Штрих-кодирование ДНК грибов
| Часть серии о |
| ДНК-штрихкодирование |
|---|
 |
| По таксонам |
| Другой |
Штрих-кодирование ДНК грибов — это процесс идентификации видов биологического царства грибов посредством амплификации и секвенирования конкретных последовательностей ДНК и их сравнения с последовательностями, хранящимися в базе данных штрих-кодов ДНК, такой как справочная база данных ISHAM, [ 1 ] или Система данных «Штрих-код жизни» (ЖИРНЫЙ). В этой попытке штрих-кодирование ДНК опирается на универсальные гены, которые в идеале присутствуют у всех грибов с одинаковой степенью вариации последовательностей. Межвидовая изменчивость, т. е. межвидовая изменчивость выбранного гена штрих-кода ДНК, должна превышать внутривидовую (внутривидовую) изменчивость. [ 2 ]
Фундаментальной проблемой систематики грибов является существование телеоморфных и анаморфных стадий в их жизненном цикле. Эти морфы обычно резко различаются по своему фенотипическому внешнему виду, что предотвращает прямую ассоциацию бесполого анаморфа с половым телеоморфом. Более того, виды грибов могут включать несколько штаммов, которые могут различаться по своей морфологии или таким признакам, как использование углерода и азота, что часто приводит к их описанию как разных видов, что в конечном итоге приводит к образованию длинных списков синонимов. [ 3 ] Штрих-кодирование ДНК грибов может помочь идентифицировать и связать анаморфные и телеоморфные стадии грибов и тем самым уменьшить запутанное множество названий грибов. По этой причине микологи были одними из первых, кто возглавил исследование дискриминации видов с помощью последовательностей ДНК. [ 3 ] [ 4 ] [ 5 ] [ 6 ] [ 7 ] [ 8 ] по крайней мере на 10 лет раньше, чем предложение Пола Д. Н. Хеберта и его коллег о штрих-кодировании ДНК для животных в 2003 году, которые популяризировали термин «штрих-кодирование ДНК». [ 9 ] [ 10 ]
Успех идентификации грибов с помощью последовательностей штрих-кодов ДНК зависит от количественного (полнота) и качественного (уровень идентификации) аспекта справочной базы данных. Без базы данных, охватывающей широкий таксономический диапазон грибов, многие идентификационные запросы не приведут к достаточно близкому совпадению. Аналогичным образом, без существенных кураторских усилий по поддержанию записей на высоком таксономическом уровне идентификации, запросы – даже если они могут иметь близкое или точное совпадение в справочной базе данных – не будут информативными, если наиболее близкое совпадение идентифицируется только с типом или уровень класса . [ 11 ] [ 12 ]
Еще одной важной предпосылкой для штрих-кодирования ДНК является способность однозначно проследить происхождение данных штрих-кода ДНК до исходного образца, так называемого ваучерного образца. Это обычная практика в биологии наряду с описанием новых таксонов становятся ваучерные экземпляры, на которых основано таксономическое описание , когда типовыми экземплярами . Если идентичность определенного таксона (или генетической последовательности в случае штрих-кодирования ДНК) вызывает сомнения, исходный образец можно повторно изучить для проверки и, в идеале, решения проблемы. Образцы ваучера должны быть четко маркированы как таковые, включая постоянный идентификатор ваучера, который однозначно связывает образец с данными штрих-кода ДНК, полученными из него. Кроме того, эти ваучерные образцы следует хранить в общедоступных хранилищах, таких как научные коллекции или гербарии, чтобы сохранить их для дальнейшего использования и облегчить исследования с использованием депонированных образцов. [ 13 ]
Маркеры ДНК штрих-кода
[ редактировать ]Внутренний транскрибируемый спейсер (ITS) – основной штрих-код грибов.
[ редактировать ]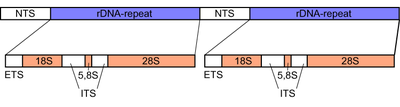
У грибов внутренний транскрибируемый спейсер ( ITS ) представляет собой участок длиной примерно 600 пар оснований в рибосомном тандемном повторяющемся кластере генов генома ядерного . Область фланкирована последовательностями ДНК малой субъединицы рибосомы (SSU) или 18S субъединицы на 5'-конце и большой субъединицы (LSU) или 28S субъединицы на 3'-конце. [ 14 ] [ 15 ] Сам внутренний транскрибируемый спейсер состоит из двух частей, ITS1 и ITS2 , которые отделены друг от друга субъединицей 5.8S , вложенной между ними. Подобно фланкирующим субъединицам 18S и 28S, субъединица 5.8S содержит высококонсервативную последовательность ДНК, поскольку они кодируют структурные части рибосомы , которая является ключевым компонентом во внутриклеточном синтезе белка .
Благодаря нескольким преимуществам ITS (см. ниже) и большому объему данных о последовательностях, накопленных в 1990-х и начале 2000-х годов, Begerow et al. (2010) и Шох и др. (2012) предложили область ITS в качестве первичной области штрих-кода ДНК для генетической идентификации грибов . [ 12 ] [ 2 ]
ОБЪЕДИНЯЙТЕСЬ [ 16 ] представляет собой открытую базу данных штрих-кодов ITS для грибов и всех других эукариот.
