Вирус клекаемого энцефалита
| Вирус клекаемого энцефалита | |
|---|---|
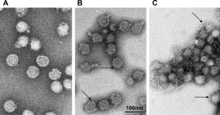
| |
| TBEV на разных уровнях pH | |
| Классификация вирусов | |
| (не вмешательство): | Вирус |
| Область : | Рибовирия |
| Королевство: | Orthornavirae |
| Филум: | Kitrinoviricota |
| Сорт: | Flasuviricetes |
| Заказ: | Желтовирал |
| Семья: | Flaviviridae |
| Род: | Флавивирус |
| Разновидность: | Вирус клекаемого энцефалита
|
| Штаммы | |
| |
Вирус энцефалита, подаваемый клелем ( TBEV ), представляет собой вирус РНК с положительной цепью, связанный с клещевым энцефалитом , в роду Flavivirus .
Классификация
[ редактировать ]Таксономия
[ редактировать ]TBEV является членом рода Flavivirus . Другие близкие родственники, члены серокомплекса TBEV, включают вируса геморрагической лихорадки OMSK , вирус заболевания леса Кьясанур , вирус Альхурма , вирус с пустым и вирусом Лангата . [ 1 ]
Подтипы
[ редактировать ]TBEV имеет три подтипа:
- Западный европейский подтип (ранее вирус центрального европейского энцефалита, CEEV; основной вектор клещей: Ixodes Ricinus );
- Сибирский подтип (ранее Западный Сибирский вирус; основной вектор клещей: Ixodes persulcatus );
- Дальневосточный подтип (ранее российский летний вирус энцефалита, RSSEV; главный вектор клещей: Ixodes persulcatus ). [ 2 ]
Опорно -деформацией является штамм Софджина. [ 3 ]
Вирусология
[ редактировать ]Структура
[ редактировать ]TBEV представляет собой с положительным смыслом одноцепочечный РНК-вирус , содержащийся в сферической капсиде 40-60 нм . [ 1 ] Геном TBEV составляет приблизительно 11 КБ размером, который содержит 5 'крышку , единственную рамку открытого считывания с 3' и 5 ' UTRS , и без полиаденилирования . [ 1 ] Как другие флавивирусы, [ 4 ] Геном TBEV кодирует для десяти вирусных белков, трех структурных и семи Нтруктурных . [ 5 ] [ 1 ]
Структурными белками являются C ( капсид ), PRM (Premembrane), который расщепляется для получения конечного мембранного белка , (M) и белка оболочки (E). Семь неструктурных белков: NS1, NS2A, NS2B, NS3, NS4A, NS4B и NS5. Роль некоторых неструктурных белков известна, NS5 служит РНК-зависимой РНК-полимеразой , NS3 обладает протеазой (в комплексе с NS2B) и геликазной активностью. [ 5 ] [ 1 ] Структурные и неструктурные белки не требуются для инфекционного генома. [ 1 ] Все вирусные белки экспрессируются в виде единого большого полипротеина, с порядком C, PRM, E, NS1, NS2A, NS2B, NS3, NS4A, NS4B, NS5. [ 5 ]
Вирусные генетические детерминанты патогенности
[ редактировать ]Белок оболочки участвует в рецепторном связывании и нейровирулентности, где повышенная гликозаминогликан-связывающая сродство ослабляет нейроинвазивность. [ 6 ] На конформацию белка E во время секреции вирусных частиц также влияет гликозилирование. [ 7 ] Была продемонстрирована иммуногенность TBEV NS1, демонстрируя его способность вызывать окислительный стресс и выявлять экспрессию иммунопротеатических субъединиц. Кроме того, было обнаружено, что он стимулирует выработку цитокинов. [ 8 ] Белок NS5 обладает активностью интерферонов антагоновой активности, поскольку он подавляет экспрессию субъединицы рецептора IFN. Не структурный белок 5 (NS5) влияет на невропатогенез путем ослабления исхода нейритов. Нетрансляционная область 3 (UTR3) и UTR 5 влияют на циклизацию и репликацию геномной РНК, а также транспорт вирусной РНК в дендритах, что влияет на нейрогенез и синаптическую связь. [ 6 ]
