Развивающее биоэлектричество
Данная статья может содержать чрезмерное количество цитат . ( Ноябрь 2023 г. ) |

Биоэлектричество развития — это регуляция формирования и поведения на уровне клеток , тканей и органов с помощью электрических сигналов во время развития эмбриональных животных и растений . Носителем заряда в развивающемся биоэлектричестве является ион (заряженный атом), а не электрон , и электрический ток и поле генерируются всякий раз, когда возникает чистый поток ионов. Клетки и ткани всех типов используют потоки ионов для электрической связи. Эндогенные электрические токи и поля , потоки ионов и различия в потенциале покоя в тканях составляют сигнальную систему. Он функционирует вместе с биохимическими факторами, транскрипционными сетями и другими физическими силами, регулируя поведение клеток и крупномасштабное формирование паттернов в таких процессах, как эмбриогенез , регенерация и подавление рака .
Обзор
[ редактировать ]Биоэлектричество развития — это раздел биологии, связанный с нейрофизиологией и биоэлектромагнетизмом , но отличающийся от них . Биоэлектричество развития относится к потокам эндогенных ионов, трансмембранным и трансэпителиальным градиентам напряжения, а также электрическим токам и полям, создаваемым и поддерживаемым в живых клетках и тканях. [2] [3] Эта электрическая активность часто используется во время эмбриогенеза, регенерации и подавления рака — это один слой сложного поля сигналов, которые воздействуют на все клетки in vivo и регулируют их взаимодействия во время формирования и поддержания структуры. Это отличается от нейронного биоэлектричества (классически называемого электрофизиологией), которое относится к быстрым и временным выбросам в хорошо известных возбудимых клетках, таких как нейроны и миоциты (мышечные клетки); [4] и от биоэлектромагнетизма, который относится к эффектам приложенного электромагнитного излучения, и эндогенного электромагнетизма, такого как биофотонная эмиссия и магнетит. [5] [6]


Разрыв внутри и снаружи на поверхности клетки, обеспечиваемый липидной двухслойной мембраной (конденсатором), лежит в основе биоэлектричества. Плазматическая мембрана была незаменимой структурой для возникновения и эволюции самой жизни. Это обеспечило разделение, позволяющее установить дифференциальное напряжение/градиент потенциала (батарея или источник напряжения) через мембрану , что, вероятно, позволило создать раннюю и рудиментарную биоэнергетику, которая питала клеточные механизмы. [9] [10] В ходе эволюции первоначально чисто пассивная диффузия ионов (носителей заряда) постепенно стала контролироваться появлением ионных каналов , насосов , обменников и переносчиков. Эти энергетически свободные (резисторы или проводники, пассивный транспорт) или дорогие (источники тока, активный транспорт) транслокаторы устанавливают и точно настраивают градиенты напряжения – потенциалы покоя – которые повсеместно распространены и необходимы для физиологии жизни, начиная от биоэнергетики, движения, восприятия и транспорта питательных веществ. , выведение токсинов и передача сигналов в гомеостатических состояниях, а также при заболеваниях/травмах. При раздражении или нарушении барьера (коротком замыкании) мембраны ионы, питаемые градиентом напряжения (электродвижущей силой), диффундируют или просачиваются соответственно через цитоплазму и интерстициальную жидкость (проводники), генерируя измеримые электрические токи – чистые потоки ионов – и поля. Некоторые ионы (например, кальций ) и молекулы (например, перекись водорода ) модулируют целевые транслокаторы, создавая ток или усиливая, смягчая или даже обращая вспять первоначальный ток, являясь переключателями. [11] [12]
Эндогенные биоэлектрические сигналы производятся в клетках в результате совокупного действия ионных каналов, насосов и транспортеров. В невозбудимых клетках потенциал покоя через плазматическую мембрану (Vmem) отдельных клеток распространяется на расстояния через электрические синапсы, известные как щелевые контакты (проводники), которые позволяют клеткам делиться своим потенциалом покоя с соседями. Выровненные и сложенные друг на друга клетки (например, в эпителии) генерируют трансэпителиальные потенциалы (например, последовательно соединенные батареи) и электрические поля, которые аналогичным образом распространяются по тканям. [13] Плотные контакты (резисторы) эффективно уменьшают параклеточную диффузию и утечку ионов, предотвращая короткое замыкание по напряжению. Вместе эти напряжения и электрические поля образуют богатые и динамичные узоры внутри живых тел, которые разграничивают анатомические особенности , действуя таким образом в некоторых случаях как чертежи экспрессии генов и морфогенеза. Эти биоэлектрические распределения являются не просто корреляциями, они динамичны, развиваются со временем, в зависимости от микроокружения и даже условий на большом расстоянии, оказывая поучительное влияние на поведение клеток и крупномасштабное формирование паттернов во время эмбриогенеза, регенерации и подавления рака. [3] [14] [8] [15] [16] Механизмы биоэлектрического контроля являются важной новой целью достижений в регенеративной медицине, врожденных дефектах , раке и синтетической биоинженерии . [17] [18]
История
[ редактировать ]18 век
[ редактировать ]Развитие биоэлектричества началось в 18 веке. Несколько плодотворных работ, стимулирующих мышечные сокращения с помощью лейденских банок, завершились публикацией классических исследований Луиджи Гальвани в 1791 и 1794 годах (De viribus electricitatis in motu Musclei). В них Гальвани считал, что обнаружил внутреннюю способность производить электричество в живых тканях или " животное электричество». Алессандро Вольта показал, что подергивание мышц лапки лягушки было вызвано генератором статического электричества и разнородными металлами, подвергающимися или катализирующими электрохимические реакции . Гальвани в исследовании 1794 года продемонстрировал подергивания без металлического электричества путем прикосновения к мышце ноги отклоняющимся перерезом седалищного нерва , окончательно демонстрируя «животное электричество». [19] [20] [21] Сам того не зная, Гальвани в этом и связанных с ним экспериментах обнаружил ток повреждения (утечка ионов, вызванная потенциалом неповрежденной мембраны/эпителия) и потенциал повреждения (разница потенциалов между поврежденной и неповрежденной мембраной/эпителием). Потенциал травмы на самом деле был электрическим источником сокращения ног, как это стало понятно в следующем столетии. [22] [23] Последующие работы в конечном итоге расширили эту область за пределы нервных и мышечных клеток на все клетки, от бактерий до невозбудимых клеток млекопитающих.
19 век
[ редактировать ]Основываясь на более ранних исследованиях, дальнейшие проблески развивающегося биоэлектричества произошли с открытием электрических токов и полей, связанных с ранами, в 1840-х годах, когда электрофизиолог Эмиль дю Буа-Реймон сообщил о электрической активности макроскопического уровня в телах лягушек, рыб и людей. Он регистрировал мельчайшие электрические токи в живых тканях и организмах с помощью современного на тот момент гальванометра, сделанного из изолированных катушек медной проволоки. Он открыл быстро меняющееся электричество, связанное с мышечными сокращениями и возбуждением нервов – потенциалы действия . [24] [25] [26] Дюбуа-Реймон также подробно сообщил о менее колеблющемся электричестве при ранах (ток и потенциал травмы), которые он нанес себе. [27] [28]

Начало 20 века
[ редактировать ]Работы по развитию биоэлектричества начались всерьез в начале 20-го века. [30] Ида Х. Хайд изучала роль электричества в развитии яиц. [31] Т.Х. Морган и другие изучали электрофизиологию дождевого червя. [32] Орен Э. Фрейзи изучал влияние электричества на регенерацию конечностей у амфибий. [33] Э. Дж. Лунд исследовал морфогенез цветковых растений. [34] Либби Хайман изучала позвоночных и беспозвоночных животных. [35] [36]
В 1920-х и 1930-х годах Элмер Дж. Лунд [37] и Гарольд Сакстон Берр [38] написал множество статей о роли электричества в эмбриональном развитии. [29] Лунд измерил токи в большом количестве живых модельных систем, сопоставив их с изменениями в структуре структур. Напротив, Берр использовал вольтметр для измерения градиентов напряжения, исследуя развивающиеся эмбриональные ткани и опухоли у ряда животных и растений. продемонстрировали, что приложенные электрические поля изменяют регенерацию планарий в 1940-х и 1950-х годах. Марш и Бимс [39] [40] вызывая образование голов или хвостов в местах разрезов, меняя основную полярность тела.
Конец 20 века
[ редактировать ]В 1970-х годах внедрение и разработка Лайонелом Яффе и Ричардом Нучиттелли вибрационного зонда, первого устройства для количественной неинвазивной характеристики внеклеточных мельчайших ионных токов, оживили эту область. [41] [42] [43] [44] [45]
Такие исследователи, как Джозеф Ванейбл, Ричард Боргенс, Кен Робинсон и Колин Маккейг, исследовали роль эндогенной биоэлектрической передачи сигналов в развитии и регенерации конечностей, эмбриогенезе, полярности органов и заживлении ран . [46] [47] [23] [48]
CD Cone изучал роль потенциала покоя в регуляции дифференцировки и пролиферации клеток . [49] [50] Последующая работа выявила определенные области спектра потенциалов покоя, которые соответствуют различным состояниям клеток, таким как покой, ствол, рак и терминальная дифференцировка. [51]
Хотя этот объем работ позволил получить значительный объем высококачественных физиологических данных, этот крупномасштабный биофизический подход исторически занимает второе место после изучения биохимических градиентов и генетических сетей в биологическом образовании, финансировании и общей популярности среди биологов. Ключевым фактором, который способствовал отставанию этой области от молекулярной генетики и биохимии, является то, что биоэлектричество по своей сути является живым явлением — его нельзя изучать на фиксированных образцах. Работа с биоэлектричеством более сложна, чем традиционные подходы к биологии развития, как методологически, так и концептуально, поскольку обычно требует весьма междисциплинарного подхода. [15]
Методы исследования
[ редактировать ]Электроды
[ редактировать ]Золотым стандартом методов количественного извлечения электрических измерений из живых образцов, начиная от уровня клетки и заканчивая организмом, являются стеклянный микроэлектрод (или микропипетка ), вибрирующий (или самореферентный) зонд напряжения и вибрирующий ионоселективный микроэлектрод . Первый по своей природе инвазивный, а два последних неинвазивные, но все они сверхчувствительны. [52] и быстродействующие датчики, широко используемые во множестве физиологических состояний в широко распространенных биологических моделях. [53] [54] [11] [55] [23] [ чрезмерное цитирование ]
Стеклянный микроэлектрод был разработан в 1940-х годах для изучения потенциала действия возбудимых клеток на основе плодотворной работы Ходжкина и Хаксли на гигантском аксоне кальмара . [56] [57] Это просто жидкий солевой мостик, соединяющий биологический образец с электродом, защищающий ткани от вымываемых токсинов и окислительно-восстановительных реакций оголенного электрода. Благодаря низкому импедансу, низкому потенциалу перехода и слабой поляризации серебряные электроды являются стандартными преобразователями ионного тока в электрический, который происходит посредством обратимой окислительно-восстановительной реакции на поверхности электрода. [58]
Вибрирующий зонд был использован в биологических исследованиях в 1970-х годах. [59] [60] [41] Чувствительный к напряжению зонд покрыт платиной, образуя емкостный шарик с черным кончиком и большой площадью поверхности. При вибрации в искусственном или естественном градиенте постоянного напряжения емкостный шарик колеблется в синусоидальном выходном переменном токе. Амплитуда волны пропорциональна измеряемой разности потенциалов на частоте вибрации и эффективно фильтруется синхронным усилителем, повышающим чувствительность зонда. [41] [61] [62]
Вибрирующий ионоселективный микроэлектрод был впервые использован в 1990 году для измерения потоков кальция в различных клетках и тканях. [63] Ионоселективный микроэлектрод представляет собой адаптацию стеклянного микроэлектрода, в которой ионспецифический жидкий ионообменник (ионофор) заполняется на кончике предварительно силанизированного (для предотвращения утечки) микроэлектрода. Кроме того, микроэлектрод вибрирует на низких частотах, чтобы работать в режиме точной самореференции. Только определенный ион проникает через ионофор , поэтому показания напряжения пропорциональны концентрации ионов в условиях измерения. Затем поток рассчитывается по первому закону Фика . [61] [64]
Новые оптические методы, [65] например, оптрод pH (или оптрод ), который можно интегрировать в самореферентную систему, может стать альтернативным или дополнительным методом в лабораториях биоэлектричества. Оптрод не требует привязки и нечувствителен к электромагнетизму. [66] упрощая настройку системы и делая ее подходящим вариантом для записей, при которых одновременно применяется электрическая стимуляция.
В большой работе по функциональному изучению биоэлектрической передачи сигналов использовались прикладные (экзогенные) электрические токи и поля через устройства подачи напряжения постоянного и переменного тока, интегрированные с солевыми мостиками агарозы. [67] Эти устройства могут генерировать бесчисленные комбинации величины и направления напряжения, импульсов и частот. В настоящее время в этой области набирает популярность применение электрических полей с использованием технологии «лаборатория на чипе», что дает возможность проводить высокопроизводительные скрининговые анализы больших комбинаторных результатов. [68]

флуоресценция
[ редактировать ]Прогресс молекулярной биологии за последние шесть десятилетий позволил создать мощные инструменты, облегчающие анализ биохимических и генетических сигналов; тем не менее, они, как правило, не очень подходят для биоэлектрических исследований in vivo . Предыдущие работы в значительной степени полагались на ток, подаваемый непосредственно через электроды, чему способствовали недавние значительные достижения в области материаловедения. [70] [71] [72] [73] [74] [75] [ чрезмерное цитирование ] и измерения внеклеточного тока, чему способствуют сложные самореферентные электродные системы. [76] [77] Хотя применение электродов для управления процессами в организме, контролируемыми нейронами, в последнее время привлекло большое внимание. [78] [79] существуют и другие возможности управления соматическими процессами, поскольку большинство типов клеток электрически активны и реагируют на ионные сигналы, исходящие от себя и своих соседей.
В начале XXI века был разработан ряд новых молекулярных методов, которые позволили исследовать биоэлектрические пути с высокой степенью механистического разрешения и связать их с каноническими молекулярными каскадами. [80] К ним относятся:
- Фармакологический скрининг для выявления эндогенных каналов и насосов, ответственных за определенные события формирования паттерна; [81] [82] [83]
- Потенциал-чувствительные флуоресцентные репортерные красители и генетически кодируемые флуоресцентные индикаторы напряжения для характеристики биоэлектрического состояния in vivo. [84] [85] [86] [87] [88]
- Панели хорошо охарактеризованных доминантных ионных каналов, которые могут неправильно экспрессироваться в интересующих клетках, чтобы изменить биоэлектрическое состояние желаемым образом; [83] [89] [90] и
- Вычислительные платформы, которые выходят в онлайн [91] [92] для оказания помощи в построении прогнозирующих моделей биоэлектрической динамики в тканях. [93] [94] [95]
По сравнению с электродными методами молекулярные зонды обеспечивают более широкое пространственное разрешение и облегчают динамический анализ с течением времени. Хотя калибровка или титрование возможны, молекулярные зонды обычно являются полуколичественными, тогда как электроды обеспечивают абсолютные биоэлектрические значения. Еще одним преимуществом флуоресцентных и других зондов является их менее инвазивная природа и пространственное мультиплексирование, позволяющее одновременно контролировать большие площади эмбриональных или других тканей in vivo во время нормальных или патологических процессов паттерна. [96]
Роли в организмах
[ редактировать ]Раннее развитие
[ редактировать ]Работа над модельными системами, такими как Xenopus laevis и рыбка данио, выявила роль биоэлектрической передачи сигналов в развитии сердца. [97] [98] лицо, [99] [100] глаз, [89] мозг, [101] [102] и другие органы. Скрининги выявили роль ионных каналов в контроле размера таких структур, как плавник рыбки данио, [103] в то время как целенаправленные исследования по увеличению функций показали, например, что части тела могут быть переопределены на уровне органов – например, создание целых глаз в энтодерме кишечника . [89] Как и в головном мозге, биоэлектрика развития может интегрировать информацию на значительном расстоянии в эмбрионе, например, например, при контроле размера мозга с помощью биоэлектрических состояний вентральной ткани. [102] и контроль онкогенеза в месте экспрессии онкогена по биоэлектрическому состоянию удаленных клеток. [104] [105]
Заболевания человека, а также многочисленные мутанты мышей показывают, что биоэлектрическая передача сигналов важна для развития человека (таблицы 1 и 2). Эти эффекты повсеместно связаны с каналопатиями — заболеваниями человека, возникающими в результате мутаций, разрушающих ионные каналы.
Некоторые каналопатии приводят к морфологическим аномалиям или врожденным дефектам в дополнение к симптомам, поражающим мышцы и/или нейроны. Например, мутации, которые нарушают внутренний выпрямляющий калиевый канал Kir2.1, вызывают доминантно наследуемый синдром Андерсена-Тавиля (ATS). У пациентов с САТ наблюдаются периодические параличи , сердечные аритмии и множественные морфологические аномалии, которые могут включать расщелину или высокое арочное небо, расщелину или тонкую верхнюю губу, уплощенный желобок , микрогнатию зубов , олигодонтию , гипоплазию эмали, задержку прорезывания зубов, неправильный прикус, широкий лоб, широкий посадку. глаза, низко посаженные уши, синдактилия , клинодактилия , брахидактилия и дисплазия почек. [106] [107] Мутации, разрушающие другой внутренне исправляющий К + канал Girk2, кодируемый KCNJ6, вызывает синдром Кеппена-Любинского , который включает микроцефалию , узкую переносицу, высокое арочное небо и тяжелую генерализованную липодистрофию (неспособность генерировать жировую ткань). [108] KCNJ6 находится в критической области синдрома Дауна , так что дупликации, включающие эту область, приводят к черепно-лицевым аномалиям и аномалиям конечностей, а дупликации, не включающие эту область, не приводят к морфологическим симптомам синдрома Дауна. [109] [110] [111] [112] Мутации в KCNH1 , потенциалзависимом калиевом канале, приводят к Темпла-Барайтсера (также известному как Циммерманна-Лабанда синдрому ). Общие черты синдрома Темпла-Барайтсера включают отсутствие или гипоплазию ногтей и фаланг пальцев рук и ног, а также нестабильность суставов. Черепно-лицевые дефекты, связанные с мутациями в KCNH1, включают расщелину или высокое арочное небо, гипертелоризм , дисморфию ушей, дисморфию носа, гипертрофию десен и аномальное количество зубов. [113] [114] [115] [116] [117] [118] [119] [ чрезмерное цитирование ]
Мутации в CaV1.2 , потенциалзависимом Ca 2+ канала, приводят к синдрому Тимоти , который вызывает тяжелую сердечную аритмию (удлиненный интервал QT) наряду с синдактилией и черепно-лицевыми дефектами, сходными с синдромом Андерсена-Тавиля, включая расщелину или высокое арочное небо, микрогнатию, низко посаженные уши, синдактилию и брахидактилию. [120] [121] Хотя эти каналопатии встречаются редко, они показывают, что функциональные ионные каналы важны для развития. Кроме того, внутриутробное воздействие противоэпилептических препаратов, воздействующих на некоторые ионные каналы, также приводит к увеличению частоты врожденных дефектов, таких как расщелина рта. [122] [123] [124] [125] [126] [ чрезмерное цитирование ] Эффекты как генетического, так и экзогенного нарушения ионных каналов позволяют понять важность биоэлектрической передачи сигналов в развитии.