Праймеры
[ редактировать ]Консервативные фланкирующие области 18S и 28S служат опорными точками для праймеров, используемых для ПЦР- амплификации региона ITS . [ 17 ] Более того, консервативная вложенная область 5.8S позволяет создавать «внутренние» праймеры, т.е. праймеры, присоединяющиеся к комплементарным последовательностям внутри области ITS. Уайт и др. (1990) предложили такие внутренние праймеры, названные ITS2 и ITS3, а также фланкирующие праймеры ITS1 и ITS4 в субъединицах 18S и 28S соответственно. [ 17 ] Благодаря своей почти универсальной применимости для ITS-секвенирования у грибов, эти праймеры широко используются и сегодня. Оптимизированные праймеры специально для секвенирования ITS у Dikarya (включая Basidiomycota и Ascomycota ) были предложены Toju et al. (2012). [ 18 ]
Для большинства грибов праймеры ITS, предложенные White et al. (1990) стали стандартными праймерами, используемыми для ПЦР-амплификации. Эти праймеры: [ 17 ]
|
Форвардные праймеры:
|
Обратные праймеры:
|
Преимущества и недостатки
[ редактировать ]Основным преимуществом использования области ITS в качестве молекулярного маркера и штрих-кода ДНК грибов является то, что весь кластер рибосомальных генов расположен в тандемных повторах, т.е. в нескольких копиях. [ 15 ] Это позволяет проводить ПЦР-амплификацию и секвенирование по Сэнгеру даже из небольших образцов материала (при условии, что ДНК не фрагментирована из-за возраста или других дегенеративных воздействий ). [ 14 ] обычно наблюдается высокая вероятность успеха ПЦР Следовательно, при амплификации ITS . Однако этот показатель успеха сильно варьируется среди групп грибов: от 65% у не-Dikarya (включая ныне парафилетические Mucoromycotina , Chytridiomycota и Blastocladiomycota ) до 100% у Saccharomycotina и Basidiomycota. [ 2 ] (за исключением очень низкого успеха при Pucciniomycotina ). [ 19 ] Более того, выбор праймеров для ITS- амплификации может привести к смещению в сторону определенных таксономических групп грибов. [ 20 ] Например, «универсальные» ITS. праймеры [ 17 ] не способны амплифицировать около 10% протестированных образцов грибов. [ 19 ]
Тандемные повторы кластера рибосомальных генов вызывают проблему значительной гетерогенности внутригеномных последовательностей, наблюдаемую среди копий ITS нескольких групп грибов. [ 21 ] [ 22 ] [ 23 ] При секвенировании по Сэнгеру это приведет к тому, что считывания последовательностей ITS разной длины будут накладываться друг на друга, что потенциально сделает полученный хроматограф нечитаемым. Более того, из-за некодирующей природы региона ITS , которая может привести к значительному количеству инделей , невозможно последовательно выровнять последовательности ITS от сильно расходящихся видов для дальнейшего более масштабного филогенетического анализа. [ 9 ] [ 14 ] Степень гетерогенности внутригеномных последовательностей можно исследовать более подробно посредством молекулярного клонирования первоначально амплифицированных ПЦР последовательностей ITS с последующим секвенированием клонов. Эта процедура первоначальной ПЦР-амплификации с последующим клонированием ампликонов и , наконец, секвенированием клонированных продуктов ПЦР является наиболее распространенным подходом получения ITS последовательностей для метабаркодирования ДНК образцов окружающей среды, в которых одновременно может присутствовать множество различных видов грибов. Однако этот подход секвенирования после клонирования редко применялся для последовательностей ITS , которые составляют эталонные библиотеки, используемые для идентификации с помощью штрих-кода ДНК, что потенциально дает недооценку существующих вариаций последовательностей ITS во многих образцах. [ 24 ]
Средневзвешенное арифметическое внутривидовой (внутривидовой) изменчивости ИТС у грибов составляет 2,51%. Однако эта изменчивость может колебаться от 0%, например, у Serpula lacrymans (n=93 образца), свыше 0,19% у Tuber melanosporum (n=179) до 15,72% у Rhizoctonia solani (n=608) или даже 24,75% у Tuber melanosporum (n=179). Pisolithustinctorius (n=113). Таким образом , в случаях высокой внутривидовой изменчивости ITS применение порога изменчивости последовательностей в 3% – канонического верхнего значения для внутривидовой изменчивости – приведет к более высокой оценке операционных таксономических единиц (OTU), то есть предполагаемых видов, чем на самом деле. находятся в образце. [ 25 ] В случае видов грибов, значимых с медицинской точки зрения, более строгий порог изменчивости ITS в 2,5% позволяет точно идентифицировать до видового уровня только около 75% всех видов. [ 1 ]
С другой стороны, морфологически четко определенные, но эволюционно молодые комплексы видов или виды-двойники могут отличаться (если вообще отличаться) в нескольких нуклеотидах последовательностей ITS . Таким образом, использование исключительно данных штрих-кода ITS для идентификации таких пар или комплексов видов может скрыть фактическое разнообразие и может привести к неправильной идентификации, если не сопровождается исследованием морфологических и экологических особенностей и/или сравнением дополнительных диагностических генетических маркеров . [ 19 ] [ 24 ] [ 26 ] [ 27 ] Для некоторых таксонов ITS (или его часть ITS2 ) недостаточно изменчива, как штрих-код ДНК грибов, как, например, было показано у Aspergillus , Cladosporium , Fusarium и Penicillium . [ 28 ] [ 29 ] [ 30 ] [ 31 ] Таким образом, попытки определить универсально применимое пороговое значение изменчивости ITS , которое отделяет внутривидовую изменчивость от межвидовой (межвидовой) изменчивости, остаются тщетными. [ 25 ]
Тем не менее, вероятность правильной идентификации вида по региону ITS высока у Dikarya , и особенно у Basidiomycota , где даже части ITS1 часто бывает достаточно для идентификации вида. [ 32 ] Однако его способность к распознаванию частично заменяется способностью РНК-полимеразы II ДНК-направленной субъединицы RPB1 (см. Также ниже). [ 2 ]
Из-за недостатков ITS' как первичного штрих-кода ДНК грибов была выражена необходимость создания второго маркера штрих-кода ДНК. [ 9 ] Было предпринято несколько попыток установить другие генетические маркеры, которые могли бы служить дополнительными штрих-кодами ДНК. [ 19 ] [ 33 ] [ 34 ] аналогично ситуации у растений , где пластидные гены rbcL , matK и trnH-psbA , а также ядерный ITS часто используются в комбинации для штрих-кодирования ДНК. [ 35 ]
Фактор трансляционной элонгации 1α (TEF1α) – вторичный грибковый штрих-код.
[ редактировать ]Фактор трансляционной элонгации 1α является частью эукариотического комплекса фактора элонгации 1 , основная функция которого заключается в содействии удлинению аминокислотной цепи полипептида во время трансляции процесса экспрессии генов . [ 36 ]
Стилоу и др. (2015) исследовали ген TEF1α , среди ряда других, как потенциальный генетический маркер для штрих-кодирования ДНК грибов. Ген TEF1α , кодирующий фактор трансляционной элонгации 1α, обычно считается имеющим медленную скорость мутаций , и поэтому он обычно лучше подходит для исследования старых расщеплений, находящихся на более глубокой стадии филогенетической истории группы организмов. Несмотря на это, авторы приходят к выводу, что TEF1α является наиболее многообещающим кандидатом на роль дополнительного маркера штрих-кода ДНК у грибов, поскольку он также имеет области последовательности с более высокой частотой мутаций. [ 19 ] После этого была создана справочная база данных с контролем качества, которая была объединена с ранее существовавшей базой данных ISHAM-ITS для штрих-кодов ДНК ITS грибов. [ 1 ] для формирования базы данных ISHAM. [ 37 ]