Жизненный цикл
[ редактировать ]Передача инфекции
[ редактировать ]Инфекция вектора начинается , когда клеща получает кровь от зараженного хозяина . Это может произойти в любой части жизненного цикла клеща, но горизонтальная передача между инфицированными нимфами и неинфицированными личинками , совместно с тем, что он на том же хозяине является ключевым для поддержания циркуляции TBEV. [ 9 ] [ 1 ] TBEV в крови хозяина заражает клеща через среднюю кишку , откуда он может передать слюнные железы, которые будут переданы следующему хозяину. В клещах, не являющихся адультами, TBEV передается трансстадиально заражая клеток, которые не разрушаются во время лишения , поэтому клеща остается инфекционным на протяжении всей своей жизни. [ 9 ] Зараженные взрослые клещи могут быть в состоянии откладывать яйца, которые заражены, передавая вирус транвариально . [ 10 ]
Репликация
[ редактировать ]
У людей инфекция начинается в коже (за исключением случаев пищевых продуктов, около 1% инфекций) в месте укуса инфицированного клеща, где клетки Лангерганса и макрофаги в коже предпочтительны. [ 5 ] Белки TBEV Envelope (E) распознают гепарансульфат (и, вероятно, другие рецепторы) на поверхности клетки -хозяина и эндоцитозируются через клатрин -опосредованный путь . Подкисление поздней эндосомы запускает конформационное изменение белков E, что приводит к слиянию, с последующим непосредственным и высвобождением одноцепочечного генома РНК в цитоплазму. [ 11 ] [ 1 ]
Вирусный полипротеин транслируется и вставляется в мембрану ER , где он обрабатывается на цитозольной стороне пептидазами -хозяевами и в просвете при действии вирусного фермента. Вирусные белки C, NS3 и NS5 расщепляются в цитозоль (хотя NS3 может комплексный с NS2B или NS4A для выполнения протеолитической или геликазной активности), в то время как оставшиеся неструктурные белки изменяют структуру мембраны ER. Эта измененная мембрана позволяет сборке репликационных комплексов, где вирусный геном реплицируется вирусной РНК-зависимой РНК-полимеразой , NS5. [ 11 ] [ 5 ]
Вновь реплицируемые геномы вирусной РНК затем упаковываются белками C, в то время как на цитозольной стороне мембраны ER образуют незрелый нуклеокапсид и получают белки E и PRM, расположенные как гетеродимер, во время подачи почки в просвет ER. Незрелый вирион колючий и геометрический по сравнению со зрелой частицей. Частица проходит через аппарат Гольджи и транс -Гольджи, при все более кислых условиях, с помощью которых вирион созревает с расщеплением сегмента PR из M -белка и гомодимеров Fusion Fusion Fusion. Хотя расщепленный пиар -сегмент остается связанным с белковым комплексом до выхода. [ 1 ] [ 11 ]
Вирус высвобождается из клетки -хозяина после слияния транспортной пузырьки с клеточной мембраной хозяина, сегменты расщепленного PR теперь диссоциации, что приводит к полностью зрелому инфекционному вирусу. [ 1 ] [ 11 ] Тем не менее, частично зрелые и незрелые вирусы иногда также высвобождаются; Незрелые вирусы неинфекционны, так как белки E не являются компетентными слияния, частично зрелые вирусы все еще способны к инфекции. [ 11 ]
Патогенез и иммунный ответ
[ редактировать ]