Заживление ран и руководство клетками
[ редактировать ]Одна из наиболее изученных ролей биоэлектрических градиентов связана с эндогенными электрическими полями на уровне тканей, используемыми во время заживления ран. Изучать электрические поля, связанные с ранами, сложно, поскольку эти поля слабы, менее колеблются и не вызывают немедленных биологических реакций по сравнению с нервными импульсами и мышечными сокращениями. Разработка вибрирующих и стеклянных микроэлектродов продемонстрировала, что раны действительно производят и, что важно, поддерживают измеримые электрические токи и электрические поля. [41] [127] [60] [128] [129] [130] Эти методы позволяют дополнительно охарактеризовать электрические поля/токи ран при ранах роговицы и кожи, которые демонстрируют активные пространственные и временные характеристики, что позволяет предположить активную регуляцию этих электрических явлений. Например, электрический ток раны всегда самый сильный на краю раны, который постепенно увеличивается и достигает пика примерно через 1 час после травмы. [131] [132] [62] При ранах у животных, больных диабетом , электрические поля раны значительно ухудшаются. [133] Ожидается, что понимание механизмов генерации и регуляции электрических токов/полей в ране откроет новые подходы к управлению электрическим аспектом для лучшего заживления ран.
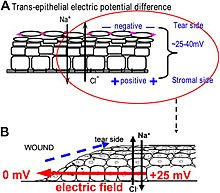
Как возникают электрические поля в ране? Эпителий активно перекачивает и дифференциально разделяет ионы. В эпителии роговицы, например, Na + и К + транспортируются внутрь из слезной жидкости во внеклеточную жидкость, а Cl − транспортируется из внеклеточной жидкости в слезную жидкость. Эпителиальные клетки соединяются плотными соединениями, образуя основной электрорезистивный барьер и, таким образом, создавая электрический градиент поперек эпителия – трансэпителиальный потенциал (ТЭП). [134] [135] Нарушение эпителиального барьера, как это происходит при любых ранах, создает отверстие, которое нарушает высокое электрическое сопротивление, создаваемое плотными соединениями в эпителиальном листке, локально замыкая эпителий. Таким образом, ТЭП на ране падает до нуля. Однако нормальный транспорт ионов продолжается в неповрежденных эпителиальных клетках за пределами края раны (обычно на расстоянии менее 1 мм), вытесняя поток положительных зарядов из раны и создавая устойчивое латерально-ориентированное электрическое поле (ЭП) с катодом в ране. Кожа также генерирует ТЭП, и когда на коже образуется рана, возникают аналогичные раневые электрические токи и поля до тех пор, пока барьерная функция эпителия не восстановится и не прекратит короткое замыкание в ране. Когда электрическими полями раны манипулируют фармакологическими агентами, которые либо стимулируют, либо ингибируют транспорт ионов, электрические поля раны также увеличиваются или уменьшаются соответственно. Заживление ран при ранениях роговицы можно соответственно ускорить или замедлить. [131] [132] [136]
Как электрические поля влияют на заживление ран? Для заживления ран клетки, окружающие рану, должны мигрировать и направленно расти в рану, чтобы покрыть дефект и восстановить барьер. Клетки, важные для заживления ран, удивительно хорошо реагируют на приложенные электрические поля той же силы, что и на ранах. Физиологические электрические поля влияют на весь спектр типов клеток и их реакцию после повреждения. К ним относятся миграция и деление эпителиальных клеток, прорастание и расширение нервов, а также миграция лейкоцитов и эндотелиальных клеток. [137] [138] [139] [140] Наиболее изученным клеточным поведением является направленная миграция эпителиальных клеток в электрических полях – электротаксис . Эпителиальные клетки мигрируют направленно к отрицательному полюсу (катоду), который при ране представляет собой полярность поля эндогенных векторных электрических полей в эпителии, направленную (от положительного к отрицательному) к центру раны. Эпителиальные клетки роговицы, кератиноциты кожи и многие другие типы клеток демонстрируют направленную миграцию при напряженности электрического поля всего несколько мВ·мм. −1 . [141] [142] [143] [144] Большие листы однослойных эпителиальных клеток и листы многослойных эпителиальных клеток также мигрируют направленно. [132] [145] Такое коллективное движение очень похоже на то, что происходит во время заживления ран in vivo, когда листы клеток коллективно перемещаются в ложе раны, чтобы покрыть рану и восстановить барьерную функцию кожи или роговицы.
Как клетки воспринимают такие мельчайшие внеклеточные электрические поля, остается неясным. Недавние исследования начали выявлять некоторые генетические, сигнальные и структурные элементы, лежащие в основе того, как клетки воспринимают небольшие физиологические электрические поля и реагируют на них. К ним относятся ионные каналы, внутриклеточные сигнальные пути, липидные рафты мембран и электрофорез компонентов клеточных мембран. [146] [147] [148] [149] [150] [151] [152] [ чрезмерное цитирование ]
Регенерация конечностей у животных
[ редактировать ]В начале 20-го века Альберт Мэтьюз принципиально связал регенерацию книдарийного полипа с разницей потенциалов между поверхностями полипа и столона и повлиял на регенерацию, наложив противотоки. Амедео Герлицка, следуя по стопам своего наставника дю Буа-Раймона, выдвинул теорию о том, что электрические токи играют раннюю роль в регенерации, возможно, инициируя пролиферацию клеток. [153] Используя электрические поля, подавляющие эндогенные, Марш и Бимс поразительным образом создали двуголовых планарий и даже полностью изменили полярность первичного тела: хвосты выросли там, где раньше существовала голова. [154] После этих исследований семян на протяжении десятилетий вплоть до сегодняшнего дня возникали вариации идеи о том, что биоэлектричество может чувствовать повреждение и запускать или, по крайней мере, играть важную роль в регенерации. Возможное объяснение лежит в потенциалах покоя (в первую очередь Vmem и TEP), которые могут быть, по крайней мере частично, спящими датчиками (сигнализациями), готовыми к обнаружению, и эффекторами (триггерами), готовыми реагировать на локальное повреждение. [127] [155] [156] [12]
Вслед за относительным успехом электростимуляции в отношении неразрешимой регенерации лягушачьей лапки с использованием имплантированного биметаллического стержня в конце 1960-х годов, [157] биоэлектрический внеклеточный аспект регенерации конечностей амфибий был широко изучен в последующие десятилетия. Получение точных описательных и функциональных физиологических данных стало возможным благодаря разработке сверхчувствительного вибрационного зонда и усовершенствованных прикладных устройств. [41] [158] Ампутация неизменно приводит к выведению тока через кожу и, как следствие, к возникновению бокового электрического поля, устанавливающего катод в месте раны. Хотя первоначально происходит чистая утечка ионов, со временем появляется активный компонент, и блокирование ионных транслокаторов обычно ухудшает регенерацию. С помощью биомиметических экзогенных электрических токов и полей была достигнута частичная регенерация, которая обычно включала рост тканей и увеличение нейрональной ткани. И наоборот, исключение или устранение эндогенного электрического тока и полей ухудшает регенерацию. [60] [159] [158] [160] Эти исследования по регенерации конечностей земноводных и связанные с ними исследования на миногах и млекопитающих. [161] в сочетании с переломов костей заживлением [162] [163] и исследования in vitro , [132] привело к общему правилу, согласно которому мигрирующие (например, кератиноциты, лейкоциты и эндотелиальные клетки) и вырастающие (например, аксоны) клетки, способствующие регенерации, подвергаются электротаксису по направлению к катоду (исходному месту повреждения). Соответственно, анод связан с резорбцией или дегенерацией тканей, что происходит при нарушенной регенерации и остеокластической резорбции в кости. [162] [160] [164] Несмотря на эти усилия, перспектива значительной эпиморфной регенерации у млекопитающих остается основным направлением будущих усилий, которые включают использование носимых биореакторов для создания среды, в которой можно управлять прорегенеративными биоэлектрическими состояниями. [165] [166] и продолжающиеся попытки электрической стимуляции. [167]
Недавние молекулярные исследования показали, что поток протонов и натрия важен для регенерации хвоста у головастиков Xenopus . [12] [168] [169] и показали, что регенерация всего хвоста (со спинным мозгом, мышцами и т. д.) может быть запущена в ряде обычно нерегенеративных состояний либо молекулярно-генетическими, либо молекулярно-генетическими, [170] фармакологический, [171] или оптогенетический [172] методы. У планарий работа над биоэлектрическим механизмом выявила контроль над поведением стволовых клеток. [173] контроль размеров при перепланировке, [174] передне-задняя полярность, [175] и форма головы. [69] [176] Изменение физиологической передачи сигналов, опосредованное щелевыми соединениями, приводит к появлению двуголовых червей у Dugesia japonica; Примечательно, что эти животные продолжают регенерировать как двуглавые в будущих раундах регенерации через несколько месяцев после того, как реагент, блокирующий щелевые соединения, покинул ткань. [177] [178] [179] Это стабильное, долговременное изменение анатомического строения, к которому животные регенерируют без геномного редактирования, является примером эпигенетического наследования структуры тела, а также единственным доступным «штаммом» видов планарий, демонстрирующим наследственное анатомическое изменение, отличающееся от других. от дикого типа. [180]



Рак
[ редактировать ]Отказ клеток от обычно жесткой координации деятельности в сторону анатомической структуры приводит к раку; поэтому неудивительно, что биоэлектричество – ключевой механизм координации роста и формирования структуры клеток – является мишенью, часто вызывающей рак и метастазирование. [181] [182] Действительно, давно известно, что щелевые соединения играют ключевую роль в канцерогенезе и прогрессировании. [183] [184] [185] Каналы могут вести себя как онкогены и поэтому подходят в качестве новых мишеней для лекарств. [3] [93] [183] [186] [187] [188] [189] [190] [191] [192] [ чрезмерное цитирование ] Недавняя работа на моделях амфибий показала, что деполяризация потенциала покоя может вызвать метастатическое поведение в нормальных клетках. [193] [194] в то время как гиперполяризация (вызванная неправильной экспрессией ионных каналов, лекарствами или светом) может подавлять онкогенез, вызванный экспрессией онкогенов человека. [195] Деполяризация потенциала покоя, по-видимому, является биоэлектрическим признаком, с помощью которого можно неинвазивно обнаружить зарождающиеся участки опухоли. [196] Уточнение биоэлектрической сигнатуры рака в биомедицинском контексте как диагностического метода является одним из возможных применений в этой области. [181] Примечательно, что амбивалентность полярности – деполяризация как маркер и гиперполяризация как лечение – делает концептуально возможным создание терапевтических подходов (сочетание терапии с диагностикой), предназначенных для одновременного выявления и лечения ранних опухолей, в данном случае основанных на нормализации мембраны. поляризация. [195]
Регулирование шаблона
[ редактировать ]Недавние эксперименты с использованием препаратов, открывающих/блокирующих ионные каналы, а также неправильной экспрессии доминантных ионных каналов на ряде модельных видов показали, что биоэлектричество, в частности, градиенты напряжения, инструктируют не только поведение стволовых клеток. [197] [198] [199] [200] [201] [202] [ чрезмерное цитирование ] но и крупномасштабный рисунок. [29] [203] [204] Сигналы формирования паттерна часто опосредуются пространственными градиентами потенциалов покоя клеток, или Vmem, которые могут трансдуцироваться в каскады вторичных мессенджеров и транскрипционные изменения с помощью нескольких известных механизмов. Эти потенциалы задаются функцией ионных каналов и насосов и формируются соединениями щелевых соединений, которые создают компартменты развития (изопотенциальные клеточные поля). [205] Поскольку и щелевые переходы, и ионные каналы сами по себе чувствительны к напряжению, группы клеток реализуют электрические цепи с богатыми возможностями обратной связи. Результаты биоэлектрической динамики развития in vivo представляют собой крупномасштабные решения по паттерну, такие как количество голов у планарий, [179] форма лица в развитии лягушки, [99] и размер хвостов у рыбок данио. [103] Экспериментальная модуляция эндогенных биоэлектрических препаттернов позволила преобразовать участки тела (например, кишечник) в целый глаз. [89] стимуляция регенерации придатков, таких как хвосты головастиков , в нерегенеративных условиях, [172] [171] [170] и преобразование формы и содержимого головы плоских червей в модели, соответствующие другим видам плоских червей, несмотря на нормальный геном. [176] Недавняя работа показала использование сред физиологического моделирования для определения прогностических вмешательств, нацеленных на биоэлектрические состояния для восстановления эмбриональных дефектов головного мозга при ряде генетических и фармакологически индуцированных тератологий. [90] [101]
Будущие исследования
[ редактировать ]Жизнь, в конечном счете, представляет собой электрохимическое предприятие; исследования в этой области развиваются по нескольким направлениям. Во-первых, это редуктивная программа понимания того, как производятся биоэлектрические сигналы, как изменения напряжения в клеточной мембране могут регулировать поведение клеток и каковы генетические и эпигенетические последующие мишени биоэлектрических сигналов. Уже известно несколько механизмов, которые преобразуют биоэлектрические изменения в изменения экспрессии генов, включая биоэлектрический контроль движения небольших молекул-посредников через клетки, включая серотонин и бутират, потенциал-чувствительные фосфатазы и другие. [206] [207] Также известны многочисленные гены-мишени передачи сигналов напряжения, такие как Notch , BMP , FGF и HIF-1α . [128] Таким образом, проксимальные механизмы биоэлектрической передачи сигналов внутри отдельных клеток становятся хорошо понятными, а оптогенетика достигает успехов. [80] [172] [4] [208] [209] [ чрезмерное цитирование ] и магнитогенетика [210] продолжать содействовать этой исследовательской программе. Однако более сложной является интегративная программа понимания того, как конкретные закономерности биоэлектрической динамики помогают контролировать алгоритмы, которые осуществляют крупномасштабную регуляцию закономерностей (регенерацию и развитие сложной анатомии). Включение биоэлектрики с химической передачей сигналов в развивающуюся область исследования сенсорного восприятия клеток и принятия решений. [211] [212] [213] [214] [215] [216] [ чрезмерное цитирование ] является важным рубежом для будущей работы.