TEF1α успешно использовался для идентификации нового вида Cantharellus из Техаса и отличия его от морфологически сходного вида. [ 38 ] Однако в родах Ochroconis и Verruconis (Sympoventuriaceae, Venturiales) маркер не позволяет различить все виды. [ 39 ] TEF1α также использовался в филогенетическом анализе на уровне рода, например, в случае Cantharellus. [ 40 ] и энтомопатогенная боверия , [ 41 ] и для филогенетики рано расходящихся грибных линий. [ 42 ]
Праймеры
[ редактировать ]Праймеры TEF1α , используемые при широкомасштабном скрининге характеристик кандидатов в гены штрих-кода ДНК Stielow et al. (2015) использовали прямой праймер EF1-983F с последовательностью 5'-GCYCCYGGHCAYCGTGAYTTYAT-3' и обратный праймер EF1-1567R с последовательностью 5'-ACHGTRCCRATACCACCRATCTT-3' . [ 41 ] Кроме того, был разработан ряд новых праймеров, пара праймеров выделена жирным шрифтом, что привело к высокому среднему успеху амплификации - 88%: [ 19 ]
|
Форвардные праймеры:
|
Обратные праймеры:
|
Праймеры, используемые для исследования Rhizophydiales и особенно Batrachochytrium dendrobatidis , возбудителя амфибий, представляют собой прямой праймер tef1F с нуклеотидной последовательностью 5'-TACAARTGYGGTGGTATYGACA-3' и обратный праймер tef1R с последовательностью 5'-ACNGACTTGACYTCAGTRGT-3' . [ 43 ] Эти праймеры также успешно амплифицировали большинство видов Cantharellus , исследованных Buyck et al. (2014), за исключением нескольких видов, для которых были разработаны более специфичные праймеры: прямой праймер tef-1Fcanth с последовательностью 5'-AGCATGGGTDCTYGACAAG-3' и обратный праймер tef-1Rcanth с последовательностью 5'-CCAATYTTRTAYACATCYTGGAG-3' . [ 40 ]
Домен D1/D2 рибосомальной РНК LSU
[ редактировать ]Домен D1/D2 является частью РНК большой ядерной субъединицы ( 28S ) рибосомальной и поэтому расположен в том же кластере генов рибосомных тандемных повторов, что и внутренний транскрибируемый спейсер ( ITS ). Но в отличие от некодирующих последовательностей ITS, домен D1/D2 содержит кодирующую последовательность. Имея около 600 пар оснований, это примерно такая же длина нуклеотидной последовательности, как и ITS . [ 44 ] что делает амплификацию и секвенирование довольно простыми, и это преимущество привело к накоплению большого количества данных о последовательностях D1/D2, особенно для дрожжей . [ 3 ] [ 7 ] [ 44 ]
Что касается молекулярной идентификации базидиомицетов, D1/D2 (или ITS ) можно использовать отдельно. [ 44 ] Однако Фелл и др. (2000) и Скорцетти и др. (2002) рекомендуют комбинированный анализ регионов D1/D2 и ITS , [ 3 ] [ 44 ] практика, которая позже стала стандартной, требовала информации для описания новых таксонов аско- и базидиомицетов дрожжей. [ 14 ] При попытке идентифицировать ранние дивергентные линии грибов исследование Schoch et al. (2012), сравнивая эффективность идентификации различных генетических маркеров, показали, что большая субъединица (а также малая субъединица ) рибосомальной РНК работает лучше, чем ITS или RPB1 . [ 2 ]
Праймеры
[ редактировать ]Для базидиомицетовых дрожжей используется прямой праймер F63 с последовательностью 5'-GCATATCAATAAGCGGAGGAAAAG-3' и обратный праймер LR3 с последовательностью 5'-GGTCCGTGTTTTCAAGACGG-3' успешно использовался для ПЦР-амплификации домена D1/D23. [ 3 ] Домен D1/D2 аскомицетовых дрожжей, таких как Candida, можно амплифицировать с помощью прямого праймера NL-1 (такого же, как F63 ) и обратного праймера NL-4 (такого же, как LR3 ). [ 6 ]
Субъединица РНК-полимеразы II RPB1
[ редактировать ]
Субъединица РНК-полимеразы II RPB1 является самой крупной субъединицей РНК-полимеразы II . У Saccharomyces cerevisiae он кодируется геном RPO21 . [ 46 ] Успех ПЦР- амплификации RPB1 во многом зависит от таксона: от 70 до 80% у Ascomycota до 14% у рано расходящихся грибковых линий. [ 2 ] Помимо ранних дивергентных линий, RPB1 имеет высокий уровень идентификации видов во всех группах грибов. У богатой видами Pezizomycotina он даже превосходит ITS. [ 2 ]
В исследовании, сравнивающем эффективность идентификации четырех генов, RPB1 был одним из наиболее эффективных генов при объединении двух генов в анализе: комбинированный анализ либо с ITS , либо с большой субъединицей рибосомальной РНК дал наивысший успех идентификации. [ 2 ]
В других исследованиях также использовался RPB2 , вторая по величине субъединица РНК-полимеразы II, например, для изучения филогенетических взаимоотношений между видами рода Cantharellus. [ 40 ] или для филогенетического исследования, проливающего свет на взаимоотношения между рано расходящимися линиями в грибном царстве. [ 42 ]
Праймеры
[ редактировать ]Праймеры, успешно амплифицирующие RPB1, особенно у Ascomycota, представляют собой прямой праймер RPB1-Af с последовательностью 5'-GARTGYCCDGGDCAYTTYGG-3' и обратный праймер RPB1-Ac-RPB1-Cr с последовательностью 5'-CCNGCDATNTCRTTRTTCCATRTA-3' . [ 2 ]
Межгенный спейсер (IGS) генов рибосомальной РНК
[ редактировать ]Межгенный спейсер ( IGS ) — это область некодирующей ДНК между отдельными тандемными повторами кластера рибосомальных генов в ядерном геноме , в отличие от внутреннего транскрибируемого спейсера (ITS), который расположен внутри этих тандемных повторов.
IGS успешно применяется для дифференциации штаммов Xanthophyllomyces dendrorous. [ 47 ] а также для видового различия психрофильного рода Мракия ( Cystofilobasidiales ) . [ 48 ] Благодаря этим результатам IGS был рекомендован в качестве генетического маркера для дополнительной дифференциации (наряду с D1/D2 и ITS ) близкородственных видов и даже штаммов внутри одного вида базидиомицетных дрожжей. [ 3 ]
Недавнее открытие дополнительных некодирующих генов РНК в области IGS некоторых базидиомицетов предостерегает от некритического использования последовательностей IGS для штрих-кодирования ДНК и филогенетических целей. [ 49 ]
Другие генетические маркеры
[ редактировать ]Ген субъединицы I цитохром с-оксидазы ( COI ) превосходит ITS при штрих-кодировании ДНК видов Penicillium (Ascomycota) с видоспецифичными штрих-кодами для 66% исследованных видов против 25% в случае ITS . Кроме того, часть гена β-тубулина А ( BenA ) демонстрирует более высокую таксономическую разрешающую способность при различении видов Penicillium по сравнению с COI и ITS . [ 50 ] в близкородственном комплексе Aspergillus niger Однако COI недостаточно изменчив для различения видов. [ 51 ] У Fusarium . COI обнаруживает паралоги во многих случаях , а гомологичные копии недостаточно изменчивы, чтобы различать виды [ 52 ]