За исключением случаев пищевых продуктов, инфекция начинается в коже на месте укуса клеща. Дендритные . (или Лангерганс) клетки (DC) являются предпочтительно целенаправленными [ 5 ] Первоначально вирус повторяет локально и иммунный ответ, запускается, когда вирусные компоненты распознаются рецепторами распознавания цитозольных картин (PRR), такими как рецепторы, похожие на Toll (TLR). [ 12 ] Распознавание вызывает высвобождение цитокинов, включая интерфероны (IFN) α, β и γ и хемокины, притягивая миграционные иммунные клетки в место укуса. [ 5 ] Инфекция может быть остановлена на этом этапе и очищена до начала заметных симптомов. Примечательно, что слюна клещей усиливает инфекцию за счет модуляции иммунного ответа хозяина, ослабляя апоптотические сигналы. [ 12 ] Если инфекция продолжается, мигрирующие DC и макрофаги заражаются и перемещаются в локальный дренирующий лимфатический узел, где активация полиморфноядерных лейкоцитов , моноцитов и системы комплемента . активируется [ 12 ]
Дренирующий лимфатический узел также может служить в качестве вирусного амплификационного сайта, откуда TBEV получает системный доступ. Эта стадия Viremic соответствует первой симптоматической фазе в прототипной двухфазной паттерне энцефалита, подаваемого клетками. [ 1 ] TBEV имеет сильное предпочтение нейрональной ткани и является нейроинвазивным. [ 13 ] Начальная стадия вируса обеспечивает доступ к ряду преференциальных тканей. Однако точный механизм, с помощью которого TBEV пересекается в центральную нервную систему (CNS), неясен. [ 13 ] [ 12 ] [ 1 ] Существует несколько предлагаемых механизма для нарушения гематоэнцефалического барьеры (BBB): 1) механизм «троянской лошади», посредством которого TBEV получает доступ к ЦНС при заражении иммунной клетки, которая проходит через BBB; [ 12 ] [ 5 ] [ 13 ] 2) нарушение и повышенная проницаемость BBB с помощью иммунных иммунных цитокинов; [ 13 ] 3) через инфекцию обонятельных нейронов; [ 5 ] 4) через ретроградный транспорт вдоль периферических нервов к ЦНС; [ 5 ] 5) Инфекция клеток, которые составляют часть BBB. [ 5 ] [ 12 ]
Инфекция ЦНС приводит на второй этап классической двухфазной модели инфекции, связанной с европейским подтипом. Болезнь ЦНС является иммунопатологической; высвобождение воспалительных цитокинов в сочетании с действием цитотоксических CD8+ Т -клеток и, возможно, NK -клеток приводит к воспалению и апоптозу инфицированных клеток, которые ответственны за многие симптомы CNS. [ 12 ] [ 13 ]
Гуморальный ответ
[ редактировать ]Специфические антитела IgM и IgG производятся в ответ на инфекцию. [ 1 ] Сначала появляются антитела IgM, а также достигают более высоких уровней, и, как правило, через 1,5 месяца после заражения, хотя существует значительное изменение от пациента к пациенту. Уровни IgG достиг пика примерно через 6 недель после появления симптомов ЦНС, затем немного снижаются, но не рассеиваются, вероятно, придавая иммунитету на длине жизни пациенту. [ 1 ] [ 5 ]
Эволюция