Биоэлектрическая модуляция продемонстрировала контроль над сложным морфогенезом и ремоделированием, а не просто устанавливает идентичность отдельных клеток. Более того, ряд ключевых результатов в этой области показал, что биоэлектрические цепи нелокальны — области тела принимают решения на основе биоэлектрических событий на значительном расстоянии. [101] [104] [105] Такие неклеточно-автономные события предполагают распределенные сетевые модели биоэлектрического контроля; [217] [218] [219] Возможно, потребуется разработать новые вычислительные и концептуальные парадигмы, чтобы понять обработку пространственной информации в биоэлектрически активных тканях. Было высказано предположение, что результаты из областей примитивного познания и нетрадиционных вычислений имеют отношение к делу. [218] [220] [69] к программе взлома биоэлектрического кода. Наконец, усилия в области биомедицины и биоинженерии направлены на разработку таких приложений, как носимые биореакторы для доставки реагентов, модифицирующих напряжение, к местам ран. [166] [165] и препараты, модифицирующие ионные каналы (разновидность электротерапевтических средств) для устранения врожденных дефектов. [90] и восстановительный ремонт. [171] Синтетические биологи также начинают включать биоэлектрические цепи в гибридные конструкции. [221]
Таблица 1: Ионные каналы и насосы, участвующие в формировании паттерна
| Белок | Морфогенетическая роль или фенотип LOF (потеря функции) | Разновидность | Ссылка |
|---|---|---|---|
| ТРХ1 К + транспортер | Оформление корневых волос | Арабидопсис | [222] |
| Кир2.1 калиевый канал | Рисунок крыльев | Дрозофила | [223] |
| Kir7.1 K + канал | Черепно-лицевой рисунок, развитие легких | Мускулистые мышцы | [224] |
| №2 Нет + /ЧАС + обменник | Эпителиальный рисунок | Дрозофила | [225] |
| Протонный насос V-АТФазы | Узор волос на крыльях, Пигментация и рисунок мозга, Черепно-лицевой рисунок. | Дрозофила, Oryzias latipes, Homo sapiens. | [226] [227] [228] |
| HCN1, Кв3,1 К + каналы | Формирование переднего мозга | Мускулистые мышцы | [229] [230] |
| КСНК1 К + канал | Дефицит роста | Мускулистые мышцы | [231] |
| ТВИК-1 К + канал (КСНК1) | Размер сердца (предсердий) | Мускулистые мышцы | [232] |
| KCNJ6 К + канал | Синдром Кеппена-Любинского – черепно-лицевой и головной мозг | Мудрый человек | [108] |
| KCNH1 (hEAG1) К + канал и протонный насос ATP6V1B2 V-ATPase | Синдром Циммермана-Лабанда и Темпла-Барайтцера – черепно-лицевые и мозговые дефекты, дисплазия/аплазия ногтей большого пальца руки и ноги. | Мудрый человек | [116] [233] |
| Хлоридный канал GLRa4 | Черепно-лицевые аномалии | Мудрый человек | [234] |
| KCNJ8 К + | Синдром Канту – дефекты лица, сердца, скелета, головного мозга | Мудрый человек | [235] [236] [237] |
| НАЛЦН (Na + канал утечки) | Синдром Фримена-Шелдона – конечности, лицо, мозг | Мудрый человек | [238] |
| Хлоридный канал CFTR | Двустороннее отсутствие семявыносящих протоков | Мудрый человек | [239] [240] |
| КСНК1 | Дисморфии головы/лица | Мудрый человек | [241] |
| КСНК9, ТАСК3 К + каналы | Синдром дисморфизма Бирка-Бареля – черепно-лицевые дефекты, дефекты головного мозга (коркового рисунка). | Мудрый человек | [242] [243] [244] |
| Кир6,2 К + канал | Черепно-лицевые дефекты | Мудрый человек | [244] |
| KCNQ1 К + канал (посредством эпигенетической регуляции) | Гипертрофия языка, печени, селезенки, поджелудочной железы, почек, надпочечников, половых органов – синдром Беквита-Видемана; черепно-лицевые дефекты и дефекты конечностей, раннее развитие | Homo sapiens, Mus musculus, дрозофила. | [245] [246] [247] [248] |
| KCNQ1 К + канал | Синдром Джервелла и Ланге-Нильсена – внутреннее ухо и конечности | Мудрый человек, Mus musculus | [249] [250] [251] |
| Кир2,1 К + канал (KNCJ2) | Синдром Андерсена-Тавиля – черепно-лицевой, конечностей, ребер. | Мудрый человек, Mus musculus | [106] [223] [252] |
| Рецептор ГАМК-А (хлоридный канал) | Синдром Ангельмана – черепно-лицевой (например, расщелина неба) и паттерн на руках | Мудрый человек, Mus musculus | [253] [254] [255] |
| Хлоридный канал TMEM16A | Морфогенез трахеи | Мускулистые мышцы | [256] |
| Гирк2 К + канал | Дефекты развития мозжечка | Мускулистые мышцы | [257] [258] [259] [260] |
| КСНХ2 К + канал | Сердечные и черепно-лицевые дефекты рисунка. | Мускулистые мышцы | [261] |
| KCNQ1 К + канал | Аномалии прямой кишки, поджелудочной железы и желудка. | Мускулистые мышцы | [262] |
| НаВ1.2 | Дефекты восстановления мышц и нервов | Ксеноп | [171] |
| Кир6,1 К + канал | Дефекты рисунка глаз | Ксеноп | [89] |
| Ионный насос V-АТФаза | Дефекты лево-правой асимметрии, восстановление мышц и нервов | Ксеноп, домашний петух Галл, Даниус рерио | [170] [82] |
| Ионный насос H,K-АТФаза | Дефекты лево-правой асимметрии | Ксенопус, Эхинодея | [263] [264] [265] |
| Kir7.1 K + канал | Дефекты развития меланосом | Данио рерио | [266] |
| КВ каналы | Регулировка размера плавников, регулировка размера сердца | Даниус рерио, Mus musculus | [103] [267] |
| НаВ 1,5, На + /К + -АТФаза | Сердечный морфогенез | Данио рерио | [268] [269] |
| КСНК3 | Доминантные мутации вызывают дисплазию мозжечка у человека, а также дефекты жилкования крыльев и глаз у дрозофилы. | Мудрец, Дрозофила | [270] |
Таблица 2. Щелевые соединения, участвующие в формировании рисунка
| Белок щелевого соединения | Морфогенетическая роль или фенотип LOF | Разновидность | Ссылки |
|---|---|---|---|
| Иннексины | Морфогенез гонад и зародышевой линии | К. Элеганс | [271] |
| Иннексин1,2 | Формирование кутикулы (эпителия), развитие передней кишки | Дрозофила | [272] [273] |
| Иннексин 2 | Размер глаз | Дрозофила | [274] |
| Cx43 | Окулоденто-пальцевая дисплазия (ODDD), пороки сердца (выносящий тракт и конотрункальный), рандомизация лево-правой асимметрии, проблемы дифференцировки остеобластов, черепно-лицевые дефекты, миогенез | Человек разумный, Mus musculus, Gallus Gallus Domestica. | [275] [276] [277] [278] [279] [280] [281] [282] [283] [284] [ чрезмерное цитирование ] |
| Cx37 | Формирование лимфатической системы | Мускулистые мышцы | [285] [286] |
| Cx45 | Пороки сердца (подушкообразный рисунок) | Мускулистые мышцы | [287] [288] |
| Сх50, Сх46 | Дефекты глаз (проблемы дифференцировки и пролиферации, особенно хрусталика), | Мускулистые мышцы | [289] |
| Cx26 | Дефекты развития улитки | Мускулистые мышцы | [290] |
| Cx41.8 | Дефекты пигментного рисунка | Данио рерио | [291] |
| Cx43 | Регулировка размера и рисунка плавников Краниофронтоназальный синдром | Даниус рерио, Mus musculus | [292] [293] [294] [295] |
| Вх4, Вх2 | Дифференциация зародышевой линии и сперматогенез | Дрозофила | [296] |
| Паннексин3 | Развитие скелета | Мускулистые мышцы | [297] |
Таблица 3: Онкогены ионных каналов
| Белок | Разновидность | Ссылки | Рак-роль |
|---|---|---|---|
| НаВ 1,5 канала | Мудрый человек | [298] [299] | Онкоген |
| ЭРГ калиевые каналы | Мудрый человек | [300] [301] | Онкоген |
| 9 калиевый канал | Мускулистые мышцы | [302] | Онкоген |
| Дуктин (компонент протонной V-АТФазы) | Мускулистые мышцы | [303] | Онкоген |
| SLC5A8 транспортер натрия/бутирата | Мудрый человек | [304] | Онкоген |
| KCNE2 калиевый канал | Мускулистые мышцы | [305] | Онкоген |
| KCNQ1 калиевый канал | Хомо Сапиенс , мышь | [246] [262] [306] | Онкоген |
| SCN5A потенциалзависимый натриевый канал | Мудрый человек | [299] | Онкоген |
| Метаботропный глутаматный рецептор | Мускулистые мышцы , Человек | [307] [308] | Онкоген |
| Хлоридный канал CFTR | Мудрый человек | [309] [310] | Супрессор опухоли |
| Коннексин43 | Мудрый человек | [311] | Супрессор опухоли |
| БКСа | Мудрый человек | [312] | Онкоген |
| Мускариновый рецептор ацетилхолина | Мудрый человек, Mus musculus | [313] | Супрессор опухоли |
| KCNJ3 (Гирк) | Мудрый человек | [314] [315] | Онкоген |
Ссылки
[ редактировать ]- ^ Левин, Майкл (2011). «Мудрость тела: будущие методы и подходы к морфогенетическим областям в регенеративной медицине, биологии развития и раке». Регенеративная медицина . 6 (6): 667–673. дои : 10.2217/rme.11.69 . ПМИД 22050517 .
- ^ Левин, М (2014). «Молекулярное биоэлектричество: как эндогенные потенциалы напряжения контролируют поведение клеток и регулируют структуру in vivo» . Молекулярная биология клетки . 25 (24): 3835–3850. дои : 10.1091/mbc.E13-12-0708 . ПМК 4244194 . ПМИД 25425556 .
- ^ Jump up to: а б с Бейтс, Эмили (2015). «Ионные каналы в развитии и раке» . Ежегодный обзор клеточной биологии и биологии развития . 31 : 231–247. doi : 10.1146/annurev-cellbio-100814-125338 . ПМИД 26566112 .
- ^ Jump up to: а б Коэн, Адам Э; Венкатачалам, Вина (2014). «Привнесение биоэлектричества в свет» . Ежегодный обзор биофизики . 43 : 211–232. doi : 10.1146/annurev-biophys-051013-022717 . ПМИД 24773017 .
- ^ Фанк, Р.Х; Монси, Т; Озкукур, Н. (2009). «Электромагнитные эффекты – от клеточной биологии к медицине». Прогресс в гистохимии и цитохимии . 43 (4): 177–264. дои : 10.1016/j.proghi.2008.07.001 . ПМИД 19167986 .
- ^ Фанк, Р.Х; Монси, Т.К. (2006). «Воздействие электромагнитных полей на клетки: Физиолого-терапевтические подходы и молекулярные механизмы взаимодействия. Обзор» . Клетки Ткани Органы . 182 (2): 59–78. дои : 10.1159/000093061 . ПМИД 16804297 . S2CID 10705650 .
- ^ Jump up to: а б Чжао, Мин; Чалмерс, Лаура; Цао, Линь; Виейра, Ана С; Маннис, Марк; Рид, Брайан (2012). «Электрическая передача сигналов в контроле поведения глазных клеток» . Прогресс в исследованиях сетчатки и глаз . 31 (1): 65–88. doi : 10.1016/j.preteyeres.2011.10.001 . ПМЦ 3242826 . ПМИД 22020127 .
- ^ Jump up to: а б с д Левин, Майкл; Мартынюк, Кристофер Дж (2018). «Биоэлектрический код: старая вычислительная среда для динамического контроля роста и формы» . Биосистемы . 164 : 76–93. doi : 10.1016/j.biosystems.2017.08.009 . ПМЦ 10464596 . ПМИД 28855098 .
- ^ Лейн, Н.; Аллен, Дж. Ф.; Мартин, В. (2010). «Как LUCA зарабатывала на жизнь? Хемиосмос в зарождении жизни». Биоэссе . 32 (4): 271–280. дои : 10.1002/bies.200900131 . ПМИД 20108228 .
- ^ Лейн, Н.; Мартин, В.Ф. (2012). «Происхождение мембранной биоэнергетики» . Клетка . 151 (7): 1406–16. дои : 10.1016/j.cell.2012.11.050 . ПМИД 23260134 .
- ^ Jump up to: а б Люксарди, Дж; Рид, Б; Майяр, П; Чжао, М (2014). «Одноклеточная рана генерирует цепь электрического тока и изменения потенциала клеточной мембраны, что требует притока кальция» . Интегр. Биол . 6 (7): 662–672. дои : 10.1039/c4ib00041b . ПМИД 24801267 . S2CID 7313742 .
- ^ Jump up to: а б с Феррейра, Фернандо; Люксарди, Гийом; Рид, Брайан; Чжао, Мин (2016). «Ранняя биоэлектрическая активность опосредует окислительно-восстановительную регенерацию» . Разработка . 143 (24): 4582–4594. дои : 10.1242/dev.142034 . ПМК 5201032 . ПМИД 27827821 .
- ^ Робинсон, К.; Мессерли, М. (1996). «Электрические эмбрионы: эмбриональный эпителий как генератор информации о развитии». В Маккейге, С. (ред.). Рост нервов и руководство . Портленд. стр. 131–141.
- ^ Маклафлин, К.А.; Левин, М (2018). «Биоэлектрическая передача сигналов при регенерации: механизмы ионного контроля роста и формы» . Биология развития . 433 (2): 177–189. дои : 10.1016/j.ydbio.2017.08.032 . ПМЦ 5753428 . ПМИД 29291972 .
- ^ Jump up to: а б с Левин, Майкл; Пеццуло, Джованни; Финкельштейн, Джошуа М (2017). «Эндогенные биоэлектрические сигнальные сети: использование градиентов напряжения для контроля роста и формы» . Ежегодный обзор биомедицинской инженерии . 19 : 353–387. doi : 10.1146/annurev-bioeng-071114-040647 . ПМЦ 10478168 . ПМИД 28633567 .
- ^ Питкэрн, Эмили; Маклафлин, Келли А. (2016). «Биоэлектрическая передача сигналов координирует решения по формированию паттерна во время эмбриогенеза» . Тенденции в биологии развития . 9 : 1–9.
- ^ Пуллар, CE. Физиология биоэлектричества в развитии, регенерации тканей и раке (CRC Press, 1996). [ нужна страница ]
- ^ Нуччителли, Р. (2003). «Роль эндогенных электрических полей в заживлении ран». Актуальные темы биологии развития . 58 : 1–26. дои : 10.1016/s0070-2153(03)58001-2 . ISBN 978-0-12-153158-4 . ПМИД 14711011 .
- ^ Кларк, Эдвин (1987). Истоки нейробиологических концепций девятнадцатого века . Джасина, Л.С. Беркли: Издательство Калифорнийского университета. ISBN 0-520-05694-9 . OCLC 13456516 .
- ^ Пера, Марчелло (1992). Неоднозначная лягушка: спор Гальвани-Вольта о животном электричестве . Тр. Мандельбаум, Джонатан. Принстон, Нью-Джерси: Издательство Принстонского университета. ISBN 978-1-4008-6249-8 . OCLC 889251161 .
- ^ Пикколино, Марко; Бресадола, Марко (2013). Шокирующие лягушки: Гальвани, Вольта и электрические истоки нейробиологии . Оксфорд; Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-978221-5 . OCLC 859536612 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Маден, М. История исследований регенерации. (Издательство Кембриджского университета, 1991). [ нужна страница ]
- ^ Jump up to: а б с Маккейг, Колин Д.; Райничек, Энн М; Сонг, Бинг; Чжао, Мин (2005). «Электрический контроль над поведением клеток: текущие взгляды и будущий потенциал». Физиологические обзоры . 85 (3): 943–978. doi : 10.1152/physrev.00020.2004 . ПМИД 15987799 .
- ^ Бернштейн, Дж (1868). «О динамике отрицательного колебания нервного тока» . Архив Пфлюгера по всей физиологии человека и животных (на немецком языке). 1 (1): 173–207. дои : 10.1007/BF01640316 . S2CID 32435163 .
- ^ Дюбуа-Реймон, Эмиль (1848). «Исследования по животному электричеству» . Анналы физики и химии (на немецком языке). 151 (11): 463–464. Бибкод : 1848АнП...151..463Д . дои : 10.1002/andp.18481511120 .
- ^ Шютце, Стивен М (1983). «Открытие потенциала действия». Тенденции в нейронауках . 6 : 164–8. дои : 10.1016/0166-2236(83)90078-4 . S2CID 53175297 .
- ^ Дюбуа-Реймон, Эмиль (1860). по электричеству животных Исследования ( на немецком языке). Берлин: Георг Раймер. [ нужна страница ]
- ^ Финкельштейн, Габриэль (2013). Эмиль дю Буа-Реймон: нейробиология, личность и общество в Германии девятнадцатого века . Массачусетский технологический институт Пресс. ISBN 978-1-4619-5032-5 . OCLC 864592470 .
- ^ Jump up to: а б с Левин, Майкл; Стивенсон, Клэр Дж. (2012). «Регуляция поведения клеток и структуры тканей с помощью биоэлектрических сигналов: проблемы и возможности биомедицинской инженерии» . Ежегодный обзор биомедицинской инженерии . 14 : 295–323. doi : 10.1146/annurev-bioeng-071811-150114 . ПМЦ 10472538 . ПМИД 22809139 .
- ^ Мэтьюз, Альберт П. (1903). «Электрическая полярность в гидроидах». Американский журнал физиологии. Устаревший контент . 8 (4): 294–299. дои : 10.1152/ajplegacy.1903.8.4.294 .
- ^ Хайд, Ида Х. (1904). «Различия в электрическом потенциале развивающихся яиц». Американский журнал физиологии. Устаревший контент . 12 (3): 241–275. дои : 10.1152/ajplegacy.1904.12.3.241 .
- ^ Морган, TH; Даймон, Эбигейл К. (1904). «Исследование проблем физиологической «полярности» и электрической полярности у дождевого червя». Журнал экспериментальной зоологии . 1 (2): 331. Бибкод : 1904JEZ.....1..331M . дои : 10.1002/jez.1400010206 . hdl : 2027/hvd.32044107333023 .
- ^ Фрейзи, Орен Э. (1909). «Влияние электростимуляции на скорость регенерации Rana pipiens и Amblystoma jeffersonianum». Журнал экспериментальной зоологии . 7 (3): 457–475. Бибкод : 1909JEZ.....7..457F . дои : 10.1002/jez.1400070304 .
- ^ Лунд, Э.Дж. (1917). «Обратимость морфогенетических процессов у бурсарий». Журнал экспериментальной зоологии . 24 (1): 1–33. Бибкод : 1917JEZ....24....1L . дои : 10.1002/jez.1400240102 .
- ^ Хайман, Либби Генриетта (15 сентября 1992 г.). Сравнительная анатомия позвоночных Хаймана . Издательство Чикагского университета. стр. 192–236. ISBN 978-0-226-87013-7 .
- ^ Хайман, Либби Генриетта (1918). «Специальные статьи». Наука . 48 (1247): 518–524. дои : 10.1126/science.48.1247.518 . ПМИД 17795612 .
- ^ Лунд, Э. Биоэлектрические злодеи и рост (Техасский университет Press, 1947). [ нужна страница ]
- ^ Берр, HS; Нортроп, FSC (1935). «Электродинамическая теория жизни». Ежеквартальный обзор биологии . 10 (3): 322–333. дои : 10.1086/394488 . JSTOR 2808474 . S2CID 84480134 .
- ^ Марш, Г.; Бимс, HW (1949). «Электрический контроль осевой полярности регенерирующего кольчатого червя». Анатомическая запись . 105 (3): 513–514.
- ^ Марш, Г.; Бимс, HW (1947). «Электрический контроль полярности роста у регенерирующей Dugesia tigrina». Труды Федерации . 6 (1 ч. 2): 163. PMID 20342775 .
- ^ Jump up to: а б с д и Яффе, Лайонел Ф.; Нуччителли, Ричард (1974). «Сверхчувствительный вибрирующий зонд для измерения постоянных внеклеточных токов» . Журнал клеточной биологии . 63 (2): 614–28. дои : 10.1083/jcb.63.2.614 . ПМК 2110946 . ПМИД 4421919 .
- ^ Яффе, Л. (1982). «Развитие токов, напряжений и градиентов» . Порядок развития, его происхождение и регулирование . стр. 183–215 . ISBN 978-0-8451-1501-5 .
- ^ Яффе, Л.Ф. (1981). «Роль ионных токов в установлении закономерностей развития» . Философские труды Королевского общества B: Биологические науки . 295 (1078): 553–566. Бибкод : 1981RSPTB.295..553J . дои : 10.1098/rstb.1981.0160 . JSTOR 2395645 . ПМИД 6117911 .
- ^ Нуччителли, Ричард (1995). «Эндогенные электрические поля, измеренные в развивающихся эмбрионах». Электромагнитные поля . Достижения химии. Том. 250. С. 109–24. дои : 10.1021/ba-1995-0250.ch007 . ISBN 978-0-8412-3135-1 .
- ^ Яффе, LF; Нуччителли, Р. (1977). «Электрическое управление развитием». Ежегодный обзор биофизики и биоинженерии . 6 : 445–476. дои : 10.1146/annurev.bb.06.060177.002305 . ПМИД 326151 .
- ^ Боргенс, РБ (1986). «Роль естественных и прикладных электрических полей в регенерации и развитии нейронов». Прогресс клинических и биологических исследований . 210 : 239–250. ПМИД 3960913 .
- ^ Боргенс, Ричард Б. (1982). «Какова роль естественного электрического тока в регенерации и заживлении позвоночных?». Международный обзор цитологии, том 76 . Том. 76. С. 245–298. дои : 10.1016/S0074-7696(08)61793-3 . ISBN 978-0-12-364476-3 . ПМИД 6749746 .
- ^ Маккейг, Колин Д.; Райничек, Энн М.; Сонг, Бинг; Чжао, Мин (2002). «Нашло ли руководство по электрическому конусу роста свой потенциал?». Тенденции в нейронауках . 25 (7): 354–9. дои : 10.1016/S0166-2236(02)02174-4 . ПМИД 12079763 . S2CID 7534545 .
- ^ Конус, CD-младший; Тонжер, М. младший (1971). «Контроль митоза соматических клеток путем моделирования изменений уровня трансмембранного потенциала». Онкология . 25 (2): 168–182. дои : 10.1159/000224567 . ПМИД 5148061 .
- ^ Стиллвелл, EF; Конус, СМ; Конус, компакт-диск (1973). «Стимуляция синтеза ДНК в нейронах ЦНС путем устойчивой деполяризации». Новая биология природы . 246 (152): 110–111. дои : 10.1038/newbio246110a0 . ПМИД 4518935 .
- ^ Бинггели, Ричард; Вайнштейн, Рой К. (1986). «Мембранные потенциалы и натриевые каналы: гипотезы регуляции роста и образования рака, основанные на изменениях в натриевых каналах и щелевых соединениях». Журнал теоретической биологии . 123 (4): 377–401. Бибкод : 1986JThBi.123..377B . дои : 10.1016/S0022-5193(86)80209-0 . ПМИД 2443763 .
- ^ Ходжкин, А.Л.; Хаксли, AF (1939). «Потенциалы действия, записанные изнутри нервного волокна». Природа . 144 (3651): 710. Бибкод : 1939Natur.144..710H . дои : 10.1038/144710a0 . S2CID 4104520 .