COI также плохо работает при идентификации базидиомикотов ржавчины порядка из - Pucciniales за присутствия интронов . Даже когда препятствие интронов преодолено, ITS и LSU рРНК ( 28S ) превосходят COI в качестве маркера штрих-кода ДНК. [ 53 ] В подразделении Agaricomycotina был плохим успех ПЦР-амплификации COI , даже при использовании нескольких комбинаций праймеров. Успешно секвенированные образцы COI также включали интроны и возможные паралогичные копии, как сообщалось для Fusarium . [ 52 ] [ 54 ] Было обнаружено, что Agaricus bisporus содержит до 19 интронов, что делает ген COI этого вида самым длинным из зарегистрированных - 29 902 нуклеотида. [ 55 ] Помимо существенных проблем с секвенированием COI , COI и ITS обычно одинаково хорошо различают грибы базидиомикоты. [ 54 ]
Топоизомераза I ( TOP1 ) была исследована в качестве дополнительного кандидата на штрих-код ДНК Lewis et al. (2011) на основе протеомных данных с разработанной универсальной парой праймеров. [ 33 ] впоследствии были протестированы на реальных образцах Stielow et al. (2015). Прямой праймер TOP1_501-F с последовательностью 5'-TGTAAAACGACGGCCAGT-ACGAT-ACTGCCAAGGTTTTCCGTACHTACAACGC-3' (где первая часть обозначает хвост универсального переднего праймера M13, вторая часть состоит из ACGAT - спейсера, а третья часть - собственно праймера) и переверните праймер TOP1_501-R с помощью 5'-CAGGAAACAGCTATGA-CCCAGTCCTCGTCAACWGACTTRATRGCCCA-3' (первая часть обозначает хвост универсального обратного праймера M13, вторая часть - фактический обратный праймер TOP1) амплифицирует фрагмент длиной примерно 800 пар оснований. [ 19 ]
Было обнаружено, что TOP1 является многообещающим маркером-кандидатом ДНК-штрих-кода для аскомицетов, где он может различать виды рода Fusarium и Penicillium , у которых первичный штрих-код ITS работает плохо. Однако плохой успех амплификации с помощью универсальных праймеров TOP1 наблюдается у рано расходящихся грибковых линий и базидиомицетов, за исключением Pucciniomycotina (где успех ITS PCR плохой). [ 19 ]
Как и TOP1 , фосфоглицераткиназа ( PGK ) была среди генетических маркеров, исследованных Lewis et al. (2011) и Стилоу и др. (2015) в качестве потенциальных дополнительных штрих-кодов ДНК грибов. Разработан ряд универсальных праймеров. [ 33 ] с парой праймеров PGK533, амплифицирующей фрагмент длиной около 1000 пар оснований, что является наиболее успешным для большинства грибов, за исключением базидиомицетов. Как и TOP1 , PGK превосходит ITS в видовой дифференциации в родах аскомицетов, таких как Penicillium и Fusarium , и как PGK , так и TOP1 работают так же хорошо, как TEF1α, в различении близкородственных видов в этих родах. [ 19 ]
Приложения
[ редактировать ]Безопасность пищевых продуктов
[ редактировать ]Гражданский научный проект исследовал консенсус между маркировкой сушеных, коммерчески продаваемых грибов и результатами штрих-кодирования ДНК этих грибов. Было обнаружено, что все образцы были правильно маркированы. Однако препятствием была ненадежность эталонных баз данных ITS с точки зрения уровня идентификации, поскольку две базы данных (GenBank и UNITE), использованные для сравнения последовательностей ITS, дали разные результаты идентификации в некоторых образцах. [ 56 ] [ 57 ]
Правильная маркировка грибов, предназначенных для употребления в пищу, также была исследована Raja et al. (2016), которые использовали область ITS для штрих-кодирования ДНК сушеных грибов, порошков мицелия и с пищевыми добавками капсул . Только в 30% из 33 образцов на этикетке продукта было правильно указано биномиальное название гриба. Еще в 30% родовое название было правильным, но видовой эпитет не совпадал, а в 15% случаев даже родовое название биномиального названия, указанного на этикетке продукта, не совпадало с результатом полученного штрих-кода ИТС . Для остальных 25% образцов последовательность ITS получить не удалось. [ 58 ]
Сян и др. (2013) показали, что с помощью ITS коммерчески ценные гусеничные грибы Ophiocordyceps sinensis и их поддельные версии ( O. nutans , O. robertsii , Cordyceps cicadae , C. Gunnii , C. militaris и растение Ligularia hodgsonii -последовательностей можно получить ). достоверно идентифицированы до видового уровня. [ 59 ]
Патогенные грибы
[ редактировать ]Исследование Ви Хоанга и др. (2019) сосредоточились на точности идентификации патогенных грибов с использованием как первичных ( ITS ), так и вторичных ( TEF1α ) маркеров штрих-кода. Их результаты показывают, что у Диутина (выделенного вида Candida [ 60 ] ) и Pichia , идентификация видов проста с помощью ITS или TEF1α , а также с помощью их комбинации. В комплексе Lodderomyces , который содержит три из пяти наиболее распространенных патогенных видов Candida ( C. albicans , C. dubliniensis и C. parapsilosis ), ITS не смог отличить Candida ortopsilosis и C. parapsilosis , которые являются частью Candida parapsilosis. комплекса близкородственных видов. [ 61 ] TEF1α , напротив, позволил идентифицировать все исследованные виды клады Lodderomyces . Аналогичные результаты были получены для видов Scedosporium , которые относятся к широкому спектру локализованных инвазионных заболеваний: ITS не мог различить S. apiospermum и S. boydii , тогда как с помощью TEF1α можно было точно идентифицировать все исследованные виды этого рода. Таким образом, это исследование подчеркивает полезность применения более чем одного маркера штрих-кодирования ДНК для идентификации видов грибов. [ 62 ]
Сохранение культурного наследия
[ редактировать ]Штрих-кодирование ДНК грибов успешно применяется для исследования явлений лисиц , что является серьезной проблемой при сохранении бумажных документов . Секейра и др. (2019) секвенировали ITS из лисьих пятен и обнаружили Chaetomium globosum , Ch. мурорум , гл. nigricolor , Chaetomium sp., Eurotium rubrum , Myxotrichum deflexum , Penicillium chrysogenum , P. citrinum , P. commune , Penicillium sp. и Stachybotryschartarum обитали в исследованных бумажных пятнах. [ 63 ]
Другое исследование изучало грибы, которые действуют как агенты биоразрушения в Старом соборе Коимбры , являющемся частью Университета Коимбры , ЮНЕСКО объекта Всемирного наследия . Секвенируя штрих-код ITS десяти образцов с помощью классического метода Сэнгера , а также с помощью Illumina методов секвенирования нового поколения , они идентифицировали 49 видов грибов. Aspergillus versicolor , Cladosporium cladosporioides , C. sphaerospermum , C. tenuissimum , Epicoccum nigrum , Parengyodontium album , Penicillium brevicompactum , P. Crustosum , P. glabrum , Talaromyces amestolkiae и T. stallii . Наиболее распространенными видами, выделенными из образцов, были [ 64 ]
Другое исследование, посвященное объектам культурного наследия, изучало разнообразие грибков на холсте Паулы Рего с использованием ITS2 субрегиона маркера ITS . Всего было обнаружено 387 OTU (предполагаемых видов) из 117 родов 13 различных классов грибов. [ 65 ]
См. также
[ редактировать ]- ДНК-штрихкодирование
- Штрих-кодирование микробной ДНК
- Штрих-кодирование ДНК пыльцы
- Штрих-кодирование ДНК при оценке диеты
- Консорциум штрих-кода жизни
Ссылки
[ редактировать ]- ^ Jump up to: а б с Ирини Л., Серена С., Гарсиа-Эрмосо Д., Арабацис М., Деснос-Оливье М., Ву Д. и др. (май 2015 г.). «Справочная база данных штрих-кодирования ДНК Международного общества микологии человека и животных (ISHAM) — ITS — стандартный инструмент с контролируемым качеством для рутинной идентификации патогенных грибов человека и животных» (PDF) . Медицинская микология . 53 (4): 313–37. дои : 10.1093/mmy/myv008 . ПМИД 25802363 . Архивировано (PDF) из оригинала 26 июля 2022 года . Проверено 6 июня 2024 г.