[ редактировать ]Предок существующих штаммов, по -видимому, разделился на несколько клад примерно 2750 лет назад. [ 14 ] Сибирские дальневосточные и подтипы разошлись около 2250 лет назад . Второй анализ предполагает более раннюю дату эволюции (3300 лет назад) с быстрым увеличением числа штаммов, начиная с 300 лет назад. [ 15 ] Различные штаммы вируса были переданы не менее трех раз в Японию между 260–430 лет назад. [ 16 ] [ 17 ] Штаммы, циркулирующие в Латвии , по -видимому, возникли как из России, так и из Западной Европы [ 18 ] в то время как те в Эстонии, похоже, возникли в России . [ 19 ] Литовские штаммы, по -видимому , связаны с таковыми из Западной Европы. [ 20 ] Филогенетический анализ указывает на то, что европейские и сибирские подтипы TBEV тесно связаны, в то время как в дальнем восточном подтипе ближе к нелющному ищному вирусу. [ 1 ] Однако в отношении антигенной связанности, основанного на белках E, NS3 и NS5, все три подтипа очень похожи, а яростный вирус-это ближайший относительный от коллективной группы TBEV. [ 21 ]
История
[ редактировать ]Хотя первое описание того, что, возможно, появилось в записях в 1700 -х годах в Скандинавии, [ 13 ] Идентификация вируса TBEV произошла в Советском Союзе в 1930 -х годах. [ 22 ] Расследование началось из -за вспышки того, что считалось японским энцефалитом («летний энцефалит»), среди советских войск, размещенных вдоль границы с японской империей (сегодняшняя Народная Республика ), недалеко от дальневосточного города Хабаровский . Экспедицию возглавляли виролог Лев А. Зилбер , который собрал команду из двадцати молодых ученых в ряде связанных областей, таких как акарология , микробиология, неврология и эпидемиология. [ 23 ] [ 22 ] Экспедиция прибыла в Хабаровск 15 мая 1937 года и разделена на отряды, под руководящими северами Элизабет Н. Левкович и работая в Хабаровском Краи , и под руководством Александры Д. Шебольдаева, работая в Примарском Край . [ 22 ]
В течение месяца мая экспедиция идентифицировала клещи как вектор, собранные I. persucatus , воздействие на голой кожи энтомологом Александра В. Гуцевича и вирусолога Михаила П. Чумакова выделяли вирус из клещей, кормивших намеренно зараженных мышей. В течение лета пять членов экспедиций заразились TBEV, и, хотя не было смертельных случаев, три из пяти пострадали от повреждений . [ 22 ]
Экспедиция вернулась в середине августа, а в октябре 1937 года Зилбер и Шеболдова были арестованы, ложно обвиненные в распространении японского энцефалита. Эпидемиолог экспедиции Тамара М. Сапонова была арестована в следующем январе за протест против обвинений против Зилбера и Шеболдовы. Как следствие арестов, одна из важных первоначальных работ была опубликована под авторством экспедиционного акаролога Васили С. Миронов. Зилбер был выпущен в 1939 году и сумел восстановить вместе с Шеболдовой, соавторство этой первоначальной работы; Тем не менее, Сафанов и Шеболдова (которые не были освобождены) провели 18 лет в трудовых лагерях . [ 22 ] [ 23 ]
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а Мэнсфилд К.Л., Джонсон Н., Фиппс Л.П., Стивенсон -младший, Фукс А.Р., Соломон Т (август 2009 г.). «Вирус клекаемого энцефалита - обзор появляющегося зооноза» . Журнал общей вирусологии . 90 (Pt 8): 1781–1794. doi : 10.1099/vir.0.011437-0 . PMID 19420159 .
- ^ Гудман Дж.Л., Деннис Д.Т., Соненшин Д.Е. (2005). «Галочный энцефалит». Глемовые заболевания людей . Вашингтон, округ Колумбия: ASM Press. п. 151. ISBN 978-1-55581-238-6 .
- ^ Kovalev SY, Mukhacheva TA, Kokorev VS, Belyaeva IV (апрель 2012 г.). «Вирус клеща-энцефалита: контрольный штамм софджин и проблема его подлинности». Вирусные гены . 44 (2): 217–224. doi : 10.1007/s11262-011-0690-9 . PMID 22095094 . S2CID 12587373 .