- ^ Монтейро, Жоана; Айрес, Рита; Беккер, Йорг Д; Хасинто, Антонио; Серталь, Ана С; Родригес-Леон, Хоакин (2014). «Активность по накачке протонов V-АТФазы необходима для регенерации придатков взрослых рыбок данио» . ПЛОС ОДИН . 9 (3): е92594. Бибкод : 2014PLoSO...992594M . дои : 10.1371/journal.pone.0092594 . ПМЦ 3966808 . ПМИД 24671205 .
- ^ Кункель, Джозеф Г; Кордейру, София; Сюй, Ю (Джефф); Шипли, Алан М; Фейхо, Хосе А. (2006). «Использование неинвазивных ионоселективных микроэлектродных методов для изучения развития растений». Электрофизиология растений . стр. 109–137. дои : 10.1007/978-3-540-37843-3_5 . ISBN 978-3-540-32717-2 .
- ^ Шен, Ю; Пфлюгер, Т; Феррейра, Ф; Лян, Дж; Наведо, МФ; Цзэн, Кью; Рид, Б; Чжао, М (2016). «Диабетические раны роговицы производят значительно более слабые электрические сигналы, что может способствовать ухудшению заживления» . Научные отчеты . 6 : 26525. Бибкод : 2016NatSR...626525S . дои : 10.1038/srep26525 . ПМК 4901296 . ПМИД 27283241 .
- ^ Ходжкин, А.Л.; Хаксли, AF (1939). «Потенциалы действия, записанные изнутри нервного волокна». Природа . 144 (3651): 710–711. Бибкод : 1939Natur.144..710H . дои : 10.1038/144710a0 . S2CID 4104520 .
- ^ Грэм, Джудит; Джерард, Р.В. (1946). «Мембранные потенциалы и возбуждение проколотых одиночных мышечных волокон». Журнал клеточной и сравнительной физиологии . 28 (1): 99–117. дои : 10.1002/jcp.1030280106 . ПМИД 21002959 . S2CID 45361295 .
- ^ Чжао, Ю; Инайят, С; Дикин Д.А.; Сингер, Дж. Х.; Руофф, РС; Трой, Дж. Б. (2009). «Техника патч-зажима: обзор современного состояния и потенциального вклада наноинженерии». Труды Института инженеров-механиков, Часть N: Журнал наноинженерии и наносистем . 222 : 1–11. дои : 10.1243/17403499JNN149 . S2CID 53316098 .
- ^ Боргенс, Ричард Б; Ванейбл, Джозеф В.; Яффе, Лайонел Ф (1979). «Роль подкожных токовых шунтов в невозможности регенерации лягушек». Журнал экспериментальной зоологии . 209 (1): 49–56. Бибкод : 1979JEZ...209...49B . дои : 10.1002/jez.1402090106 . ПМИД 314968 .
- ^ Jump up to: а б с Боргенс, Р.Б.; Ванейбл, Дж. В.; Яффе, Л.Ф. (1977). «Биоэлектричество и регенерация. I. Инициирование регенерации конечностей лягушки минутными токами». Журнал экспериментальной зоологии . 200 (3): 403–416. Бибкод : 1977JEZ...200..403B . дои : 10.1002/jez.1402000310 . ПМИД 301554 .
- ^ Jump up to: а б Шипли, AM; Фейхо, Дж. А. (1999). «Использование метода вибрационного зонда для изучения устойчивых внеклеточных токов во время прорастания пыльцы и роста трубок». Оплодотворение у высших растений . стр. 235–252. дои : 10.1007/978-3-642-59969-9_17 . ISBN 978-3-642-64202-9 .
- ^ Jump up to: а б Рид, Брайан; Нуччителли, Ричард; Чжао, Мин (2007). «Неинвазивное измерение биоэлектрических токов вибрирующим зондом». Протоколы природы . 2 (3): 661–669. дои : 10.1038/нпрот.2007.91 . ПМИД 17406628 . S2CID 15237787 .
- ^ Кутрайбер, ВМ; Яффе, LF (1990). «Обнаружение внеклеточных градиентов кальция с помощью кальций-специфического вибрирующего электрода» . J Клеточная Биол . 110 (5): 1565–1573. дои : 10.1083/jcb.110.5.1565 . ПМК 2200169 . ПМИД 2335563 .
- ^ Люксарди, Гийом; Рид, Брайан; Феррейра, Фернандо; Майяр, Полина; Чжао, Мин (2015). «Измерение внеклеточных потоков ионов с использованием метода ионоселективного самореферентного микроэлектрода» . Журнал визуализированных экспериментов (99): e52782. дои : 10.3791/52782 . ПМК 4541607 . ПМИД 25993490 .
- ^ Тантама, Мэтью; Хун, Инь Пун; Йеллен, Гэри (2012). «Оптогенетические репортеры». Оптогенетика: инструменты контроля и мониторинга активности нейронов . Прогресс в исследованиях мозга. Том. 196. стр. 235–263. дои : 10.1016/B978-0-444-59426-6.00012-4 . ISBN 978-0-444-59426-6 . ПМК 3494096 . ПМИД 22341329 .
- ^ Чатни, Мохаммад Рамиз; Ли, Банда; Портерфилд, Дэвид Маршалл (2009). «Проектирование и приборное обеспечение системы оптродов для флуоресценции в частотной области со сроком службы без одновременного использования эталонного светодиода». Прикладная оптика . 48 (29): 5528–5536. Бибкод : 2009ApOpt..48.5528C . дои : 10.1364/AO.48.005528 . ПМИД 19823237 .
- ^ Сонг, Бинг; Гу, Ю; Пу, Джин; Рид, Брайан; Чжао, Чжицян; Чжао, Мин (2007). «Применение электрических полей постоянного тока к клеткам и тканям in vitro и модуляция электрического поля раны in vivo». Протоколы природы . 2 (6): 1479–1489. дои : 10.1038/нпрот.2007.205 . ПМИД 17545984 . S2CID 25924011 .
- ^ Чжао, Сивэй; Чжу, Кан; Чжан, Ян; Чжу, Цзыцзе; Сюй, Чжэнпин; Чжао, Мин; Пан, Тингруй (2014). «Электротаксис-на-чипе (ETC): интегрированная платформа количественного высокопроизводительного скрининга для миграции клеток, управляемой электрическим полем» . Лабораторный чип . 14 (22): 4398–4405. дои : 10.1039/C4LC00745J . ПМЦ 4437771 . ПМИД 25242672 .
- ^ Jump up to: а б с Салливан, Келли Дж; Эммонс-Белл, Майя; Левин, Майкл (2016). «Физиологические факторы регулируют видовую анатомию во время эмбриогенеза и регенерации» . Коммуникативная и интегративная биология . 9 (4): e1192733. дои : 10.1080/19420889.2016.1192733 . ПМЦ 4988443 . ПМИД 27574538 .
- ^ Борнат, Янник; Рау, Матье; Бутаиб, Юсеф; и др. (2010). «Обнаружение электрической активности бета-клеток поджелудочной железы с использованием микроэлектродных матриц». Пятый международный симпозиум IEEE по проектированию, испытаниям и приложениям электроники, 2010 г. (PDF) . стр. 233–236. дои : 10.1109/DELTA.2010.60 . ISBN 978-1-4244-6025-0 . S2CID 12107878 .
- ^ Кодзима, Дзюнъитиро; Синохара, Хироаки; Икарияма, Ёсихито; и др. (1991). «Электрически контролируемая пролиферация клеток карциномы человека, культивируемых на поверхности электрода». Журнал биотехнологии . 18 (1–2): 129–139. дои : 10.1016/0168-1656(91)90241-М . ПМИД 1367098 .
- ^ Лангхаммер, Кристофер Дж; Катцинг, Мелинда К; Луо, Винсент; и др. (2011). «Интеграция скелетных мышечных трубок с планарными микроэлектродными матрицами in vitro для пространственно-селективной записи и стимуляции: сравнение потенциалов внеклеточного действия нейронов и мышечных трубок» . Биотехнологический прогресс . 27 (3): 891–5. дои : 10.1002/btpr.609 . ПМЦ 4557870 . ПМИД 21574266 .
- ^ Маккаллен, Сет Д.; Маккуиллинг, Джон П.; Гроссфельд, Роберт М; и др. (2010). «Применение низкочастотных электрических полей переменного тока через встречно-штыревые электроды: влияние на жизнеспособность клеток, цитоплазматический кальций и остеогенную дифференцировку стволовых клеток человека, полученных из жировой ткани» . Тканевая инженерия. Часть C: Методы . 16 (6): 1377–86. дои : 10.1089/ten.tec.2009.0751 . ПМК 3003917 . ПМИД 20367249 .
- ^ Арьясомаяджула, Адитья; Дерикс, Джонатан; Перике, Шрикант; Герлах, Джеральд; Фанк, Р.Х. (2010). «Микроэлектродная матрица постоянного тока для исследования внутриклеточных ионных изменений». Биосенсоры и биоэлектроника . 26 (4): 1268–1272. дои : 10.1016/j.bios.2010.06.068 . ПМИД 20656468 .
- ^ Джаярам, Дханья Т; Ло, Цинцзе; Турсон, Скотт Б; Финли, Адам Х; Пейн, Кристин К. (2017). «Контроль мембранного потенциала покоя клеток с помощью проводящих полимерных микропроводов» . Маленький . 13 (27): 1700789. doi : 10.1002/smll.201700789 . ПМК 5560653 . ПМИД 28556571 .
- ^ Смит, Питер Дж.С.; Хаммар, Кэтрин; Портерфилд, Д. Маршалл; Сэнгер, Ричард Х; Тримарчи, Джеймс Р. (1999). «Самоссылающийся, неинвазивный, ион-селективный электрод для обнаружения в отдельных клетках потока кальция через трансплазматическую мембрану». Микроскопические исследования и техника . 46 (6): 398–417. doi : 10.1002/(SICI)1097-0029(19990915)46:6<398::AID-JEMT8>3.0.CO;2-H . ПМИД 10504217 . S2CID 25177705 .
- ^ Смит, Питер Дж.С.; Сэнгер, Ричард Х.; Мессерли, Марк А. (2006). «Принципы, разработка и применение самореферентных электрохимических микроэлектродов для определения потоков на клеточных мембранах» . У Майкла Адриан С.; Борланд, Лаура (ред.). Электрохимические методы нейробиологии . КПР. стр. 373–405. ISBN 978-1-4200-0586-8 . ПМИД 21204387 .
- ^ Синха, Гунджан (2013). «За счет инвестиций GSK продвигается аккумулятор электротехники» . Природная медицина . 19 (6): 654. doi : 10.1038/nm0613-654 . ПМИД 23744134 . S2CID 2260750 .
- ^ Фамм, Кристоффер; Литт, Брайан; Трейси, Кевин Дж; Бойден, Эдвард С; Слауи, Монсеф (2013). «Прыжок в электрохимию» . Природа . 496 (7444): 159–161. дои : 10.1038/496159а . ПМЦ 4179459 . ПМИД 23579662 .
- ^ Jump up to: а б Спенсер Адамс, Дэни; Лемир, Джоан М.; Крамер, Ричард Х.; Левин, Майкл (2014). «Оптогенетика в биологии развития: использование света для управления сигналами, зависящими от потока ионов, в эмбрионах Xenopus» . Международный журнал биологии развития . 58 (10–12): 851–861. дои : 10.1387/ijdb.140207ml . ПМЦ 10468825 . ПМИД 25896279 .
- ^ Адамс, Дэни С; Левин, Майкл (2006). «Обратный скрининг наркотиков: быстрый и недорогой метод выявления молекулярных мишеней» . Бытие . 44 (11): 530–540. дои : 10.1002/dvg.20246 . ПМК 3142945 . ПМИД 17078061 .
- ^ Jump up to: а б Адамс, Д.С.; Робинсон, КР; Фукумото, Т.; Юань, С; Альбертсон, Колорадо; Йелик, П; Куо, Л.; Максвини, М.; Левин, М. (2006). «На ранних этапах H+-V-АТФаза-зависимый поток протонов необходим для последовательного формирования лево-правого паттерна у немлекопитающих позвоночных» . Разработка . 133 (9): 1657–1671. дои : 10.1242/dev.02341 . ПМК 3136117 . ПМИД 16554361 .
- ^ Jump up to: а б Адамс, Дэни С; Левин, Майкл (2012). «Эндогенные градиенты напряжения как медиаторы межклеточной коммуникации: стратегии исследования биоэлектрических сигналов во время формирования паттернов» . Исследования клеток и тканей . 352 (1): 95–122. дои : 10.1007/s00441-012-1329-4 . ПМЦ 3869965 . ПМИД 22350846 .
- ^ Адамс, Д.С.; Левин, М (2012). «Общие принципы измерения мембранного потенциала покоя и концентрации ионов с использованием флуоресцентных репортеров биоэлектричества» . Протоколы Колд-Спринг-Харбора . 2012 (4): 385–397. дои : 10.1101/pdb.top067710 . ПМК 4001120 . ПМИД 22474653 .
- ^ Адамс, Д.С.; Левин, М (2012). «Измерение мембранного потенциала покоя с использованием репортеров флуоресцентного напряжения DiBAC4 (3) и CC2-DMPE» . Протоколы Колд-Спринг-Харбора . 2012 (4): 459–464. дои : 10.1101/pdb.prot067702 . ПМК 4001116 . ПМИД 22474652 .
- ^ Браунер, Томас; Хюльзер, Дитер Ф; Штрассер, Рето Дж (1984). «Сравнительные измерения мембранных потенциалов с помощью микроэлектродов и потенциалчувствительных красителей» . Biochimica et Biophysical Acta (BBA) – Биомембраны . 771 (2): 208–216. дои : 10.1016/0005-2736(84)90535-2 . ПМИД 6704395 .
- ^ Дил, Паркер Э; Кулкарни, Ришикеш У; Аль-Абдуллатиф, Сара Х; Миллер, Эван В. (2016). «Репортеры напряжения изомерно чистого тетраметилродамина» . Журнал Американского химического общества . 138 (29): 9085–9088. дои : 10.1021/jacs.6b05672 . ПМЦ 5222532 . ПМИД 27428174 .
- ^ Овьедо, Нью-Джерси; Николас, CL; Адамс, Д.С.; Левин, М (2008). «Живое изображение потенциала планарийной мембраны с использованием DiBAC4 (3)» . Протоколы Колд-Спринг-Харбора . 2008 (11): pdb.prot5055. дои : 10.1101/pdb.prot5055 . ПМЦ 10468776 . ПМИД 21356693 .
- ^ Jump up to: а б с д и Пай, В.П.; Ой, С; Шомрат, Т; Лемир, Дж. М.; Левин, М (2011). «Потенциал трансмембранного напряжения контролирует структуру глаз эмбриона у Xenopus laevis» . Разработка . 139 (2): 313–323. дои : 10.1242/dev.073759 . ПМК 3243095 . ПМИД 22159581 .
- ^ Jump up to: а б с Пай, Вайбхав П; Пиетак, Алексис; Уиллок, Валери; Да, Бин; Ши, Нянь-Цин; Левин, Майкл (2018). «HCN2 устраняет дефекты головного мозга, поддерживая предварительные шаблоны эндогенного напряжения» . Природные коммуникации . 9 (1): 998. Бибкод : 2018NatCo...9..998P . дои : 10.1038/s41467-018-03334-5 . ПМЦ 5843655 . ПМИД 29519998 .
- ^ Пиетак, Алексис; Левин, Майкл (2016). «Изучение инструктивной физиологической передачи сигналов с помощью механизма моделирования биоэлектрической ткани» . Границы биоинженерии и биотехнологии . 4 : 55. дои : 10.3389/fbioe.2016.00055 . ПМЦ 4933718 . ПМИД 27458581 .
- ^ Пиетак, Алексис; Левин, Майкл (2017). «Биоэлектрические гены и реакционные сети: компьютерное моделирование генетической, биохимической и биоэлектрической динамики в паттерн-регуляции» . Журнал интерфейса Королевского общества . 14 (134): 20170425. doi : 10.1098/rsif.2017.0425 . ПМК 5636277 . ПМИД 28954851 .
- ^ Jump up to: а б Сервера, Хавьер; Алькарас, Антонио; Мафе, Сальвадор (2016). «Биоэлектрические сигналы и ионные каналы в моделировании многоклеточных структур и биофизике рака» . Научные отчеты . 6 : 20403. Бибкод : 2016NatSR...620403C . дои : 10.1038/srep20403 . ПМЦ 4740742 . ПМИД 26841954 .
- ^ Сервера, Хавьер; Месегер, Сальвадор; Мафе, Сальвадор (2016). «Взаимодействие между генетической и биоэлектрической передачей сигналов позволяет осуществлять пространственную регионализацию мембранных потенциалов в модельных многоклеточных ансамблях» . Научные отчеты . 6 : 35201. Бибкод : 2016NatSR...635201C . дои : 10.1038/srep35201 . ПМК 5059667 . ПМИД 27731412 .
- ^ Сервера, Хавьер; Мансанарес, Хосе Антонио; Мафе, Сальвадор (2015). «Электрическая связь в ансамблях невозбудимых клеток: моделирование пространственной карты потенциалов отдельных ячеек». Журнал физической химии Б. 119 (7): 2968–2978. дои : 10.1021/jp512900x . ПМИД 25622192 .
- ^ Муто, Хироки; Перрон, Амели; Акеманн, Вальтер; Ивамото, Юка; Кнопфель, Томас (2011). «Оптогенетический мониторинг мембранных потенциалов» . Экспериментальная физиология . 96 (1): 13–18. doi : 10.1113/expphysicalol.2010.053942 . ПМИД 20851856 . S2CID 5265189 .
- ^ Питкэрн, Эмили; Харрис, Ханна; Эпини, Жюстин; Пай, Вайбхав П; Лемир, Джоан М; Да, Бин; Ши, Нянь-Цин; Левин, Майкл; Маклафлин, Келли А. (2017). «Координация морфогенеза сердца: новая роль гиперполяризационно-активируемых циклических нуклеотид-зависимых (HCN) каналов во время кардиогенеза у Xenopus laevis» . Коммуникативная и интегративная биология . 10 (3): e1309488. дои : 10.1080/19420889.2017.1309488 . ПМК 5501196 . ПМИД 28702127 .
- ^ Пай, Вайбхав П; Уиллок, Валери; Питкэрн, Эмили Дж; Лемир, Джоан М; Паре, Жан-Франсуа; Ши, Нянь-Цин; Маклафлин, Келли А; Левин, Майкл (2017). «Функция ионного канала HCN4 необходима для ранних событий, которые регулируют анатомическое лево-правое паттернирование узловым и левоасимметричным образом, независимо от экспрессии генов» . Биология Открытая . 6 (10): 1445–1457. дои : 10.1242/bio.025957 . ПМЦ 5665463 . ПМИД 28818840 .
- ^ Jump up to: а б Адамс, Дэни Спенсер; Узель, Себастьен Г.М; Акаги, Джин; Влодкович, Дональд; Андреева Виктория; Йелик, Памела Кротти; Девитт-Ли, Адриан; Паре, Жан-Франсуа; Левин, Майкл (2016). «Биоэлектрическая передача сигналов через калиевые каналы: механизм черепно-лицевого дисморфогенеза при синдроме Андерсена-Тавиля, связанном с KCNJ2» . Журнал физиологии . 594 (12): 3245–3270. дои : 10.1113/JP271930 . ПМК 4908029 . ПМИД 26864374 .