- ^ Jump up to: а б с д и ж г час я Шох К.Л., Зайферт К.А., Хундорф С., Роберт В., Спудж Дж.Л., Левеск К.А., Чен В. (апрель 2012 г.). «Область внутреннего транскрибируемого спейсера ядерной рибосомы (ITS) как универсальный маркер штрих-кода ДНК для грибов» (PDF) . Труды Национальной академии наук Соединенных Штатов Америки . 109 (16): 6241–6. дои : 10.1073/pnas.1117018109 . ПМК 3341068 . ПМИД 22454494 . Архивировано (PDF) из оригинала 7 июля 2020 года . Проверено 12 марта 2020 г.
- ^ Jump up to: а б с д и ж Фелл Дж.В., Бёкхаут Т., Фонсека А., Скорцетти Г., Статцелл-Толлман А. (май 2000 г.). «Биоразнообразие и систематика базидиомицетов, определенные с помощью анализа последовательности домена D1/D2 крупных субъединиц рДНК» . Международный журнал систематической и эволюционной микробиологии . 50, ч. 3 (3): 1351–1371. дои : 10.1099/00207713-50-3-1351 . ПМИД 10843082 . S2CID 44194598 .
- ^ Брунс Т.Д., Уайт Т.Дж., Тейлор Дж.В. (1991). «Молекулярная систематика грибов». Ежегодный обзор экологии и систематики . 22 (1): 525–564. doi : 10.1146/annurev.es.22.110191.002521 . ПМИД 12702331 .
- ^ Месснер Р., Приллингер Х., Ибл М., Гиммлер Г. (1995). «Последовательности рибосомальных генов и внутренние транскрибируемые спейсеры перемещают три гриба-паразита растений, Eremothecium ashbyi , Ashbya gossypii и Nematospora coryli , в сторону Saccharomyces cerevisiae » . Журнал общей и прикладной микробиологии . 41 : 31–42. дои : 10.2323/jgam.41.31 . Архивировано из оригинала 4 мая 2019 года . Проверено 9 апреля 2020 г.
- ^ Jump up to: а б Курцман CP, Робнетт CJ (май 1997 г.). «Идентификация клинически важных аскомицетов на основе дивергенции нуклеотидов на 5'-конце гена рибосомальной ДНК с большой субъединицей (26S)» . Журнал клинической микробиологии . 35 (5): 1216–23. doi : 10.1128/JCM.35.5.1216-1223.1997 . ПМК 232732 . ПМИД 9114410 .
- ^ Jump up to: а б Курцман CP, Робнетт CJ (май 1998 г.). «Идентификация и филогения аскомицетовых дрожжей на основе анализа частичных последовательностей рибосомальной ДНК большой субъединицы ядра (26S)» . Антони ван Левенгук . 73 (4): 331–71. дои : 10.1023/а:1001761008817 . ПМИД 9850420 . S2CID 29373623 . Архивировано из оригинала 31 марта 2023 года . Проверено 9 апреля 2020 г.
- ^ Курцман CP, Робнетт CJ (октябрь 1998 г.). «Три новых вида дрожжей рода Candida , связанных с насекомыми » . Канадский журнал микробиологии . 44 (10): 965–73. дои : 10.1139/w98-085 . ПМИД 9933915 . Архивировано из оригинала 22 июля 2023 года . Проверено 12 марта 2020 г.
- ^ Jump up to: а б с Зайферт К.А. (май 2009 г.). «Прогресс в направлении ДНК-баркодирования грибов». Ресурсы молекулярной экологии . 9 Приложение с1 (Приложение 1): 83–9. дои : 10.1111/j.1755-0998.2009.02635.x . ПМИД 21564968 .
- ^ Хеберт П.Д. , Цивинска А., Болл С.Л., деВаард Дж.Р. (февраль 2003 г.). «Биологическая идентификация посредством штрих-кодов ДНК» . Слушания. Биологические науки . 270 (1512): 313–21. дои : 10.1098/рспб.2002.2218 . ПМЦ 1691236 . ПМИД 12614582 .
- ^ Нильссон Р.Х., Райберг М., Абаренков К., Шёквист Э., Кристианссон Э. (июль 2009 г.). «Регион ITS как цель для характеристики грибковых сообществ с использованием новых технологий секвенирования» . Письма FEMS по микробиологии . 296 (1): 97–101. дои : 10.1111/j.1574-6968.2009.01618.x . ПМИД 19459974 .
- ^ Jump up to: а б Бегероу Д., Нильссон Х., Унтерсехер М., Майер В. (июнь 2010 г.). «Современное состояние и перспективы штрих-кодирования ДНК грибов и процедур быстрой идентификации». Прикладная микробиология и биотехнология . 87 (1): 99–108. дои : 10.1007/s00253-010-2585-4 . ПМИД 20405123 . S2CID 25172732 .
- ^ Агерер Р., Аммирати Дж., Барони Т.Дж., Бланц П., Куртекюисс Р.Э., Дежарден Д.Е. и др. (2000). «Открытое письмо научному сообществу микологов» . Прикладная экология почв . 15 (3): 295–298. дои : 10.1016/S0929-1393(00)00076-7 .
- ^ Jump up to: а б с д Сюй Дж (ноябрь 2016 г.). «Штрих-кодирование ДНК грибов» . Геном . 59 (11): 913–932. дои : 10.1139/gen-2016-0046 . ПМИД 27829306 .
- ^ Jump up to: а б Вурцбахер С., Ларссон Е., Бенгтссон-Пальме Дж., Ван ден Вингаерт С., Свантессон С., Кристианссон Е. и др. (январь 2019 г.). «Представляем штрих-кодирование рибосомальных тандемных повторов для грибов» . Ресурсы молекулярной экологии . 19 (1): 118–127. дои : 10.1111/1755-0998.12944 . ПМИД 30240145 . S2CID 52309438 . Архивировано из оригинала 15 августа 2020 года . Проверено 7 июня 2020 г.
- ^ Нильссон, Рольф Хенрик; Ларссон, Карл-Хенрик; Тейлор, Энди Ф.С.; Бенгтссон-Пальме, Йохан; Джеппесен, Томас С.; Шигель, Дмитрий; Кеннеди, Питер; Пикард, Кэтрин; Глёкнер, Фрэнк Оливер (8 января 2019 г.). «База данных UNITE для молекулярной идентификации грибов: работа с темными таксонами и параллельными таксономическими классификациями» . Исследования нуклеиновых кислот . 47 (Д1): Д259–Д264. дои : 10.1093/nar/gky1022 . ISSN 0305-1048 . ПМК 6324048 . ПМИД 30371820 .
- ^ Jump up to: а б с д Уайт Т.Дж., Брунс Т., Ли С.Дж., Тейлор Дж. (1990). «Амплификация и прямое секвенирование генов рибосомальной РНК грибов для филогенетики». В Иннис М.А., Гельфанд Д.Х., Снинский Дж.Дж., Уайт Т.Дж. (ред.). Протоколы ПЦР: Руководство по методам и применению . Нью-Йорк: Academic Press, Inc., стр. 315–322. Архивировано из оригинала 6 июня 2024 года . Проверено 12 марта 2020 г.
- ^ Тодзю Х., Танабэ А.С., Ямамото С., Сато Х. (2012). «Праймеры ITS с высокой степенью покрытия для идентификации аскомицетов и базидиомицетов на основе ДНК в образцах окружающей среды» . ПЛОС ОДИН . 7 (7): e40863. Бибкод : 2012PLoSO...740863T . дои : 10.1371/journal.pone.0040863 . ПМК 3395698 . ПМИД 22808280 .
