ребекка (2021). «Как дождевые черви могут помочь вашей почве» . www.dpi.nsw.gov.au. Архивировано из оригинала 08 октября 2021 г. Проверено 30 ноября 2021 г.
- ^ Моллисон, Билл, Пермакультура - Руководство дизайнера , Tagari Press, 1988.
- ^ РТ Фишер (январь 1928 г.). «Изменения почвы и лесоводство в Гарвардском лесу» (PDF) . Экология . 9 (1): 6–11. Бибкод : 1928Ecol....9....6F . дои : 10.2307/1929537 . JSTOR 1929537 . Проверено 19 марта 2022 г.
- ^ Лангмейд, К.К. (1 февраля 1964 г.). «Некоторые последствия нашествия дождевых червей на целинные подзолы» . Канадский журнал почвоведения . 44 (1): 34–37. дои : 10.4141/cjss64-005 . Проверено 19 марта 2022 г.
- ^ Купер, Шевелл; Почва, гумус и здоровье ISBN 978-0-583-12796-7
- ^ Блейкмор, Р.Дж. (2017) (12 февраля 2017 г.). «Статья о природе в честь дня рождения Чарльза Дарвина 12 февраля» . ВермЭкология.
{{cite web}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Перейти обратно: а б Бланшар, Эрик (1 декабря 1992 г.). «Восстановление дождевыми червями (megascolecidae) макроагрегатной структуры деструктурированной почвы саванны в полевых условиях». Биология и биохимия почвы . 24 (12): 1587–1594. Бибкод : 1992SBiBi..24.1587B . дои : 10.1016/0038-0717(92)90155-Q .
- ^ Берк, Дэвид (26 декабря 2019 г.). «Сила фекалий дождевых червей и как они могут повлиять на изменение климата» . ЦБК .
- ^ Блейкмор, Р.Дж. (2018) (2018). «Критическое сокращение численности дождевых червей органического происхождения в условиях интенсивного сельского хозяйства, истощающего гуминовые ПОВ» . Почвенные системы . 2 (2). Soil Systems 2(2): 33: 33. doi : 10.3390/soilsystems2020033 .
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Блейкмор, Р.Дж. (2015). «Эко-таксономический профиль культового биогумуса - африканского ночного змея, Eudrilus eugeniae (Кинберг, 1867)» . Африканские беспозвоночные 56: 527-548. Архивировано из оригинала 22 октября 2016 г.
{{cite web}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Колликатт, Дуг. «Биология ночного ползуна (Lumbricus terrestris)» . ПриродаСевер . Проверено 5 июня 2022 г.
Цитируемые работы
- Блейкмор, Роберт Дж. (2012). Космополитические дождевые черви - эко-таксономический справочник по видам сапсанов мира. (5-е изд.) . Иокогама, Япония: VermEcology.
- Эдвардс, Клайв А.; Болен, П.Дж. (1996). Биология и экология дождевых червей . Springer Science & Business Media. ISBN 978-0-412-56160-3 .
- Симс, Реджинальд Уильям; Джерард, Б. (1985). Дождевые черви: определители и примечания для определения и изучения видов . Лондон: Опубликовано для Лондонского Линнеевского общества и Ассоциации наук об эстуариях и солоноватой воде Э. Дж. Бриллом / доктором. В. Бакхейс.
Дальнейшее чтение
- Эдвардс, Клайв А. (ред.) Экология дождевых червей . Бока-Ратон: CRC Press, 2004. Второе исправленное издание. ISBN 0-8493-1819-X
- Ли, Кенет Э. Дождевые черви: их экология и связь с почвами и землепользованием . Академическая пресса. Сидней, 1985 год. ISBN 0-12-440860-5
- Стюарт, Эми. Земля сдвинулась: о выдающихся достижениях дождевых червей . Чапел-Хилл, Северная Каролина: Algonquin Books, 2004. ISBN 1-56512-337-9
Внешние ссылки
 СМИ, связанные с дождевыми червями, на Викискладе?
СМИ, связанные с дождевыми червями, на Викискладе?  Данные, относящиеся к Лумбрицине , в Wikispecies
Данные, относящиеся к Лумбрицине , в Wikispecies - Чисхолм, Хью , изд. (1911). . Британская энциклопедия (11-е изд.). Издательство Кембриджского университета.
| Базы данных органов управления : Национальные |
|---|