- ^ Ванденберг, Лаура Н; Морри, Райан Д; Адамс, Дэни Спенсер (2011). «V-АТФаза-зависимая эктодермальная напряженность и регионализация ph необходимы для краниофациального морфогенеза» . Динамика развития . 240 (8): 1889–1904. дои : 10.1002/dvdy.22685 . ПМЦ 10277013 . ПМИД 21761475 . S2CID 205768092 .
- ^ Jump up to: а б с Пай, В.П.; Лемир, Дж. М.; Паре, Ж.-Ф; Лин, Г; Чен, Ю; Левин, М (2015). «Эндогенные градиенты потенциала покоя формируют структуру эмбриональной нервной ткани посредством передачи сигналов Notch и регуляции пролиферации» . Журнал неврологии . 35 (10): 4366–85. doi : 10.1523/JNEUROSCI.1877-14.2015 . ПМК 4355204 . ПМИД 25762681 .
- ^ Jump up to: а б Пай, Вайбхав П; Лемир, Джоан М; Чен, Ин; Лин, Гуфа; Левин, Майкл (2015). «Локальные и дальнодействующие эндогенные градиенты потенциала покоя антагонистически регулируют апоптоз и пролиферацию в эмбриональной ЦНС» . Международный журнал биологии развития . 59 (7–8–9): 327–40. дои : 10.1387/ijdb.150197ml . ПМЦ 10505512 . ПМИД 26198142 .
- ^ Jump up to: а б с Ператонер, Саймон; Даан, Джейкоб М; Генрион, Ульрика; Сибом, Гвискар; Хигдон, Чарльз В.; Джонсон, Стивен Л; Нюсляйн-Фольхард, Кристиана; Харрис, Мэтью П. (2014). «Биоэлектрическая сигнализация регулирует размер плавников данио» . ПЛОС Генетика . 10 (1): e1004080. дои : 10.1371/journal.pgen.1004080 . ПМЦ 3894163 . ПМИД 24453984 .
- ^ Jump up to: а б Чернет, Брук Т; Филдс, Крис; Левин, Майкл (2015). «Передача сигналов через щелевые соединения на большие расстояния контролирует онкогенно-опосредованный онкогенез у эмбрионов Xenopus laevis» . Границы в физиологии . 5 : 519. doi : 10.3389/fphys.2014.00519 . ПМК 4298169 . ПМИД 25646081 .
- ^ Jump up to: а б Чернет, Брук Т; Левин, Майкл (2014). «Трансмембранный потенциал соматических клеток контролирует опосредованный онкогенами онкогенез на больших расстояниях» . Онкотаргет . 5 (10): 3287–306. doi : 10.18632/oncotarget.1935 . ПМК 4102810 . ПМИД 24830454 .
- ^ Jump up to: а б Юн, Джи; Оберой, С; Тристани-Фирузи, М; Этеридж, СП; Китания, Л; Крамер, Дж. Х.; Миллер, БЛ; Фу, Ю.Х.; Птачек, ЖЖ (2006). «Синдром Андерсена-Тавиля: проспективный когортный анализ и расширение фенотипа». Американский журнал медицинской генетики, часть A. 140А (4): 312–321. дои : 10.1002/ajmg.a.31092 . ПМИД 16419128 . S2CID 33899188 .
- ^ Пластер, Никки М; Тавил, Лави; Тристани-Фирузи, Мартин; Канун, Соня; Бендаху, Саид; Цунода, Акико; Дональдсон, Мэтью Р.; Ианнакконе, Сьюзан Т; Брант, Юаут; Барон, Ричард; Кларк, Джон; Деймер, Феза; Джордж, Альфред Л.; Фиш, Фрэнк А; Хан, Анжелика; Ниту, Александру; Оздемир, Джошкун; Сердароглу, Пирайе; Субрамони, Ш.Х.; Вулф, Гил; Фу, Ин-Хуэй; Птачек, Луи Дж (2001). «Мутации в Kir2.1 вызывают развитие и эпизодические электрические фенотипы синдрома Андерсена» . Клетка . 105 (4): 511–519. дои : 10.1016/S0092-8674(01)00342-7 . ПМИД 11371347 . S2CID 17015195 .
- ^ Jump up to: а б Масотти, Андреа; Ува, Паоло; Дэвис-Кеппен, Лаура; Базель-Ванагайте, Лина; Коэн, Лиор; Писанески, Элиза; Селлуцци, Антонелла; Бенчивенга, Паола; Фанг, Мингьян; Тиан, Мингю; Сюй, Сюнь; Каппа, Марко; Даллапиккола, Бруно (2015). «Синдром Кеппена-Любинского вызван мутациями во внутреннем выпрямляющем канале K+, кодируемом KCNJ6» . Американский журнал генетики человека . 96 (2): 295–300. дои : 10.1016/j.ajhg.2014.12.011 . ПМК 4320262 . ПМИД 25620207 .
- ^ Папулидис, И.; Папагеоргиу, Э.; Сиому, Э.; и др. (2014). «Пациент с частичной трисомией 21 и делецией 7q демонстрирует легкий фенотип синдрома Дауна». Джин . 536 (2): 441–443. дои : 10.1016/j.gene.2013.11.078 . ПМИД 24334122 .
- ^ Вальо, Стефано (2010). «Изменчивые сигналы во время беременности». Феромоны . Витамины и гормоны. Том. 83. С. 289–304. дои : 10.1016/S0083-6729(10)83012-2 . ISBN 978-0-12-381516-3 . ПМИД 20831951 .
- ^ Ямамото, Тецуо; Киносита, Манабу; Шиномия, Нариёси; и др. (2010). «Предварительная обработка аскорбиновой кислотой предотвращает летальный желудочно-кишечный синдром у мышей, получивших огромное количество радиации» . Журнал радиационных исследований . 51 (2): 145–156. Бибкод : 2010JRadR..51..145Y . дои : 10.1269/jrr.09078 . ПМИД 19959877 .
- ^ Чапкова, Павлина; Мисовикова, Надежда; Врбицка, Дита (2013). «Частичная трисомия и тетрасомия хромосомы 21 без фенотипа синдрома Дауна и краткий обзор корреляции генотип-фенотип. Отчет о случае» . Биомедицинские статьи . 158 (2): 321–325. дои : 10.5507/bp.2013.077 . ПМИД 24145769 .
- ^ Мегарбане, Андре; Аль-Али, Рашид; Шукайр, Нэнси; и др. (2016). «Синдром Темпла-Барайтсера и синдром Циммермана-Лабанда: одна клиническая сущность?» . BMC Медицинская генетика . 17 (1): 42. дои : 10.1186/s12881-016-0304-4 . ПМК 4901505 . ПМИД 27282200 .
- ^ Мастранжело, М.; Шеффер, И.Е.; Брамсвиг, Северная Каролина; Наир, LD; Майерс, Коннектикут; Дентичи, М.Л.; Коренке, Г.К.; Шох, К; Кампо, премьер-министр; Уайт, С.М.; Шаши, В; Кансагра, С; Ван Эссен, AJ; Леуцци, В. (2016). «Эпилепсия при синдромах, связанных с KCNH1». Эпилептические расстройства . 18 (2): 123–136. дои : 10.1684/epd.2016.0830 . ПМИД 27267311 .
- ^ Брамсвиг, Нурия С; Окелоен, CW; Чещик, Дж. К.; Ван Эссен, AJ; Пфундт, Р; Смитинк, Дж; Опрос-The, BT; Энгельс, Х; Стром, Т.М; Вечорек, Д; Клифстра, Т; Людеке, Х.-Ю (2015). « Расщепление против объединения в кучу: синдромы Темпла-Барайцера и Циммермана-Лабанда». Генетика человека . 134 (10): 1089–1097. дои : 10.1007/s00439-015-1590-1 . ПМИД 26264464 . S2CID 14238362 .
- ^ Jump up to: а б Кортюм, Фанни; Капуто, Вивиана; Бауэр, Кристиана К; и др. (2015). «Мутации в KCNH1 и ATP6V1B2 вызывают синдром Циммермана-Лабанда». Природная генетика . 47 (6): 661–7. дои : 10.1038/ng.3282 . hdl : 2108/118197 . ПМИД 25915598 . S2CID 12060592 .
- ^ Кастори, Марко; Морлино, Сильвия; Рителли, Марко; и др. (2014). «Поздняя диагностика синдрома латерального менингоцеле у женщины 55 лет с симптомами нестабильности суставов и хронической скелетно-мышечной болью». Американский журнал медицинской генетики, часть A. 164 (2): 528–534. дои : 10.1002/ajmg.a.36301 . ПМИД 24311540 . S2CID 12063113 .
- ^ Перкс, Т; Попат, Х.; Кронин, AJ; Дёрнинг, П; Мэггс, Р. (2013). «Ортодонтическое и хирургическое лечение синдрома Циммермана-Лабанда». Ортодонтия . 14 (1): e168–176. дои : 10.11607/ortho.897 . ПМИД 23646327 .
- ^ Саваки, К.; Мисима, К.; Сато, А.; и др. (2012). «Синдром Циммермана-Лабанда». Журнал клинической детской стоматологии . 36 (3): 297–300. дои : 10.17796/jcpd.36.3.k854128176u764l8 . ПМИД 22838235 .
- ^ Дуфендач, Калифорния; Джудисесси, младший; Бочек, Нью-Джерси; Акерман, MJ (2013). «Материнский мозаицизм затрудняет неонатальную диагностику синдрома Тимоти 1 типа» . Педиатрия . 131 (6): e1991–1995. дои : 10.1542/пед.2012-2941 . ПМК 3666110 . ПМИД 23690510 .
- ^ Сплавский, Игорь; Тимоти, Кэтрин В.; Шарп, Лия М; и др. (2004). «Дисфункция кальциевых каналов CaV1.2 вызывает мультисистемное расстройство, включая аритмию и аутизм» . Клетка . 119 (1): 19–31. дои : 10.1016/j.cell.2004.09.011 . ПМИД 15454078 . S2CID 15325633 .
- ^ Маргулис, Андреа В.; Митчелл, Аллен А.; Гильбоа, Сюзанна М.; Верлер, Марта М.; Миттлман, Мюррей А; Глинн, Роберт Дж.; Эрнандес-Диас, Соня (2012). «Использование топирамата во время беременности и риск расщелины рта» . Американский журнал акушерства и гинекологии . 207 (5): 405.e1–7. дои : 10.1016/j.ajog.2012.07.008 . ПМЦ 3484193 . ПМИД 22917484 .
- ^ Хилл, Дениз С.; Влодарчик, Богдан Ю.; Паласиос, Ана М.; Финнелл, Ричард Х. (2014). «Тератогенное действие противоэпилептических препаратов» . Экспертный обзор нейротерапии . 10 (6): 943–959. дои : 10.1586/ern.10.57 . ПМЦ 2970517 . ПМИД 20518610 .
- ^ Уайт, Х. Стив; Смит, Мисти Д.; Уилкокс, Карен С. (2007). «Механизмы действия противоэпилептических препаратов» . Нейробиология эпилепсии и старения . Международное обозрение нейробиологии. Том. 81. С. 85–110 . дои : 10.1016/S0074-7742(06)81006-8 . ISBN 978-0-12-374018-2 . ПМИД 17433919 .
- ^ Фриц, Х.; Мюллер, Д.; Хесс, Р. (1976). «Сравнительное исследование тератогенности фенобарбитона, дифенгидатоина и карбамазепина на мышах». Токсикология . 6 (3): 323–330. дои : 10.1016/0300-483X(76)90036-6 . ПМИД 996878 .
- ^ Фельдман, Джеральд Л.; Уивер, Д.Д.; Ловриен, EW (1977). «Фетальный триметадионовый синдром». Американский журнал болезней детей . 131 (12): 1389–1392. дои : 10.1001/archpedi.1977.02120250071012 . ПМИД 412416 .
- ^ Jump up to: а б Баркер, AT; Яффе, LF; Ванейбл, JW (1982). «Голый эпидермис кариеса содержит мощную батарею». Американский журнал физиологии. Регуляторная, интегративная и сравнительная физиология . 242 (3): Р358–366. дои : 10.1152/ajpregu.1982.242.3.R358 . ПМИД 7065232 .
- ^ Jump up to: а б Блю, О; Скотт, БиГ (1950). «Вибрационный зондовый электрометр для измерения биоэлектрических потенциалов». Обзор научных инструментов . 21 (10): 867–868. Бибкод : 1950RScI...21..867B . дои : 10.1063/1.1745444 . ПМИД 14786543 .
- ^ Чан, Мэйчэн; Робинсон, Кеннет Р.; Ванейбл, Джозеф В. (1992). «Электрические поля вблизи эпителиальных ран изолированного бычьего глаза». Экспериментальное исследование глаз . 54 (6): 999–1003. дои : 10.1016/0014-4835(92)90164-Н . ПМИД 1521590 .
- ^ Чан, Мэйчэн; Краго, Эдвард Дж; Ванейбл, Джозеф В. (1991). «Собственные электрические поля способствуют эпителизации ран у тритона Notophthalmus viridescens». Биология развития . 146 (2): 377–385. дои : 10.1016/0012-1606(91)90239-Y . ПМИД 1864462 .
- ^ Jump up to: а б Рид, Брайан; Сонг, Бинг; Маккейг, Колин Д.; Чжао, Мин (2005). «Заживление ран роговицы крыс: роль электрического тока» . Журнал ФАСЭБ . 19 (3): 379–386. дои : 10.1096/fj.04-2325com . ПМЦ 1459277 . ПМИД 15746181 .
- ^ Jump up to: а б с д Чжао, Мин; Сонг, Бинг; Пу, Джин; и др. (2006). «Электрические сигналы контролируют заживление ран посредством фосфатидилинозитол-3-ОН-киназы-γ и PTEN». Природа . 442 (7101): 457–460. Бибкод : 2006Natur.442..457Z . дои : 10.1038/nature04925 . ПМИД 16871217 . S2CID 4391475 .
- ^ Шен, Юньюнь; Пфлюгер, Триша; Феррейра, Фернандо; Лян, Цзебин; Наведо, Мануэль Ф; Цзэн, Цюньли; Рид, Брайан; Чжао, Мин (2016). «Диабетические раны роговицы производят значительно более слабые электрические сигналы, что может способствовать ухудшению заживления» . Научные отчеты . 6 : 26525. Бибкод : 2016NatSR...626525S . дои : 10.1038/srep26525 . ПМК 4901296 . ПМИД 27283241 .
- ^ Морис, Д.М. Проницаемость для ионов натрия роговицы живого кролика. J Physiol 112, 367–391. Опубликованный центральный ссылочный номер: PMC1393020.
- ^ Клайс, С.Д. Электрические профили эпителия роговицы. J Physiol 226, 407-429. Опубликованный центральный справочный номер: PMC1331188.
- ^ Песня, Б (2004). «Регенерация нервов и заживление ран стимулируются и направляются эндогенным электрическим полем in vivo» . Журнал клеточной науки . 117 (20): 4681–4690. дои : 10.1242/jcs.01341 . ПМИД 15371524 .
- ^ Лин, Ф.; Балдессари, Ф.; Дьенге, CC; и др. (2008). «Электротаксис лимфоцитов in vitro и in vivo» . Журнал иммунологии . 181 (4): 2465–2471. дои : 10.4049/jimmunol.181.4.2465 . ПМЦ 2572691 . ПМИД 18684937 .
- ^ Ян, Х.-й; Чарльз, Р.-П; Хаммлер, Э; Бэйнс, Д.Л.; Иссеров, Р.Р. (2013). «Эпителиальный натриевый канал обеспечивает направленность гальванотаксиса в кератиноцитах человека» . Журнал клеточной науки . 126 (9): 1942–1951. дои : 10.1242/jcs.113225 . ПМЦ 3666251 . ПМИД 23447677 .
- ^ Аллен, Грег М.; Могильнер, Алекс; Териот, Джули А. (2013). «Электрофорез компонентов клеточных мембран создает сигнал направления, управляющий гальванотаксисом кератоцитов» . Современная биология . 23 (7): 560–568. Бибкод : 2013CBio...23..560A . дои : 10.1016/j.cub.2013.02.047 . ПМЦ 3718648 . ПМИД 23541731 .
- ^ Чанг, Фред; Минк, Николас (2014). «Электрохимический контроль полярности клеток и тканей» . Ежегодный обзор клеточной биологии и биологии развития . 30 : 317–336. doi : 10.1146/annurev-cellbio-100913-013357 . ПМИД 25062359 .
- ^ Робинсон, КР (1985). «Реакция клеток на электрические поля: обзор» . Журнал клеточной биологии . 101 (6): 2023–2037. дои : 10.1083/jcb.101.6.2023 . ПМК 2114002 . ПМИД 3905820 .
- ^ Нисимура, Кентукки; Иссеров, Р.Р.; Нуччителли, Р. (1996). «Человеческие кератиноциты мигрируют к отрицательному полюсу в электрических полях постоянного тока, сравнимых с теми, которые измеряются в ранах млекопитающих» . Журнал клеточной науки . 109 (1): 199–207. дои : 10.1242/jcs.109.1.199 . ПМИД 8834804 .
- ^ Чжао, М.; Агиус-Фернандес, А.; Форрестер, СП; Маккейг, компакт-диск (1996). «Ориентация и направленная миграция культивируемых эпителиальных клеток роговицы в небольших электрических полях зависят от сыворотки» . Журнал клеточной науки . 109 (6): 1405–1414. дои : 10.1242/jcs.109.6.1405 . ПМИД 8799828 .
- ^ Грулер, Ганс; Нуччителли, Ричард (2000). «Механизм реакции гальванотаксиса кератиноцитов можно смоделировать как пропорциональный контроллер». Клеточная биохимия и биофизика . 33 (1): 33–51. дои : 10.1385/CBB:33:1:33 . ПМИД 11322511 . S2CID 11731666 .
- ^ Чжао, М; Агиус-Фернандес, А; Форрестер, СП; Маккейг, CD (1996). «Направленная миграция эпителиальных листов роговицы в физиологических электрических полях» . Исследовательская офтальмология и визуальные науки . 37 (13): 2548–2558. ПМИД 8977469 .
- ^ Накадзима, Кен-Ичи; Чжу, Кан; Сунь, Яо-Хуэй; и др. (2015). «KCNJ15/Kir4.2 соединяется с полиаминами, чувствуя слабые внеклеточные электрические поля при гальванотаксисе» . Природные коммуникации . 6 : 8532. Бибкод : 2015NatCo...6.8532N . дои : 10.1038/ncomms9532 . ПМЦ 4603535 . ПМИД 26449415 .
- ^ Гао, Ранчи; Чжао, Сивэй; Цзян, Сюпин; и др. (2015). «Крупномасштабный скрининг выявил гены, которые опосредуют электротаксис у Dictyostelium discoideum » . Научная сигнализация . 8 (378): ра50. doi : 10.1126/scisignal.aab0562 . ПМЦ 4470479 . ПМИД 26012633 .