- ^ Jump up to: а б с д и ж г час я Стилов Дж.Б., Левеск К.А., Зейферт К.А., Мейер В., Ирини Л., Смитс Д. и др. (декабрь 2015 г.). «Один гриб, какие гены? Разработка и оценка универсальных праймеров для потенциальных вторичных штрих-кодов ДНК грибов» . Персония . 35 : 242–63. дои : 10.3767/003158515X689135 . ПМЦ 4713107 . ПМИД 26823635 .
- ^ Бельмен Э., Карлсен Т., Брохманн С., Куассак Э., Таберле П., Каузеруд Х. (июль 2010 г.). «ITS как штрих-код ДНК окружающей среды для грибов: подход in silico выявляет потенциальные ошибки ПЦР» . БМК Микробиология . 10 (189): 189. дои : 10.1186/1471-2180-10-189 . ПМК 2909996 . ПМИД 20618939 .
- ^ Смит М.Э., Духан Г.В., Риццо Д.М. (декабрь 2007 г.). «Внутривидовые и внутриспорокарпические ITS-вариации эктомикоризных грибов, оцененные с помощью секвенирования рДНК спорокарпов и объединенных эктомикоризных корней из лесного массива Quercus » . Микориза . 18 (1): 15–22. Бибкод : 2007Mycor..18...15S . дои : 10.1007/s00572-007-0148-z . ПМИД 17710446 . S2CID 195072428 . Архивировано из оригинала 6 июня 2024 года . Проверено 21 апреля 2020 г.
- ^ Линднер Д.Л., Баник М.Т. (2011). «Внутригеномные вариации в регионе ITS рДНК скрывают филогенетические отношения и завышают оценки операционных таксономических единиц рода Laetiporus ». Микология . 103 (4): 731–40. дои : 10.3852/10-331 . ПМИД 21289107 . S2CID 21154111 .
- ^ Ковач ГМ, Балаж ТК, Калонже ФД, Мартин MP (2011). «Разнообразие пустынных трюфелей Терфеция : новые виды и весьма изменчивый комплекс видов с внутриспорокарпической гетерогенностью ITS нрДНК» (PDF) . Микология . 103 (4): 841–53. дои : 10.3852/10-312 . ПМИД 21289106 . S2CID 22648182 . Архивировано (PDF) из оригинала 6 июня 2024 года . Проверено 9 апреля 2020 г.
- ^ Jump up to: а б Kiss L (июль 2012 г.). «Границы последовательностей внутренних транскрибируемых спейсеров (ITS) ядерной рибосомальной ДНК как видовых штрих-кодов грибов» (PDF) . Труды Национальной академии наук Соединенных Штатов Америки . 109 (27): E1811, ответ автора E1812. Бибкод : 2012PNAS..109E1811K . дои : 10.1073/pnas.1207143109 . ПМК 3390822 . ПМИД 22715287 . Архивировано (PDF) из оригинала 23 марта 2020 г. Проверено 14 апреля 2020 г.
- ^ Jump up to: а б Нильссон Р.Х., Кристианссон Э., Райберг М., Халленберг Н., Ларссон К.Х. (май 2008 г.). «Внутривидовая изменчивость ITS в королевстве грибов, выраженная в международных базах данных последовательностей, и ее значение для идентификации молекулярных видов» . Эволюционная биоинформатика онлайн . 4 : 193–201. дои : 10.4137/EBO.S653 . ПМК 2614188 . ПМИД 19204817 .
- ^ Сюй Дж., Вилгалис Р., Митчелл Т.Г. (октябрь 2000 г.). «Множественные генеалогии генов показывают недавнюю дисперсию и гибридизацию патогенного для человека грибка Cryptococcus neoformans ». Молекулярная экология . 9 (10): 1471–81. Бибкод : 2000MolEc...9.1471X . дои : 10.1046/j.1365-294x.2000.01021.x . ПМИД 11050543 . S2CID 18291790 .
- ^ Стокингер Х., Крюгер М., Шюсслер А. (июль 2010 г.). «Штрих-кодирование ДНК арбускулярных микоризных грибов» (PDF) . Новый фитолог . 187 (2): 461–74. дои : 10.1111/j.1469-8137.2010.03262.x . ПМИД 20456046 . Архивировано (PDF) из оригинала 6 июня 2024 года . Проверено 6 июня 2024 г.
- ^ Гейзер Д.М., Клих М.А., Фрисвад Дж.К., Петерсон С.В., Варга Дж., Самсон Р.А. (2007). «Современное состояние распознавания и идентификации видов Aspergillus » . Исследования по микологии . 59 : 1–10. дои : 10.3114/sim.2007.59.01 . ПМК 2275194 . ПМИД 18490947 .
- ^ Шуберт К., Гроеневальд Дж. З., Браун У., Дейкстерхейс Дж., Старинк М., Хилл К.Ф. и др. (2007). «Биоразнообразие комплекса Cladosporium herbarum (Davidiellaceae, Capnodiales) со стандартизацией методов систематики и диагностики Cladosporium» . Исследования по микологии . 58 : 105–56. дои : 10.3114/sim.2007.58.05 . ПМК 2104742 . ПМИД 18490998 .
- ^ О'Доннелл К., Цигельник Э. (февраль 1997 г.). «Два дивергентных внутригеномных типа рДНК ITS2 в монофилетической линии гриба Fusarium неортологичны» . Молекулярная филогенетика и эволюция . 7 (1): 103–16. Бибкод : 1997МОЛПЭ...7..103О . дои : 10.1006/mpev.1996.0376 . ПМИД 9007025 .
- ^ Скубо П., Фрисвад Дж. К., Тейлор Дж. В., Лауритсен Д., Бойсен М., Россен Л. (1999). «Филогенетический анализ нуклеотидных последовательностей из области ITS тервертициллятных видов Penicillium » . Микологические исследования . 103 (7): 873–881. дои : 10.1017/S0953756298007904 . Архивировано из оригинала 6 июня 2024 года . Проверено 16 марта 2020 г.
- ^ Осмундсон Т.В., Роберт В.А., Шох К.Л., Бейкер Л.Дж., Смит А., Робич Г. и др. (2013). «Заполнение пробелов в знаниях о биоразнообразии макрогрибов: вклад и оценка проекта секвенирования штрих-кода ДНК коллекции гербария» . ПЛОС ОДИН . 8 (4): e62419. Бибкод : 2013PLoSO...862419O . дои : 10.1371/journal.pone.0062419 . ПМК 3640088 . ПМИД 23638077 .
- ^ Jump up to: а б с Льюис К.Т., Билху С., Роберт В., Эберхардт Ю., Соке С., Зейферт К.А., Левеск К.А. (2011). «Идентификация целей штрих-кода ДНК грибов и праймеров для ПЦР на основе семейств белков Pfam и таксономической иерархии» (PDF) . Открытый журнал прикладной информатики . 5 (дополнение 1–М5): 30–44. дои : 10.2174/1874136301005010030 . Архивировано (PDF) из оригинала 19 октября 2021 года . Проверено 14 апреля 2020 г.