- ^ Уайлдер-Смит А., Оой Э.Е., Хорстик О, Уиллс Б (январь 2019 г.). "Денге". Лансет . 393 (10169): 350–363. doi : 10.1016/s0140-6736 (18) 32560-1 . PMID 30696575 . S2CID 208789595 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л Рузек Д., Авсич Жупанкант Т., Борде Дж., Крдл А., Эйер Л., Каргана Г. и др. (Апрель 2019). «Глемовой энцефалит в Европе и России: обзор патогенеза, клинических особенностей, терапии и вакцин». Антивирусные исследования . 164 : 23–51. doi : 10.1016/j.antiviral.2019.01.014 . PMID 30710567 . S2CID 73414822 .
- ^ Jump up to: а беременный Йоший К (март 2019 г.). «Эпидемиология и патологические механизмы клещевого энцефалита» . Журнал ветеринарной медицинской науки . 81 (3): 343–347. doi : 10.1292/jvms.18-0373 . PMC 6451894 . PMID 30674746 .
- ^ Йоший К., Янагихара Н., Ишизука М., Сакай М., Карива Х (октябрь 2013 г.). «N-связанный гликан в вирусе вируса вируса энцефалита, подаваемого клеком, влияет на вирусную секрецию в клетках млекопитающих, но не в клетках» . Журнал общей вирусологии . 94 (Pt 10): 2249–2258. doi : 10.1099/vir.0.055269-0 . HDL : 2115/56524 . PMID 23824303 .
- ^ Starodubova E, Tuchynkaya K, Kuzmenko Y, Latanova A, Tutyaeva V, Karpov V, Karganova G (январь 2023 г.). «Активация ранних провоспалительных ответов TBEV NS1 варьируется между штаммами различных подтипов» . Международный журнал молекулярных наук . 24 (2): 1011. doi : 10.3390/ijms24021011 . PMC 9863113 . PMID 36674524 .
- ^ Jump up to: а беременный Nuttall PA, Jones LD, Labuda M, Kaufman WR (январь 1994 г.). «Адаптация арбовирусов к клещам». Журнал медицинской энтомологии . 31 (1): 1–9. doi : 10.7939/r3m03xx77 . PMID 8158611 .
- ^ Danielová V, Holubová J, Pejcoch M, Daniel M (2002). «Потенциальная значимость трансвариальной передачи в циркуляции вируса клеща-барне энцефалита» . Folia Parasitologica . 49 (4): 323–325. Doi : 10.14411/fp.2002.060 . PMID 12641208 .
- ^ Jump up to: а беременный в дюймовый и Пулккинен Ли, Мясник С.Дж., Анастасина М (июнь 2018 г.). «Вирус клекаемого энцефалита: структурный взгляд» . Вирусы . 10 (7): 350. doi : 10.3390/v10070350 . PMC 6071267 . PMID 29958443 .
- ^ Jump up to: а беременный в дюймовый и фон глин Velay A, Paz M, Cesbron M, Gantner P, Solis M, Sulier E, et al. (Август 2019). «Вирус клеща-энцефалита: молекулярные детерминанты невропатогенеза появляющегося патогена». Критические обзоры в микробиологии . 45 (4): 472–493. doi : 10.1080/1040841x.2019.1629872 . PMID 31267816 . S2CID 195787988 .
- ^ Jump up to: а беременный в дюймовый и фон Blom K, Cuapio A, Sandberg JT, Varnaite R, Michaëlsson J, Björkström NK, et al. (2018). «Клеточно-опосредованные иммунные ответы и иммунопатогенез вируса вируса энцефалита человека» человека » . Границы в иммунологии . 9 : 2174. DOI : 10.3389/fimmu.2018.02174 . PMC 6168641 . PMID 30319632 .
- ^ Subbotina EL, Loktev VB (2012). «[Молекулярная эволюция энцефалита и вирусов Powassan, подаваемого клетками]». Molekuliarnaia Biologiia . 46 (1): 82–92. doi : 10.1134/s0026893311060148 . PMID 22642104 . S2CID 18500235 .