- ^ Джамгоз, МБ А; Мисельска, М; Мадея, З; и др. (2001). «Направленное движение клеток рака предстательной железы крыс в электрическом поле постоянного тока: участие активности потенциалзависимых Na+-каналов» . Журнал клеточной науки . 114 (14): 2697–2705. дои : 10.1242/jcs.114.14.2697 . ПМИД 11683396 .
- ^ Чжан, Гаофэн; Эдмундсон, Мэтью; Тележкин, Всеволод; и др. (2016). «Роль канала Kv1.2 в миграции клеток электротаксиса» . Журнал клеточной физиологии . 231 (6): 1375–1384. дои : 10.1002/jcp.25259 . ПМЦ 4832312 . ПМИД 26580832 .
- ^ Чжан, Гаофэн; Гу, Ю; Бегум, Румена; и др. (2016). «Киндлин-1 регулирует электротаксис кератиноцитов» . Журнал исследовательской дерматологии . 136 (11): 2229–2239. дои : 10.1016/j.jid.2016.05.129 . ПМЦ 5756539 . ПМИД 27427485 .
- ^ Чжао, МИН; Пу, Цзинь; Форрестер, Джон В.; и др. (2002). «Мембранные липиды, рецепторы ЭФР и внутриклеточные сигналы колокализуются и поляризуются в эпителиальных клетках, движущихся направленно в физиологическом электрическом поле» . Журнал ФАСЭБ . 16 (8): 857–859. doi : 10.1096/fj.01-0811fje . ПМИД 11967227 . S2CID 31682478 .
- ^ Линь, Бо-Цзянь; Цао, Шунь-хао; Чен, Алекс; и др. (2017). «Липидные рафты чувствуют и направляют миграцию, вызванную электрическим полем» . Труды Национальной академии наук . 114 (32): 8568–8573. Бибкод : 2017PNAS..114.8568L . дои : 10.1073/pnas.1702526114 . ПМК 5559012 . ПМИД 28739955 .
- ^ Маден, М. (1991). История исследований регенерации . Кембриджский университет. [ нужна страница ]
- ^ Марш, Гордон; Бимс, HW (1952). «Электрический контроль морфогенеза у регенерирующей dugesia tigrina. I. Связь осевой полярности с силой поля». Журнал клеточной и сравнительной физиологии . 39 (2): 191–213. дои : 10.1002/jcp.1030390203 . ПМИД 14946235 .
- ^ Боргенс, Ричард Б. (1984). «Инициируются ли развитие конечностей и регенерация конечностей повреждением покровов?». Дифференциация . 28 (2): 87–93. дои : 10.1111/j.1432-0436.1984.tb00270.x . ПМИД 6526168 .
- ^ Ликкен, Дэвид Т. (1970). «Прямоугольный анализ импеданса кожи». Психофизиология . 7 (2): 262–275. дои : 10.1111/j.1469-8986.1970.tb02232.x . ПМИД 5499129 .
- ^ Смит, Стивен Д. (1967). «Индукция частичной регенерации конечностей у Rana pipiens путем гальванической стимуляции». Анатомическая запись . 158 (1): 89–97. дои : 10.1002/ar.1091580110 . ПМИД 6033441 . S2CID 22547794 .
- ^ Jump up to: а б Дженкинс, Лиза С; Дуэрсток, Брэдли С; Боргенс, Ричард Б. (1996). «Уменьшение тока травмы, выходящей за пределы ампутации, тормозит регенерацию конечностей у краснопятнистого тритона» . Биология развития . 178 (2): 251–262. дои : 10.1006/dbio.1996.0216 . ПМИД 8812127 .
- ^ Боргенс, Р.Б.; Ванейбл, Дж. В.; Яффе, Л.Ф. (1977). «Биоэлектричество и регенерация: сильные токи оставляют культи регенерирующих конечностей тритона» . Труды Национальной академии наук . 74 (10): 4528–32. Бибкод : 1977PNAS...74.4528B . дои : 10.1073/pnas.74.10.4528 . ПМК 431978 . ПМИД 270701 .
- ^ Jump up to: а б Боргенс, Ричард Б; Ванейбл, Джозеф В.; Яффе, Лайонел Ф (1979). «Небольшие искусственные токи улучшают регенерацию конечностей Xenopus ». Журнал экспериментальной зоологии . 207 (2): 217–226. Бибкод : 1979JEZ...207..217B . дои : 10.1002/jez.1402070206 .
- ^ Маккейг, CD «Электрические поля при восстановлении позвоночных». (Физиологическое общество, 1989).
- ^ Jump up to: а б Ясуда, Ивао (1974). «Механическая и электрическая мозоль». Анналы Нью-Йоркской академии наук . 238 : 457–465. дои : 10.1111/j.1749-6632.1974.tb26812.x . ПМИД 4531275 . S2CID 84676921 .
- ^ Фукада, Эйичи; Ясуда, Ивао (1957). «О пьезоэлектрическом эффекте кости». Журнал Физического общества Японии . 12 (10): 1158–1162. Бибкод : 1957JPSJ...12.1158F . дои : 10.1143/JPSJ.12.1158 .
- ^ Брюс М. Карлсон, доктор медицинских наук, доктор философии. Принципы регенеративной биологии. (Академическая пресса, 2007). [ нужна страница ]
- ^ Jump up to: а б Голдинг, Энн; Гуэй, Джастин А; Эррера-Ринкон, Селия; Левин, Майкл; Каплан, Дэвид Л. (2016). «Настраиваемое устройство из гидрогеля шелка для изучения регенерации конечностей у взрослых Xenopus Laevis» . ПЛОС ОДИН . 11 (6): e0155618. Бибкод : 2016PLoSO..1155618G . дои : 10.1371/journal.pone.0155618 . ПМЦ 4892606 . ПМИД 27257960 .
- ^ Jump up to: а б Хечаваррия, Даниэль; Девильде, Абиш; Браунхут, Сьюзен; Левин, Майкл; Каплан, Дэвид Л. (2010). «Регенеративная манжета BioDome для биохимической и биофизической стимуляции регенерации тканей» . Медицинская инженерия и физика . 32 (9): 1065–1073. дои : 10.1016/j.medengphy.2010.07.010 . ПМЦ 2967604 . ПМИД 20708956 .
- ^ Леппик, Людмила П; Фремель, Дара; Славичи, Андрей; Овадия, Закри Н; Худак, Лукаш; Генрих, Дирк; Марци, Инго; Баркер, Джон Х (2015). «Влияние электростимуляции на регенерацию конечностей крыс: новый взгляд на старую модель» . Научные отчеты . 5 : 18353. Бибкод : 2015NatSR...518353L . дои : 10.1038/srep18353 . ПМЦ 4683620 . ПМИД 26678416 .
- ^ Рид, Брайан; Сонг, Бинг; Чжао, Мин (2009). «Электрические токи при регенерации хвоста головастика Xenopus» . Биология развития . 335 (1): 198–207. дои : 10.1016/j.ydbio.2009.08.028 . ПМИД 19733557 .
- ^ Ценг, Айсун; Левин, Майкл (2014). «Взлом биоэлектрического кода: исследование эндогенного ионного контроля формирования паттернов» . Коммуникативная и интегративная биология . 6 (1): e22595. дои : 10.4161/cib.22595 . ПМЦ 3689572 . ПМИД 23802040 .
- ^ Jump up to: а б с Адамс, Д.С.; Маси, А; Левин, М. (2007). «H+-насос-зависимые изменения мембранного напряжения являются ранним механизмом, необходимым и достаточным для индукции регенерации хвоста Xenopus» . Разработка . 134 (7): 1323–1335. дои : 10.1242/dev.02812 . ПМИД 17329365 .
- ^ Jump up to: а б с д Ценг, А.-С; Бин, Вашингтон; Лемир, Дж. М.; Маси, А; Левин, М (2010). «Индукция регенерации позвоночных кратковременным натриевым током» . Журнал неврологии . 30 (39): 13192–13200. doi : 10.1523/JNEUROSCI.3315-10.2010 . ПМЦ 2965411 . ПМИД 20881138 .
- ^ Jump up to: а б с Адамс, Д.С.; Ценг, А.-С; Левин, М (2013). «Световая активация архаэрродопсинового H+-насоса обращает вспять возрастную потерю регенерации позвоночных: активный контроль на системном уровне in vivo» . Биология Открытая . 2 (3): 306–313. дои : 10.1242/bio.20133665 . ПМК 3603412 . ПМИД 23519324 .
- ^ Овьедо, Нью-Джерси; Левин, М. (2007). «Smedinx-11 — это ген щелевого соединения планарийных стволовых клеток, необходимый для регенерации и гомеостаза» . Разработка . 134 (17): 3121–3131. дои : 10.1242/dev.006635 . ПМИД 17670787 .
- ^ Бин, Вашингтон; Морокума, Дж; Лемир, Дж. М.; Левин, М (2012). «Биоэлектрическая передача сигналов регулирует размер головы и органов во время регенерации планарий» . Разработка . 140 (2): 313–322. дои : 10.1242/dev.086900 . ПМЦ 3597208 . ПМИД 23250205 .
- ^ Бин, Венди С; Морокума, Джунджи; Адамс, Дэни С; Левин, Майкл (2011). «Подход химической генетики показывает, что для регенерации головы планарий необходимо мембранное напряжение, опосредованное H,K-АТФазой» . Химия и биология . 18 (1): 77–89. doi : 10.1016/j.chembiol.2010.11.012 . ПМЦ 3278711 . ПМИД 21276941 .
- ^ Jump up to: а б Эммонс-Белл, Майя; Дюрант, Фэллон; Хаммельман, Дженнифер; Бессонов, Николай; Вольперт, Виталий; Морокума, Джунджи; Пинет, Кайлиннетт; Адамс, Дэни; Пиетак, Алексис; Лобо, Дэниел; Левин, Майкл (2015). «Блокада щелевого соединения стохастически индуцирует различную видоспецифичную анатомию головы у генетически диких плоских червей Girardia dorotocephala» . Международный журнал молекулярных наук . 16 (11): 27865–27896. дои : 10.3390/ijms161126065 . ПМЦ 4661923 . ПМИД 26610482 .
- ^ Ноги, Тайсаку; Левин, Майкл (2005). «Характеристика экспрессии гена иннексина и функциональной роли щелевых соединений в регенерации планарий» . Биология развития . 287 (2): 314–335. дои : 10.1016/j.ydbio.2005.09.002 . ПМИД 16243308 .
- ^ Овьедо, Нестор Х; Морокума, Джунджи; Валентек, Питер; Кема, Идо П; Гу, Ман Бок; Ан, Джу-Мён; Хван, Юнг Шан; Годобори, Такаши; Левин, Майкл (2010). «Дальнодействующие сигналы, опосредованные нейронами и белками щелевых соединений, контролируют полярность во время регенерации планарий» . Биология развития . 339 (1): 188–199. дои : 10.1016/j.ydbio.2009.12.012 . ПМЦ 2823934 . ПМИД 20026026 .
- ^ Jump up to: а б Дюрант, Фэллон; Морокума, Джунджи; Филдс, Кристофер; Уильямс, Кэтрин; Адамс, Дэни Спенсер; Левин, Майкл (2017). «Долгосрочное стохастическое редактирование регенеративной анатомии посредством нацеливания на эндогенные биоэлектрические градиенты» . Биофизический журнал . 112 (10): 2231–2243. Бибкод : 2017BpJ...112.2231D . дои : 10.1016/j.bpj.2017.04.011 . ПМЦ 5443973 . ПМИД 28538159 .
- ^ Нойхоф, Моран; Левин, Майкл; Рехави, Одед (2016). «Вертикально- и горизонтально передаваемые воспоминания – стирание границ между регенерацией и наследованием у планарий» . Биология Открытая . 5 (9): 1177–1188. дои : 10.1242/bio.020149 . ПМК 5051648 . ПМИД 27565761 .
- ^ Jump up to: а б Лобикин, Мария; Чернет, Брук; Лобо, Дэниел; Левин, Майкл (2012). «Потенциал покоя, индуцированный онкогенами онкогеногенез и метастазирование: биоэлектрическая основа рака in vivo» . Физическая биология . 9 (6): 065002. Бибкод : 2012PhBio...9f5002L . дои : 10.1088/1478-3975/9/6/065002 . ПМК 3528107 . ПМИД 23196890 .
- ^ Ян, Мин; Брекенбери, Уильям Дж. (2013). «Мембранный потенциал и прогрессирование рака» . Границы в физиологии . 4 : 185. дои : 10.3389/fphys.2013.00185 . ПМЦ 3713347 . ПМИД 23882223 .
- ^ Jump up to: а б Кандуз, Мустафа; Батист, Джеральд (2010). «Щелевые соединения и коннексины как терапевтические мишени при раке». Мнение экспертов о терапевтических целях . 14 (7): 681–692. дои : 10.1517/14728222.2010.487866 . ПМИД 20446866 . S2CID 30844116 .
- ^ Лейт, Эдвард; Сирнес, Сольвейг; Омори, Ясуфуми; Риведал, Эдгар (2006). «Понижение уровня щелевых соединений в раковых клетках». Критические обзоры онкогенеза . 12 (3–4): 225–256. дои : 10.1615/CritRevOncog.v12.i3-4.30 . ПМИД 17425504 .
- ^ Троско, Дж. Э. (2005). «Роль стволовых клеток и щелевых соединений как мишеней для химиопрофилактики рака и химиотерапии». Биомедицина и фармакотерапия . 59 : С326–331. дои : 10.1016/S0753-3322(05)80065-4 . ПМИД 16507402 .
- ^ Пардо, Луис А; Штюмер, Вальтер (2013). «Роль K+-каналов при раке». Обзоры природы Рак . 14 (1): 39–48. дои : 10.1038/nrc3635 . ПМИД 24336491 . S2CID 28497543 .
- ^ Хуан, Си; Ян, Лили Йе (2014). «Нацеливание на калиевые каналы при раке» . Журнал клеточной биологии . 206 (2): 151–162. дои : 10.1083/jcb.201404136 . ПМК 4107787 . ПМИД 25049269 .
- ^ Арканджели, Аннароса; Беккетти, Андреа (2010). «Новые тенденции в терапии рака: воздействие на ионные каналы и транспортеры» . Фармацевтика . 3 (4): 1202–1224. дои : 10.3390/ph3041202 . ПМК 4034029 . ПМИД 27713296 .
- ^ Фрейзер, С.П.; Озерлат-Гюндуз, I; Брэкенбери, WJ; Фицджеральд, Э.М.; Кэмпбелл, Т.М; Кумбс, Р.К.; Джамгоз, МБ А (2014). «Регуляция экспрессии потенциалзависимых натриевых каналов при раке: гормоны, факторы роста и ауторегуляция» . Философские труды Королевского общества B: Биологические науки . 369 : 20130105.doi : (1638 ) 10.1098/rstb.2013.0105 . ПМЦ 3917359 . ПМИД 24493753 .
- ^ Джамгоз, МБ А; Кумбс, Р.К.; Шваб, А (2014). «Ионный транспорт и рак: от инициации до метастазирования» . Философские труды Королевского общества B: Биологические науки . 369 (1638): 20130092. doi : 10.1098/rstb.2013.0092 . ПМЦ 3917347 . ПМИД 24493741 .
- ^ Фреде, Джулия; Фрейзер, Скотт П.; Оскай-Озчелик, Гюльтен; Хон, Ёсон; Иоана Браику, Э; Сехули, Джалид; Габра, Хани; Джамгоз, Мустафа Б.А. (2013). «Рак яичников: ионный канал и экспрессия аквапоринов как новые мишени клинического потенциала». Европейский журнал рака . 49 (10): 2331–2344. дои : 10.1016/j.ejca.2013.03.016 . ПМИД 23683551 .
- ^ Йылдирим, Сенай; Алтун, Сейхан; Гумушан, Хатидже; Патель, Ануп; Джамгоз, Мустафа Б.А. (2012). «Активность потенциалзависимых натриевых каналов способствует метастазированию рака простаты in vivo». Письма о раке . 323 (1): 58–61. дои : 10.1016/j.canlet.2012.03.036 . ПМИД 22484465 .
- ^ Блэкистон, Д; Адамс, Д.С.; Лемир, Дж. М.; Лобикин М.; Левин, М (2010). «Трансмембранный потенциал инструкторских клеток, экспрессирующих Gly Cl, индуцирует неопластическую конверсию меланоцитов по серотонинергическому пути» . Модели и механизмы заболеваний . 4 (1): 67–85. дои : 10.1242/dmm.005561 . ПМК 3008964 . ПМИД 20959630 .
- ^ Морокума, Дж; Блэкистон, Д; Адамс, Д.С.; Сибом, Г; Триммер, Б; Левин, М (2008). «Модуляция функции калиевых каналов придает эмбриональным стволовым клеткам гиперпролиферативный инвазивный фенотип» . Труды Национальной академии наук . 105 (43): 16608–13. Бибкод : 2008PNAS..10516608M . дои : 10.1073/pnas.0808328105 . JSTOR 25465142 . ПМЦ 2575467 . ПМИД 18931301 .
- ^ Jump up to: а б Чернет, Брук Т; Адамс, Дэни С; Лобикин, Мария; Левин, Майкл (2016). «Использование генетически закодированных светозависимых ионных транслокаторов для контроля онкогенеза» . Онкотаргет . 7 (15): 19575–19588. дои : 10.18632/oncotarget.8036 . ПМЦ 4991402 . ПМИД 26988909 .
- ^ Чернет, Б.Т; Левин, М (2013). «Потенциал трансмембранного напряжения является важным клеточным параметром для обнаружения и контроля развития опухоли в модели Xenopus» . Модели и механизмы заболеваний . 6 (3): 595–607. дои : 10.1242/dmm.010835 . ПМЦ 3634644 . ПМИД 23471912 .
- ^ Ли, Чунмей; Левин, Майкл; Каплан, Дэвид Л. (2016). «Биоэлектрическая модуляция поляризации макрофагов» . Научные отчеты . 6 : 21044. Бибкод : 2016NatSR...621044L . дои : 10.1038/srep21044 . ПМЦ 4751571 . ПМИД 26869018 .
- ^ Озкукур, Нурдан; Куинн, Кайл П.; Панг, Джин С; Ду, Чуан; Георгакуди, Ирен; Миллер, Эрик; Левин, Майкл; Каплан, Дэвид Л. (2015). «Деполяризация мембранного потенциала вызывает изменения в расположении и связности нейронов в совместных культурах» . Мозг и поведение . 5 (1): 24–38. дои : 10.1002/brb3.295 . ПМЦ 4321392 . ПМИД 25722947 .
- ^ Лобикин, Мария; Паре, Жан-Франсуа; Каплан, Дэвид Л; Левин, Майкл (2015). «Селективная деполяризация трансмембранного потенциала изменяет структуру мышц и локализацию мышечных клеток у эмбрионов Xenopus laevis» . Международный журнал биологии развития . 59 (7–8–9): 303–311. дои : 10.1387/ijdb.150198ml . ПМЦ 10461602 . ПМИД 26198143 .