- ^ Винсент Роберт Л., Сёке С., Эберхардт У., Кардинали Г., Мейер В., Зейферт К.А., Левеск К.А., Льюис К.Т. (2011). «В поисках общего и надежного штрих-кода ДНК грибов» (PDF) . Открытый журнал прикладной информатики . 5 (дополнение 1–М6): 45–61. дои : 10.2174/1874136301005010045 . Архивировано (PDF) из оригинала 23 сентября 2020 г. Проверено 14 апреля 2020 г.
- ^ Кресс WJ (2017). «Штрих-коды ДНК растений: применение сегодня и в будущем» . Журнал систематики и эволюции . 55 (4): 291–307. дои : 10.1111/jse.12254 .
- ^ Сасикумар А.Н., Перес В.Б., Кинзи Т.Г. (2012). «Множество ролей эукариотического комплекса фактора элонгации 1» . Междисциплинарные обзоры Wiley. РНК . 3 (4): 543–55. дои : 10.1002/wrna.1118 . ПМЦ 3374885 . ПМИД 22555874 .
- ^ Мейер В., Ирини Л., Хоанг М.Т., Роберт В., Гарсия-Эрмосо Д., Деснос-Олливер М. и др. (март 2019 г.). «Создание базы данных по фактору элонгации трансляции штрих-кода вторичной ДНК грибов 1α (TEF1α)» . Геном . 62 (3): 160–169. дои : 10.1139/gen-2018-0083 . hdl : 1807/93998 . ПМИД 30465691 .
- ^ Буйк Б., Круо С., Кулу А., Хофстеттер В. (2011). « Cantharellus texensis sp. nov. из Техаса, южный двойник C. cinnabarinus , выявленный с помощью данных о последовательности tef-1». Микология . 103 (5): 1037–46. дои : 10.3852/10-261 . ПМИД 21558500 . S2CID 29384238 .
- ^ Самерпитак К., Герритс ван ден Энде Б.Х., Стилов Дж.Б., Менкен С.Б., де Хоог Г.С. (февраль 2016 г.). «Штрих-кодирование и распознавание видов условно-патогенных микроорганизмов у Ochroconis и Verruconis » (PDF) . Грибковая биология . 120 (2): 219–30. Бибкод : 2016FunB..120..219S . дои : 10.1016/j.funbio.2015.08.010 . ПМИД 26781378 . Архивировано (PDF) из оригинала 18 февраля 2019 года . Проверено 14 апреля 2020 г.
- ^ Jump up to: а б с Буйк Б., Кауфф Ф., Эйссартье Г., Кулу А., Хофстеттер В. (2014). «Мультилокусная филогения Cantharellus (Cantharellales, Agaricomycetidae) во всем мире» (PDF) . Грибное разнообразие . 64 : 101–121. дои : 10.1007/s13225-013-0272-3 . S2CID 11264350 . Архивировано (PDF) из оригинала 6 июня 2024 года . Проверено 12 марта 2020 г.
- ^ Jump up to: а б Ренер С.А., Бакли Э. (2005). « Филогения Beauveria , выведенная на основе ядерных последовательностей ITS и EF1-альфа: свидетельства загадочной диверсификации и связи с телеоморфами Кордицепса ». Микология . 97 (1): 84–98. дои : 10.1080/15572536.2006.11832842 . ПМИД 16389960 . S2CID 22209059 .
- ^ Jump up to: а б Джеймс Т., Кауфф Ф., Шох К.Л., Матени П.Б., Хофстеттер В., Кокс С.Дж. и др. (октябрь 2006 г.). «Реконструкция ранней эволюции грибов с использованием филогении из шести генов» . Природа . 443 (7113): 818–22. Бибкод : 2006Natur.443..818J . дои : 10.1038/nature05110 . ПМИД 17051209 . S2CID 4302864 . Архивировано из оригинала 11 июля 2022 года . Проверено 9 апреля 2020 г.
- ^ Морхаус Э.А., Джеймс Тай, Гэнли А.Р., Вилгалис Р., Бергер Л., Мерфи П.Дж., Лонгкор Дж.Е. (февраль 2003 г.). «Мультилокусное типирование последовательностей позволяет предположить, что хитридийный патоген амфибий представляет собой недавно появившийся клон». Молекулярная экология . 12 (2): 395–403. Бибкод : 2003MolEc..12..395M . дои : 10.1046/j.1365-294X.2003.01732.x . ПМИД 12535090 . S2CID 13448384 .
- ^ Jump up to: а б с д Скорцетти Дж., Фелл Дж.В., Фонсека А., Статцелл-Толлман А. (декабрь 2002 г.). «Систематика базидиомицетов: сравнение большой субъединицы D1/D2 и внутренних транскрибируемых спейсерных областей рДНК». Исследование дрожжей FEMS . 2 (4): 495–517. дои : 10.1111/j.1567-1364.2002.tb00117.x . ПМИД 12702266 .
- ^ Армаш К.Дж., Миттервегер С., Мейнхарт А., Крамер П. (февраль 2005 г.). «Структуры полной РНК-полимеразы II и ее подкомплекса Rpb4/7» (PDF) . Журнал биологической химии . 280 (8): 7131–4. doi : 10.2210/pdb1wcm/pdb . ПМИД 15591044 . Архивировано (PDF) из оригинала 24 июля 2018 года . Проверено 13 марта 2020 г.
- ^ Стратерн Дж., Малагон Ф., Ирвин Дж., Готте Д., Шафер Б., Киреева М. и др. (январь 2013 г.). «Правильность транскрипции: мутации RPB1 (RPO21), которые увеличивают проскальзывание транскрипции у S. cerevisiae » . Журнал биологической химии . 288 (4): 2689–99. дои : 10.1074/jbc.M112.429506 . ПМЦ 3554935 . ПМИД 23223234 .
- ^ Фелл Дж.В., Блатт ГМ (июль 1999 г.). «Разделение штаммов дрожжей Xanthophyllomyces dendrorous и Phaffia rhodozyma на основе анализа последовательностей рДНК IGS и ITS» . Журнал промышленной микробиологии и биотехнологии . 23 (1): 677–81. дои : 10.1038/sj.jim.2900681 . ПМИД 10455500 . S2CID 22545332 .
- ^ Диас М.Р., Фелл Дж.В. (январь 2000 г.). «Молекулярный анализ областей IGS и ITS рДНК психрофильных дрожжей рода Mrakia ». Антони ван Левенгук . 77 (1): 7–12. дои : 10.1023/А:1002048008295 . ПМИД 10696872 . S2CID 41560178 .
- ^ Альм Розенблад М., Ларссон Э., Уокер А., Тонгкланг Н., Вурцбахер Н., Нильссон Р.Х. (2022). «Доказательства наличия дополнительных некодирующих генов РНК в рДНК грибов» . MycoKeys (90): 203–213. дои : 10.3897/mycokeys.90.84866 . ПМЦ 9849065 . ПМИД 36760425 .
- ^ Зайферт К.А., Самсон Р.А., Деваард Дж.Р., Хубракен Дж., Левеск К.А., Монкальво Дж.М. и др. (март 2007 г.). «Перспективы идентификации грибов с использованием штрих-кодов ДНК CO1 Penicillium на примере » (PDF) . Труды Национальной академии наук Соединенных Штатов Америки . 104 (10): 3901–6. дои : 10.1073/pnas.0611691104 . ПМК 1805696 . ПМИД 17360450 .