- ^ Uzcátegui NY, Sironen T, Golovljova I, Jääskeläinen Ae, Välimaa H, Lundkvist Å, et al. (Апрель 2012 г.). «Скорость эволюции и молекулярной эпидемиологии вируса клещей энцефалита в Европе, включая две изоляции из одного и того же фокуса с интервалом» . Журнал общей вирусологии . 93 (Pt 4): 786–796. doi : 10.1099/vir.0.035766-0 . PMID 22205716 .
- ^ Suzuki Y (июнь 2007 г.). «Многочисленные трансмиссии вируса клещей энцефалита между Японией и Россией» . Гены и генетические системы . 82 (3): 187–195. doi : 10.1266/ggs.82.187 . PMID 17660689 .
- ^ Такашима I, Хаясака Д., Гото А., Карива Х, Мизутани Т (февраль 2001 г.). «Эпидемиология клещевого энцефалита (TBE) и филогенетический анализ вирусов TBE в Японии и на крайней России». Японский журнал инфекционных заболеваний . 54 (1): 1–11. PMID 11326122 .
- ^ Vene S, Golovljova I, Mavtchoutko V, Forsgren M, Kalnina V, Plyusnin A (декабрь 2001 г.). «Характеристика вируса клещей энцефалита из Латвии: свидетельство совместной циркуляции трех различных подтипов». Журнал медицинской вирусологии . 65 (4): 730–735. doi : 10.1002/jmv.2097 . PMID 11745938 . S2CID 22860154 .
- ^ Golovljova I, Vene S, Sjölander KB, Vasilenko V, Plyusnin A, Lundkvist A (декабрь 2004 г.). «Характеристика вируса энцефалита, передаваемого клетками из Эстонии». Журнал медицинской вирусологии . 74 (4): 580–588. doi : 10.1002/jmv.20224 . PMID 15484275 . S2CID 28491834 .
- ^ Микиен А., Вен С., Головльжова И., Лайсконис А., Линдквист Л., Плиуснин А., Лундквист А (декабрь 2001 г.). «Вирус клекаемого энцефалита в Литве». Европейский журнал клинической микробиологии и инфекционных заболеваний . 20 (12): 886–888. doi : 10.1007/s10096-001-0637-5 . PMID 11837641 . S2CID 38061544 .
- ^ Heinz FX, Stiasny K (2019). «Глава 2B: молекулярная и антигенная структура TBEV» . Телечный энцефалит - книга . doi : 10.33442/978-981-14-0914-1_2b . ISSN 2661-3980 . S2CID 150118109 .
- ^ Jump up to: а беременный в дюймовый и Zlobin VI, Pogodina VV, Kahl O (октябрь 2017). «Краткая история открытия вируса энцефалита, передаваемого клетками в конце 1930-х годов (на основе воспоминаний о членах экспедиций, их коллег и родственников)». Клеща и заболевания клещей . 8 (6): 813–820. doi : 10.1016/j.ttbdis.2017.05.001 . PMID 28526419 .
- ^ Jump up to: а беременный USPENSKY I (май 2018). «Несколько слов в дополнение к« краткой истории открытия вируса энцефалита клеща в конце 1930-х годов »Vi Zlobin, VV Pogodina и O. Kahl (Ttbdis, 2017, 8, 813-820)». Клеща и заболевания клещей . 9 (4): 834–835. doi : 10.1016/j.ttbdis.2018.03.007 . PMID 29559213 .
Внешние ссылки
[ редактировать ]- Энцефалит+вирусы,+клеща в Национальной медицинской библиотеке Медицинской библиотеки США (Mesh)
- Телевой энцефалит во Всемирной организации здравоохранения
- Книга 4 -го издания, Герхард Доблер, Вильгельм Эрбер, Майкл Брёкер, Хайнц -Джосеф Шмитт, Global Health Press, 25 мая 2021 г. -pp 386pp
| Базы данных управления авторитетом : национальный |
|---|