- ^ Сунделакруз, Сара; Ли, Чунмей; Чхве, Ён Джун; Левин, Майкл; Каплан, Дэвид Л. (2013). «Биоэлектрическая модуляция заживления ран в 3D-модели тканеинженерной кости in vitro» . Биоматериалы . 34 (28): 6695–6705. doi : 10.1016/j.bimaterials.2013.05.040 . ПМЦ 3724996 . ПМИД 23764116 .
- ^ Сунделакруз, Сара; Левин, Майкл; Каплан, Дэвид Л. (2013). «Деполяризация изменяет фенотип, поддерживает пластичность предифференцированных мезенхимальных стволовых клеток» . Тканевая инженерия, часть А. 19 (17–18): 1889–1908. дои : 10.1089/ten.tea.2012.0425.rev . ПМЦ 3726227 . ПМИД 23738690 .
- ^ Хинард, В.; Белин, Д; Кениг, С; Бадер, CR; Бернхейм, Л. (2008). «Инициация дифференцировки миобластов человека посредством дефосфорилирования каналов Kir2.1 K+ по тирозину 242» . Разработка . 135 (5): 859–867. дои : 10.1242/dev.011387 . ПМИД 18216177 .
- ^ Левин, Майкл (2012). «Молекулярное биоэлектричество в биологии развития: новые инструменты и недавние открытия» . Биоэссе . 34 (3): 205–217. дои : 10.1002/bies.201100136 . ПМК 3430077 . ПМИД 22237730 .
- ^ Левин, Майкл (2013). «Перепрограммирование клеток и структурирования тканей с помощью биоэлектрических путей: молекулярные механизмы и биомедицинские возможности» . Междисциплинарные обзоры Wiley: системная биология и медицина . 5 (6): 657–676. дои : 10.1002/wsbm.1236 . ПМЦ 3841289 . ПМИД 23897652 .
- ^ Мэтьюз, Хуанита; Левин, Майкл (2017). «Передача сигналов через щелевые соединения в регуляции паттернов: связь физиологических сетей управляет ростом и формой» . Развивающая нейробиология . 77 (5): 643–673. дои : 10.1002/dneu.22405 . ПМЦ 10478170 . ПМИД 27265625 .
- ^ Ценг, Ай-Сун; Левин, Майкл (2012). «Преобразование биоэлектрических сигналов в эпигенетические пути во время регенерации хвоста головастика» . Анатомическая запись . 295 (10): 1541–1451. дои : 10.1002/ar.22495 . ПМЦ 3442154 . ПМИД 22933452 .
- ^ Левин, Майкл (2007). «Крупномасштабная биофизика: потоки ионов и регенерация». Тенденции в клеточной биологии . 17 (6): 261–270. дои : 10.1016/j.tcb.2007.04.007 . ПМИД 17498955 .
- ^ Кнопфель, Т; Лин, М.З.; Левская, А; Тиан, Л; Лин, Дж. Ю.; Бойден, ES (2010). «На пути ко второму поколению оптогенетических инструментов» . Журнал неврологии . 30 (45): 14998–5004. doi : 10.1523/JNEUROSCI.4190-10.2010 . ПМЦ 2997431 . ПМИД 21068304 .
- ^ Фенно, Лиф; Йижар, Офер; Дейсерот, Карл (2011). «Развитие и применение оптогенетики» . Ежегодный обзор неврологии . 34 : 389–412. doi : 10.1146/annurev-neuro-061010-113817 . ПМК 6699620 . ПМИД 21692661 .
- ^ Лонг, Сяоян; Йе, Цзин; Чжао, Ди; Чжан, Шэн-Цзя (2015). «Магнитогенетика: Дистанционная неинвазивная магнитная активация активности нейронов с помощью магниторецептора» . Научный вестник . 60 (24): 2107–2119. Бибкод : 2015SciBu..60.2107L . дои : 10.1007/s11434-015-0902-0 . ПМК 4692962 . ПМИД 26740890 .
- ^ Уилсон, Максвелл З.; Равиндран, Павитран Т; Лим, Венделл А; Тетчер, Джаред Э. (2017). «Отслеживание потока информации от Erk до индукции целевого гена раскрывает механизмы динамического и комбинаторного контроля» . Молекулярная клетка . 67 (5): 757–769.e5. doi : 10.1016/j.molcel.2017.07.016 . ПМК 5591080 . ПМИД 28826673 .
- ^ Бугай, Лукаш Дж; о'Донохью, Джефф П.; Лим, Венделл А. (2017). «Опрос клеточного восприятия и принятия решений с помощью оптогенетических инструментов» . Журнал клеточной биологии . 216 (1): 25–28. дои : 10.1083/jcb.201612094 . ПМЦ 5223619 . ПМИД 28003330 .
- ^ Митчелл, Амир; Лим, Венделл (2016). «Клеточное восприятие и неправильное восприятие: внутренние модели принятия решений, сформированные эволюционным опытом» . Биоэссе . 38 (9): 845–849. doi : 10.1002/bies.201600090 . ПМЦ 4996742 . ПМИД 27461864 .
- ^ Фишбах, Массачусетс; Блюстоун, Дж. А.; Лим, Вашингтон (2013). «Клеточная терапия: следующий столп медицины» . Наука трансляционной медицины . 5 (179): 179пс7. doi : 10.1126/scitranslmed.3005568 . ПМЦ 3772767 . ПМИД 23552369 .
- ^ Чау, Анджела Х; Уолтер, Джессика М; Жерарден, Жалин; Тан, Чао; Лим, Венделл А. (2012). «Проектирование синтетических регуляторных сетей, способных к самоорганизации поляризации клеток» . Клетка . 151 (2): 320–332. дои : 10.1016/j.cell.2012.08.040 . ПМЦ 3498761 . ПМИД 23039994 .
- ^ Башор, Калеб Дж; Хорвиц, Эндрю А; Пейсайович, Серджио Г; Лим, Венделл А. (2010). «Перемонтирование клеток: синтетическая биология как инструмент исследования принципов организации живых систем» . Ежегодный обзор биофизики . 39 : 515–37. doi : 10.1146/annurev.biophys.050708.133652 . ПМЦ 2965450 . ПМИД 20192780 .
- ^ Пеццуло, Джованни; Левин, Майкл (2016). «Нисходящие модели в биологии: объяснение и контроль сложных живых систем выше молекулярного уровня» . Журнал интерфейса Королевского общества . 13 (124): 20160555. doi : 10.1098/rsif.2016.0555 . ПМК 5134011 . ПМИД 27807271 .
- ^ Jump up to: а б Пеццуло, Дж; Левин, М (2015). «Воспоминание о теле: применение вычислительной нейробиологии для нисходящего контроля регенерации конечностей и других сложных органов» . Интегративная биология . 7 (12): 1487–1517. дои : 10.1039/c5ib00221d . ПМЦ 4667987 . ПМИД 26571046 .
- ^ Фристон, К; Левин, М; Сенгупта, Б; Пеццуло, Дж. (2015). «Знать свое место: подход свободной энергии к регулированию моделей» . Журнал интерфейса Королевского общества . 12 (105): 20141383. doi : 10.1098/rsif.2014.1383 . ПМЦ 4387527 . ПМИД 25788538 .
- ^ Левин, Майкл (2014). «Эндогенные биоэлектрические сети хранят негенетическую информацию о паттернах во время развития и регенерации» . Журнал физиологии . 592 (11): 2295–2305. дои : 10.1113/jphysicalol.2014.271940 . ПМК 4048089 . ПМИД 24882814 .
- ^ Макнамара, Гарольд М; Чжан, Гонконг; Верли, Кристофер А; Коэн, Адам Э (2016). «Оптически управляемые генераторы в инженерной биоэлектрической ткани» . Физический обзор X . 6 (3): 031001. Бибкод : 2016PhRvX...6c1001M . дои : 10.1103/PhysRevX.6.031001 .
- ^ Ригас, С; Дебросы, Г; Харалампидис, К; Висенте-Агулло, нападающий; Фельдманн, К.А.; Грабов А; Долан, Л; Хацопулос, П. (2001). «TRH1 кодирует транспортер калия, необходимый для роста кончиков корневых волосков арабидопсиса» . Растительная клетка . 13 (1): 139–151. дои : 10.1105/tpc.13.1.139 . ПМЦ 102205 . ПМИД 11158535 .
- ^ Jump up to: а б Дахал, Г.Р.; Роусон, Дж; Гассауэй, Б; Квок, Б; Тонг, Ю; Птачек, Л.Дж.; Бейтс, Э. (2012). «Для формирования паттерна необходим внутренний выпрямляющий канал K+» . Разработка . 139 (19): 3653–3664. дои : 10.1242/dev.078592 . ПМЦ 3436115 . ПМИД 22949619 .
- ^ Вильянуэва, С; Бургос, Дж.; Лопес-Каюкео, К.И.; и др. (2015). «Расщелина неба, умеренная задержка развития легких и ранняя послеродовая летальность у мышей с дефицитом канала Kir7.1, внутренне исправляющего K+» . ПЛОС ОДИН . 10 (9): e0139284. Бибкод : 2015PLoSO..1039284V . дои : 10.1371/journal.pone.0139284 . ПМК 4581704 . ПМИД 26402555 .
- ^ Саймонс, М; Голт, штат Вашингтон; Готхардт, Д.; и др. (2009). «Электрохимические сигналы регулируют сборку комплекса «Вьющиеся/Растрепанные» на плазматической мембране во время плоской эпителиальной поляризации» . Природная клеточная биология . 11 (3): 286–294. дои : 10.1038/ncb1836 . ПМК 2803043 . ПМИД 19234454 .
- ^ Гермле, Т; Салтукоглу, Д.; Грюневальд, Дж.; и др. (2010). «Регуляция передачи сигналов планарной полярности, зависимой от Frizzled, с помощью субъединицы V-АТФазы» . Современная биология . 20 (14): 1269–1276. Бибкод : 2010CBio...20.1269H . дои : 10.1016/j.cub.2010.05.057 . ПМИД 20579879 . S2CID 15407237 .
- ^ Мюллер, К.; Маэзо, я; Витбродт, Дж.; Мартинес-Моралес-младший (2013). «Мутация тинтахины медаки проливает свет на эволюцию субъединиц V-АТФазы B у позвоночных» . Научные отчеты . 3 : 3217. Бибкод : 2013NatSR...3E3217M . дои : 10.1038/srep03217 . ПМЦ 3827601 . ПМИД 24225653 .
- ^ Бортвик, Кей Джей; Кандемир, Н.; Топалоглу, Р.; и др. (2003). «Фенокопия дефицита CAII: новое генетическое объяснение наследственного детского остеопетроза с дистальным почечным тубулярным ацидозом» . Журнал медицинской генетики . 40 (2): 115–121. дои : 10.1136/jmg.40.2.115 . ПМЦ 1735376 . ПМИД 12566520 .
- ^ Олдрич, Ричард В. (2015). «Новый стандарт: обзор Справочника по ионным каналам» . Журнал общей физиологии . 146 (2): 119–121. дои : 10.1085/jgp.201511461 . ПМЦ 4516783 . ПМИД 26216856 .
- ^ Дуке, А.; Газула, ВР; Качмарек, Л.К. (2013). «Экспрессия калиевых каналов Kv1.3 регулирует плотность корковых интернейронов» . Развивающая нейробиология . 73 (11): 841–855. дои : 10.1002/dneu.22105 . ПМЦ 3829632 . ПМИД 23821603 .
- ^ Чжэн, Дж. А. Т., М.С. Справочник по ионным каналам. (ЦРК Пресс, 2015). [ нужна страница ]
- ^ Кристенсен, А.Х; Шатлен, ФК; Хаттнер, И.Г.; и др. (2016). «Двухпоровый калиевый канал, TWIK-1, играет роль в регуляции частоты сердечных сокращений и размера предсердий». Журнал молекулярной и клеточной кардиологии . 97 : 24–35. дои : 10.1016/j.yjmcc.2016.04.006 . ПМИД 27103460 .
- ^ Саймонс, К; Раш, Л.Д.; Кроуфорд, Дж; и др. (2015). «Мутации в гене потенциалзависимого калиевого канала KCNH1 вызывают синдром Темпла-Барайтсера и эпилепсию». Природная генетика . 47 (1): 73–77. дои : 10.1038/ng.3153 . ПМИД 25420144 . S2CID 52799681 .
- ^ Лабонн, доктор медицинских наук; Грейвс, Т.Д.; Шен, Ю.; и др. (2016). «Микроделеция Xq22.2 связана с глициновым рецептором GLRA4, связанным с умственной отсталостью, поведенческими проблемами и черепно-лицевыми аномалиями» . БМК Неврология . 16 : 132. дои : 10.1186/s12883-016-0642-z . ПМЦ 4979147 . ПМИД 27506666 .
- ^ Хираки, Ю.; Миятаке, С.; Хаяшидани, М.; и др. (2014). «Аневризма аорты и краниосиностоз в семье с синдромом Канту». Американский журнал медицинской генетики, часть A. 164А (1): 231–236. дои : 10.1002/ajmg.a.36228 . ПМИД 24352916 . S2CID 73121 .
- ^ Купер, ЧП; Ройтер, Х.; Вулфле, Дж.; и др. (2014). «Синдром Канту, возникающий в результате активации мутации гена KCNJ8» . Человеческая мутация . 35 (7): 809–813. дои : 10.1002/humu.22555 . ПМЦ 4277879 . ПМИД 24700710 .
- ^ Браунштейн, Калифорния; Таун, MC; Люкетт, LJ; и др. (2013). «Мутация KCNJ8 у пациента с синдромом Канту с уникальными сосудистыми аномалиями - подтверждение роли каналов K (АТФ) в этом состоянии» . Европейский журнал медицинской генетики . 56 (12): 678–682. дои : 10.1016/j.ejmg.2013.09.009 . ПМК 3902017 . ПМИД 24176758 .
- ^ Чонг, JX; Макмиллин, MJ; Шивели, К.М.; и др. (2015). «Мутации de novo в NALCN вызывают синдром, характеризующийся врожденными контрактурами конечностей и лица, гипотонией и задержкой развития» . Американский журнал генетики человека . 96 (3): 462–473. дои : 10.1016/j.ajhg.2015.01.003 . ПМЦ 4375444 . ПМИД 25683120 .
- ^ Узун, С; Гекче, С.; Вагнер, К. (2005). «Мутации гена регулятора трансмембранной проводимости при муковисцидозе у бесплодных мужчин с врожденным двусторонним отсутствием семявыносящих протоков» . Журнал экспериментальной медицины Тохоку . 207 (4): 279–285. дои : 10.1620/tjem.207.279 . ПМИД 16272798 .
- ^ Вильшанский, М.; Дюпюи, А.; Эллис, Л.; и др. (2006). «Мутации в гене трансмембранного регулятора муковисцидоза и трансэпителиальные потенциалы in vivo» . Американский журнал респираторной медицины и медицины интенсивной терапии . 174 (7): 787–794. дои : 10.1164/rccm.200509-1377OC . ПМК 2648063 . ПМИД 16840743 .
- ^ Пуарье, К.; Виот, Г.; Ломбарди, Л.; Жони, К.; Биллюарт, П.; Бьенвеню, Т. (2017). «Потеря функции KCNC1 связана с умственной отсталостью без судорог» . Европейский журнал генетики человека . 25 (5): 560–564. дои : 10.1038/ejhg.2017.3 . ПМЦ 5437909 . ПМИД 28145425 .
- ^ Вил, Э.Л; Хасан, М.; Уолш, Ю.; и др. (2014). «Восстановление тока через мутированные калиевые каналы TASK3, лежащие в основе синдрома Бирка Бареля». Молекулярная фармакология . 85 (3): 397–407. дои : 10.1124/моль.113.090530 . ПМИД 24342771 . S2CID 14790826 .
- ^ Барель, О; Шалев, С.А.; Офир Р.; и др. (2008). «Наследственный по материнской линии синдром умственной отсталости Бирка Бареля, вызванный мутацией геномно импринтированного калиевого канала KCNK9» . Американский журнал генетики человека . 83 (2): 193–199. дои : 10.1016/j.ajhg.2008.07.010 . ПМК 2495061 . ПМИД 18678320 .
- ^ Jump up to: а б Глойн, Анна Л; Пирсон, Юэн Р.; Антклифф, Дженнифер Ф.; и др. (2004). «Активация мутаций в гене, кодирующем АТФ-чувствительную субъединицу Kir6.2 калийного канала, и постоянный неонатальный диабет» (PDF) . Медицинский журнал Новой Англии . 350 (18): 1838–1849. doi : 10.1056/NEJMoa032922 . ПМИД 15115830 .
- ^ Ли, член парламента; Равенел, доктор медицинских наук; Ху, Р.Дж.; и др. (2000). «Направленное нарушение гена Kvlqt1 вызывает глухоту и гиперплазию желудка у мышей» . Журнал клинических исследований . 106 (12): 1447–1455. дои : 10.1172/JCI10897 . ПМЦ 387258 . ПМИД 11120752 .
- ^ Jump up to: а б Вексберг, Р; Нисикава, Дж.; Калусериу, О.; и др. (2001). «Развитие опухоли при синдроме Беквита-Видемана связано с различными конституциональными молекулярными изменениями 11p15, включая дефекты импринтинга KCNQ1OT1» . Молекулярная генетика человека . 10 (26): 2989–3000. дои : 10.1093/hmg/10.26.2989 . ПМИД 11751681 .
- ^ Мур, ES; Уорд, Р.Э; Эскобар, ЛФ; Карлин, Мэн (2000). «Гетерогенность синдрома Видемана-Беквита: антропометрические данные». Американский журнал медицинской генетики . 90 (4): 283–290. doi : 10.1002/(SICI)1096-8628(20000214)90:4<283::AID-AJMG4>3.0.CO;2-F . ПМИД 10710224 .
- ^ Вэнь, Х.; Вейгер, ТМ; Фергюсон, Т.С.; и др. (2005). «Канал KCNQ дрозофилы, необходимый для раннего эмбрионального развития» . Журнал неврологии . 25 (44): 10147–10156. doi : 10.1523/JNEUROSCI.3086-05.2005 . ПМК 6725806 . ПМИД 16267222 .
- ^ Ривас, А; Фрэнсис, HW (2005). «Аномалии внутреннего уха у мышей с нокаутом Kcnq1 (Kvlqt1): модель синдрома Джервелла и Ланге-Нильсена». Отология и невротология . 26 (3): 415–424. дои : 10.1097/01.mao.0000169764.00798.84 . ПМИД 15891643 . S2CID 1700736 .
- ^ Казимиро, MC; Ноллманн, Британская Колумбия; Ямоа, Э. Н.; Не, Л; Вари-младший, Джей Си; Сиренко С.Г.; Грин, А.Е.; Гринберг, А; Хуанг, СП; Эберт, С.Н.; Пфайфер, К. (2004). «Направленный точечный мутагенез мышиного Kcnq1: фенотипический анализ мышей с точечными мутациями, вызывающими синдром Романо-Уорда у людей». Геномика . 84 (3): 555–564. дои : 10.1016/j.ygeno.2004.06.007 . ПМИД 15498462 .
- ^ Шуабе, К; Нейруд, Н; Гишени, П; и др. (1997). «Свойства мутаций K+-канала KvLQT1 у Романо-Уорда, Джервелла и Ланге-Нильсена, унаследовавших сердечные аритмии» . Журнал ЭМБО . 16 (17): 5472–5479. дои : 10.1093/emboj/16.17.5472 . ПМК 1170178 . ПМИД 9312006 .