- ^ Гейзер Д.М., Клих М.А., Фрисвад Дж.К., Петерсон С.В., Варга Дж., Самсон Р.А. (2007). «Современное состояние распознавания и идентификации видов Aspergillus » . Исследования по микологии . 59 : 1–10. дои : 10.3114/sim.2007.59.01 . ПМК 2275194 . ПМИД 18490947 .
- ^ Jump up to: а б Гилмор С.Р., Грефенхан Т., Луи-Сейз Дж., Зайферт К.А. (май 2009 г.). «Множественные копии цитохромоксидазы 1 у видов грибов рода Fusarium » . Ресурсы молекулярной экологии . 9 Приложение с1 (Приложение 1): 90–8. дои : 10.1111/j.1755-0998.2009.02636.x . ПМИД 21564969 .
- ^ Виалле А., Фео Н., Аллер М., Дидух М., Мартин Ф., Монкальво Дж.М., Хамельн Р.К. (май 2009 г.). «Оценка митохондриальных генов как штрих-кода ДНК базидиомикоты» . Ресурсы молекулярной экологии . 9 Приложение с1 (Приложение 1): 99–113. дои : 10.1111/j.1755-0998.2009.02637.x . ПМИД 21564970 .
- ^ Jump up to: а б Дентингер Б.Т., Дидух М.Ю., Монкальво Дж.М. (2011). «Сравнение COI и ITS как маркеров штрих-кода ДНК грибов и их союзников (Agaricomycotina)» . ПЛОС ОДИН . 6 (9): e25081. Бибкод : 2011PLoSO...625081D . дои : 10.1371/journal.pone.0025081 . ПМК 3178597 . ПМИД 21966418 .
- ^ Ферандон С., Муха С., Каллак П., Бенедетто Х.П., Кастровьехо М., Баррозу Г. (ноябрь 2010 г.). « Ген Agaricus bisporus cox1: самый длинный митохондриальный ген и самый большой резервуар интронов митохондриальной группы I» . ПЛОС ОДИН . 5 (11): e14048. Бибкод : 2010PLoSO...514048F . дои : 10.1371/journal.pone.0014048 . ПМЦ 2987802 . ПМИД 21124976 .
- ^ Йенсен-Варгас Э., Марицци С. (июнь 2018 г.). «Штрих-кодирование ДНК для идентификации потребительских грибов, продаваемых в Нью-Йорке: мощный инструмент для гражданских ученых?» . Еда . 7 (6): 87. doi : 10.3390/foods7060087 . ПМК 6025134 . ПМИД 29890621 .
- ^ Йенсен-Варгас Э., Абреу А. Штрих-кодирование ДНК для идентификации потребительских грибов, продаваемых в Нью-Йорке (PDF) (Отчет). Архивировано (PDF) из оригинала 3 мая 2022 года . Проверено 4 мая 2020 г.
- ^ Раджа Х.А., Бейкер Т.Р., Литтл Дж.Г., Оберлис, Нью-Хэмпшир (январь 2017 г.). «ДНК-штрих-кодирование для идентификации потребительских грибов: частичное решение для сертификации продукции?» (PDF) . Пищевая химия . 214 : 383–392. doi : 10.1016/j.foodchem.2016.07.052 . ПМИД 27507489 . Архивировано (PDF) из оригинала 18 ноября 2023 года . Проверено 6 июня 2024 г.
- ^ Сян Л., Сун Дж., Синь Т., Чжу Ю., Ши Л., Сюй Икс и др. (октябрь 2013 г.). «ДНК-штрих-кодирование коммерческого китайского гусеничного гриба» . Письма FEMS по микробиологии . 347 (2): 156–62. дои : 10.1111/1574-6968.12233 . ПМИД 23927075 .
- ^ Кхуннамвонг П., Лертваттанасакул Н., Джиндаморакот С., Лимтонг С., Лашанс М.А. (декабрь 2015 г.). «Описание Diutina gen. nov., Diutina siamensis , fa sp. nov. и отнесение Candida catenulata , Candida mesorugosa ( Candida neorugosa , , Candida pseudorugosa , Candida ranongensis , Candida Rugosa и Candida scorzettiae к роду Diutina » PDF) . Международный журнал систематической и эволюционной микробиологии . 65 (12): 4701–9. дои : 10.1099/ijsem.0.000634 . ПМИД 26410375 . Архивировано (PDF) из оригинала 6 июня. Получено 5 мая.
- ^ Таванти А., Дэвидсон А.Д., Гоу Н.А., Мейден МС, Оддс ФК (январь 2005 г.). « Candida ortopsilosis и Candida Metapsilosis spp. Ноябрь для замены Candida parapsilosis групп II и III» . Журнал клинической микробиологии . 43 (1): 284–92. doi : 10.1128/JCM.43.1.284-292.2005 . ПМК 540126 . ПМИД 15634984 .
- ^ Хоанг М.Т., Ирини Л., Чен С.С., Соррелл Т.К., Мейер В. (2019). «Двойное ДНК-штрихкодирование для молекулярной идентификации возбудителей инвазивных грибковых инфекций» . Границы микробиологии . 10 (1647): 1647. doi : 10.3389/fmicb.2019.01647 . ПМК 6657352 . ПМИД 31379792 .
- ^ Секейра СО, ХП С, Мескита НУ, Португалия АН, Маседо МФ (2019). «Грибковые пятна на бумаге: то, что вы видите, то и получаете?». (PDF) . Сохранить наследие . 32 :18–27. дои : 10.14568/cp2018007 . Архивировано (PDF) из оригинала 15 марта 2020 г. Проверено 17 апреля 2020 г.
- ^ Трован Х, Португалия А, Соареш Ф, Пайва ДС, Мескита Н, Коэлью К, Пиньейру АС, Катарино Л, Жил Ф, Тьягу И (2019). «Грибковое разнообразие и распространение через отчетливое явление биологического разрушения в известняковых стенах старого катетера Коимбры, объекта Всемирного наследия ЮНЕСКО». Международная биопорча и биодеградация . 142 : 91–102. Бибкод : 2019IBiBi.142...91T . дои : 10.1016/j.ibiod.2019.05.008 . S2CID 182913598 .
- ^ Пайва де Карвалью Х, Оливейра Секейра С, Пиньо Д, Трован Х, Фернандеш да Кошта РМ, Эгаш К, Маседу МФ, Португалия А (2019). «Сочетание инновационного неинвазивного метода отбора проб и высокопроизводительного секвенирования для характеристики грибковых сообществ на холсте». Международная биопорча и биодеградация . 145 : 104816. Бибкод : 2019IBiBi.14504816P . дои : 10.1016/j.ibiod.2019.104816 . S2CID 208554023 .
Дальнейшее чтение
[ редактировать ]- Эберхардт Ю (июль 2010 г.). «Конструктивный шаг к выбору штрих-кода ДНК грибов» . Новый фитолог . 187 (2): 265–8. дои : 10.1111/j.1469-8137.2010.03329.x . ПМИД 20642723 .
Внешние ссылки
[ редактировать ]- Список праймеров для афтола (используемый в шестигенной филогении Джеймса и др., 2006 г.)