- ^ Бендаху, С; Дональдсон, MR; Пластер, Нью-Йорк; и др. (2003). «Дефектный трафик Kir2.1 калиевого канала лежит в основе синдрома Андерсена-Тавиля» . Журнал биологической химии . 278 (51): 51779–51785. дои : 10.1074/jbc.M310278200 . ПМИД 14522976 .
- ^ Кулиат, Коннектикут; Стаббс, Эл Джей; Войчик Р.П.; Рассел, LB; Джонсон, ДК; Ринчик, Э.М. (1995). «Дефицит субъединицы бета-3 типа рецептора гамма-аминомасляной кислоты вызывает расщелину неба у мышей». Природная генетика . 11 (3): 344–346. дои : 10.1038/ng1195-344 . ПМИД 7581464 . S2CID 19397785 .
- ^ Ви, Э.Л; Циммерман, Э. Ф. (1985). «Поглощение ГАМК в мезенхимальных клетках эмбрионального неба двух линий мышей». Нейрохимические исследования . 10 (12): 1673–1688. дои : 10.1007/bf00988609 . ПМИД 4088436 . S2CID 26049392 .
- ^ Хоманикс, GE; Делори, ТМ; Файерстоун, LL; и др. (1997). «Мыши, лишенные субъединицы бета3-рецептора гамма-аминобутиратного типа, страдают эпилепсией, расщелиной неба и сверхчувствительным поведением» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (8): 4143–4148. Бибкод : 1997PNAS...94.4143H . дои : 10.1073/pnas.94.8.4143 . ПМК 20582 . ПМИД 9108119 .
- ^ Рок, младший; Футтнер, ЧР; Харф, Б.Д. (2008). «Трансмембранный белок TMEM16A необходим для нормального развития трахеи мышей» . Биология развития . 321 (1): 141–149. дои : 10.1016/j.ydbio.2008.06.009 . ПМИД 18585372 .
- ^ Ракич, П; Сидман, Р.Л. (1973). «Последовательность аномалий развития, приводящих к дефициту гранулярных клеток в коре мозжечка мышей-мутантов Weaver». Журнал сравнительной неврологии . 152 (2): 103–132. дои : 10.1002/cne.901520202 . ПМИД 4128371 . S2CID 6553698 .
- ^ Ракич, П; Сидман, Р.Л. (1973). «Мозжечок мыши-мутанта Уивера: дефектная миграция нейронов, вторичная по отношению к аномалии глии Бергмана» . Труды Национальной академии наук Соединенных Штатов Америки . 70 (1): 240–244. Бибкод : 1973ПНАС...70..240Р . дои : 10.1073/pnas.70.1.240 . ПМК 433223 . ПМИД 4509657 .
- ^ Хаттен, МЭ; Лием, Р.К.; Мейсон, Калифорния (1986). «Нейроны гранул мозжечка мыши Weaver не могут мигрировать по астроглиальным отросткам дикого типа in vitro» . Журнал неврологии . 6 (9): 2676–2683. doi : 10.1523/jneurosci.06-09-02676.1986 . ПМК 6568692 . ПМИД 3528411 .
- ^ Патил, Н.; Кокс, Д.Р.; Бхат, Д; Фахам, М; Майерс, Р.М.; Петерсон, А.С. (1995). «Мутация калиевого канала у мышей-ткачей влияет на возбудимость мембраны при дифференцировке гранулярных клеток». Природная генетика . 11 (2): 126–129. дои : 10.1038/ng1095-126 . ПМИД 7550338 . S2CID 23470275 .
- ^ Дэн, GQ; Чжао, X.; Лиз-Миллер, JP; Куинн, Франция; Ли, П.; Ранкур, Делавэр; Лондон, Б.; Кросс, Дж. К.; Дафф, HJ (2008). «Гомозиготная миссенс-мутация калиевого канала N629D hERG (KCNH2) вызывает дефекты развития правого желудочка и его выносящего тракта, а также эмбриональную летальность» . Исследование кровообращения . 103 (12): 1483–1491. дои : 10.1161/CIRCRESAHA.108.177055 . ПМЦ 2774899 . ПМИД 18948620 .
- ^ Jump up to: а б Чем, Б.Л.; Гус, Дж.А.; Сарвер, Алабама; и др. (2014). «Роль KCNQ1 в развитии рака желудочно-кишечного тракта у мышей и человека» . Онкоген . 33 (29): 3861–3868. дои : 10.1038/onc.2013.350 . ПМЦ 3935979 . ПМИД 23975432 .
- ^ Монтейро, Дж; Айрес, Р; Беккер, JD; Хасинто, А; Серталь, AC; Родригес-Леон, Дж (2014). «Активность протонной накачки V-АТФазы необходима для регенерации придатков взрослых рыбок данио» . ПЛОС ОДИН . 9 (3): е92594. Бибкод : 2014PLoSO...992594M . дои : 10.1371/journal.pone.0092594 . ПМЦ 3966808 . ПМИД 24671205 .
- ^ Левин, М; Торлин, Т; Робинсон, КР; Ноги, Т; Меркола, М. (2002). «Асимметрия H+/K+-АТФазы и потенциалов клеточных мембран представляет собой очень раннюю стадию формирования паттерна «лево-право» . Клетка . 111 (1): 77–89. дои : 10.1016/s0092-8674(02)00939-x . ПМИД 12372302 . S2CID 2502945 .
- ^ Дубок, В; Реттингер, Э; Лапраз, Ф; и др. (2005). «Лево-правая асимметрия у эмбриона морского ежа регулируется узловой передачей сигналов на правой стороне» . Развивающая клетка . 9 (1): 147–158. дои : 10.1016/j.devcel.2005.05.008 . ПМИД 15992548 .
- ^ Ивашита, М; Ватанабэ, М; Исии, М; и др. (2006). «Пигментный рисунок у ягуара/обеликса данио вызван мутацией Kir7.1: значение для регуляции движения меланосом» . ПЛОС Генетика . 2 (11): е197. doi : 10.1371/journal.pgen.0020197 . ПМК 1657052 . ПМИД 17121467 .
- ^ Тур, Дж; Чапаламадугу, К.К.; Падавер, Т; Бадоле, С.Л.; Килфойл Пи Джей, 2-й; Бхатнагар, А; Типпараджу, С.М. (2016). «Удаление субъединицы Kvβ1.1 приводит к электрическим и гемодинамическим изменениям, вызывающим гипертрофию сердца в сердце самок мышей» . Экспериментальная физиология . 101 (4): 494–508. дои : 10.1113/EP085405 . ПМЦ 4827621 . ПМИД 27038296 .
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Чопра, С.С.; Страуд, Д.М.; Ватанабэ, Х; Беннетт, Дж. С.; Бернс, К.Г.; Уэллс, К.С.; Ян, Т; Чжун, Т.П; Роден, Д.М. (2010). «Потенциал-управляемые натриевые каналы необходимы для развития сердца у рыбок данио» . Исследование кровообращения . 106 (8): 1342–1350. дои : 10.1161/CIRCRESAHA.109.213132 . ПМЦ 2869449 . ПМИД 20339120 .
- ^ Шу, Х; Ченг, К; Патель, Н; и др. (2003). «Na,K-АТФаза необходима для эмбрионального развития сердца рыбок данио» . Разработка . 130 (25): 6165–6173. дои : 10.1242/dev.00844 . ПМИД 14602677 .
- ^ Харе, С; Ник, Дж. А.; Чжан, Ю; и др. (2017). «Мутация KCNC3 вызывает непрогрессирующий подтип SCA13, связанный с развитием нервной системы, связанный с доминирующими негативными эффектами и аберрантной торговлей EGFR» . ПЛОС ОДИН . 12 (5): e0173565. Бибкод : 2017PLoSO..1273565K . дои : 10.1371/journal.pone.0173565 . ПМЦ 5414954 . ПМИД 28467418 .
- ^ Старич, Т.А.; Холл, ДХ; Гринштейн, Д. (2014). «Два класса каналов щелевых соединений опосредуют взаимодействия сомы и зародышевой линии, необходимые для пролиферации зародышевой линии и гаметогенеза у Caenorhabditis elegans» . Генетика . 198 (3): 1127–1153. дои : 10.1534/genetics.114.168815 . ПМЦ 4224157 . ПМИД 25195067 .
- ^ Бауэр, Р; Леманн, К; Мартини, Дж; Экардт, Ф; Хох, М. (2004). «Белок канала щелевого соединения иннексин 2 необходим для морфогенеза эпителия у эмбриона дрозофилы» . Молекулярная биология клетки . 15 (6): 2992–3004. дои : 10.1091/mbc.E04-01-0056 . ПМК 420120 . ПМИД 15047872 .
- ^ Бауэр, Р; Леманн, К; Фусс, Б; и др. (2002). «Ген канала щелевого соединения дрозофилы иннексин 2 контролирует развитие передней кишки в ответ на передачу сигналов Wingless» . Журнал клеточной науки . 115 (Часть 9): 1859–1867. дои : 10.1242/jcs.115.9.1859 . ПМИД 11956317 .
- ^ Ричард, М.; Хох, М (2015). «Размер глаз дрозофилы определяется иннексин-2-зависимой декапентаплегической передачей сигналов» . Биология развития . 408 (1): 26–40. дои : 10.1016/j.ydbio.2015.10.011 . ПМИД 26455410 .
- ^ Дебир, П; Ван Эш, Х; Гюисманс, К; Пейкелс, Э; Де Смет, Л; Ван Де Вен, Ж; Девриендт, К; Фринс, JP (2005). «Новые мутации GJA1 у пациентов с окуло-денто-цифровой дисплазией (ODDD)». Европейский журнал медицинской генетики . 48 (4): 377–387. дои : 10.1016/j.ejmg.2005.05.003 . ПМИД 16378922 .
- ^ Пиццути, А; Флекс, Э; Мингарелли, Р; Сальпьетро, К; Зеланте, Л; Даллапиккола, Б. (2004). «Гомозиготная мутация гена GJA1 вызывает фенотип спектра Халлермана-Штрайффа/ODDD» . Человеческая мутация . 23 (3): 286. doi : 10.1002/humu.9220 . ПМИД 14974090 . S2CID 13345970 .
- ^ Юарт, Дж. Л.; Коэн, МФ; Мейер, Р.А.; Хуанг, Г.Ю.; Вессельс, А; Гурди, Р.Г.; Чин, Эй Джей; Парк, С.М.; Лазатин Б. О; Виллабон, С; Ло, CW (1997). «Дефекты сердца и нервной трубки у трансгенных мышей со сверхэкспрессией гена щелевого соединения Cx43». Разработка . 124 (7): 1281–1292. дои : 10.1242/dev.124.7.1281 . ПМИД 9118799 . S2CID 189036 .
- ^ Реоме, AG; Де Соуза, Пенсильвания; Кулкарни, С; Ланжиль, Б.Л.; Чжу, Д; Дэвис, ТК; Джунеха, Южная Каролина; Киддер, GM; Россант, Дж (1995). «Пороки развития сердца у новорожденных мышей, лишенных коннексина43». Наука . 267 (5205): 1831–1834. Бибкод : 1995Sci...267.1831R . дои : 10.1126/science.7892609 . ПМИД 7892609 .
- ^ Бритц-Каннингем, С.Х; Шах, ММ; Зуппан, CW; Флетчер, WH (1995). «Мутации гена щелевого соединения Connexin43 у пациентов с пороками развития сердца и дефектами латеральности» . Медицинский журнал Новой Англии . 332 (20): 1323–1329. дои : 10.1056/NEJM199505183322002 . ПМИД 7715640 .
- ^ Чивителли, Р. (2008). «Клеточно-клеточная связь в линии остеобластов/остеоцитов» . Архив биохимии и биофизики . 473 (2): 188–192. дои : 10.1016/j.abb.2008.04.005 . ПМК 2441851 . ПМИД 18424255 .
- ^ Левин, М; Меркола, М. (1999). «Передача сигналов формирования левого и правого паттернов, опосредованная щелевыми соединениями, в ранней бластодерме цыпленка находится выше асимметрии Shh в узле». Разработка . 126 (21): 4703–4714. дои : 10.1242/dev.126.21.4703 . ПМИД 10518488 .
- ^ Беккер, Д.Л.; МакГоннелл, я; Макаренкова, Х.П.; Патель, К; Щекотать, С; Лоример, Дж; Грин, CR (1999). «Роль альфа-1-коннексина в морфогенезе куриных эмбрионов выявлена с использованием нового антисмыслового подхода». Генетика развития . 24 (1–2): 33–42. doi : 10.1002/(SICI)1520-6408(1999)24:1/2<33::AID-DVG5>3.0.CO;2-F . ПМИД 10079509 .
- ^ Леканда, Ф; Варлоу, П.М.; Шейх, С; Фурлан, Ф; Стейнберг, TH; Чивителли, Р. (2000). «Дефицит коннексина 43 вызывает задержку окостенения, черепно-лицевые аномалии и дисфункцию остеобластов» . Журнал клеточной биологии . 151 (4): 931–944. дои : 10.1083/jcb.151.4.931 . ПМК 2169447 . ПМИД 11076975 .
- ^ Арайя, Р; Экардт, Д; Рикельме, Массачусетс; Виллеке, К; Саес, Дж. К. (2003). «Присутствие и важность коннексина 43 во время миогенеза». Клеточная связь и адгезия . 10 (4–6): 451–456. дои : 10.1080/cac.10.4-6.451.456 . hdl : 10533/174413 . ПМИД 14681056 . S2CID 33491307 .
- ^ Канади, JD; Деллинджер, М.Т; Мангер, С.Дж.; Витте, М.Х.; Саймон, AM (2011). «Дефицит коннексина 37 и коннексина 43 у мышей нарушает развитие лимфатических клапанов и приводит к лимфатическим расстройствам, включая лимфедему и хилоторакс» . Биология развития . 354 (2): 253–266. дои : 10.1016/j.ydbio.2011.04.004 . ПМК 3134316 . ПМИД 21515254 .
- ^ Канади, JD; Мангер, С.Дж.; Витте, М.Х.; Саймон, AM (2015). «Сочетание делеций Foxc2 и Connexin37 у мышей приводит к серьезным нарушениям роста и ремоделирования лимфатических сосудов» . Биология развития . 405 (1): 33–46. дои : 10.1016/j.ydbio.2015.06.004 . ПМЦ 4529811 . ПМИД 26079578 .
- ^ Кумаи, М; Нишии, К; Накамура, К; Такеда, Н.; Сузуки, М; Шибата, Ю (2000). «Потеря коннексина 45 вызывает дефект подушки на раннем этапе кардиогенеза». Разработка . 127 (16): 3501–3512. дои : 10.1242/dev.127.16.3501 . ПМИД 10903175 .
- ^ Нишии, К; Кумаи, М; Шибата, Ю (2001). «Регуляция эпителиально-мезенхимальной трансформации через каналы щелевых соединений в развитии сердца». Тенденции сердечно-сосудистой медицины . 11 (6): 213–218. дои : 10.1016/s1050-1738(01)00103-7 . ПМИД 11673050 .
- ^ Уайт, Т.В. (2002). «Уникальный и избыточный вклад коннексина в развитие хрусталика». Наука . 295 (5553): 319–320. Бибкод : 2002Sci...295..319W . дои : 10.1126/science.1067582 . ПМИД 11786642 . S2CID 25744002 .
- ^ Чанг, К; Тан, Вт; Ким, Ю; Лин, X (2015). «Временное условное отсутствие коннексина26 у мышей показывает временную потребность в коннексине26 в ключевых событиях развития улитки до появления слуха». Нейробиология болезней . 73 : 418–427. дои : 10.1016/j.nbd.2014.09.005 . ПМИД 25251605 . S2CID 207068577 .
- ^ Ватанабэ, М; Ивашита, М; Исии, М; Курачи, Ю; Каваками, А; Кондо, С; Окада, Н. (2006). «Пятнистый рисунок леопарда Danio вызван мутацией в гене коннексина 41.8 рыбки данио» . Отчеты ЭМБО . 7 (9): 893–897. дои : 10.1038/sj.embor.7400757 . ПМЦ 1559663 . ПМИД 16845369 .
- ^ Айовин, М.К.; Хиггинс, Э.П.; Хиндес, А; Коблиц, Б; Джонсон, С.Л. (2005). «Мутации в коннексине 43 (GJA1) нарушают рост костей в плавниках рыбок данио» . Биология развития . 278 (1): 208–219. дои : 10.1016/j.ydbio.2004.11.005 . ПМИД 15649473 .
- ^ Дэви, А; Буш, Дж.О.; Сориано, П. (2006). «Подавление связи щелевых соединений на эктопических границах Эф/эфрин лежит в основе краниофронтоназального синдрома» . ПЛОС Биология . 4 (10): е315. дои : 10.1371/journal.pbio.0040315 . ПМЦ 1563491 . ПМИД 16968134 .
- ^ Симс-младший, К.; Эбл, Д.М.; Айовин, МК (2009). «Коннексин43 регулирует расположение суставов в плавниках рыбок данио» . Биология развития . 327 (2): 410–418. дои : 10.1016/j.ydbio.2008.12.027 . ПМЦ 2913275 . ПМИД 19150347 .
- ^ Хоптак-Солга, А.Д.; Нильсен, С; Джайн, я; Таммел, Р; Хайд, Д.Р.; Айовин, МК (2008). «Коннексин43 (GJA1) необходим в популяции делящихся клеток во время регенерации плавников» . Биология развития . 317 (2): 541–548. дои : 10.1016/j.ydbio.2008.02.051 . ПМК 2429987 . ПМИД 18406403 .
- ^ Смендзюк, С.М.; Мессенберг, А; Фогль, А.В.; Таненцапф, Г (2015). «Двунаправленная связь сомы и зародышевой линии, опосредованная щелевыми соединениями, необходима для сперматогенеза» . Разработка . 142 (15): 2598–2609. дои : 10.1242/dev.123448 . ПМК 6514411 . ПМИД 26116660 .
- ^ О, С. К.; Шин, Дж.О.; Бэк, Дж. И.; Ли, Дж; Бэ, JW; Анкамердди, Х; Ким, MJ; Ха, Т.Л.; Рю, З.Ю.; Ким, Великобритания; Бок, Дж; Ли, К.Ю. (2015). «Паннексин 3 необходим для нормального развития скелета у позвоночных» . Журнал ФАСЭБ . 29 (11): 4473–4484. дои : 10.1096/fj.15-273722 . ПМИД 26183770 . S2CID 8219978 .
- ^ Онкал, Р; Джамгоз, МБ (2009). «Молекулярная фармакология экспрессии потенциалзависимых натриевых каналов при метастатическом заболевании: клинический потенциал неонатального Nav1.5 при раке молочной железы». Европейский журнал фармакологии . 625 (1–3): 206–219. дои : 10.1016/j.ejphar.2009.08.040 . ПМИД 19835862 .
- ^ Jump up to: а б Хаус, CD; Васке, CJ; Шварц, AM; Обиас, В; Фрэнк, Б; Луу, Т; Сарвазян Н; Ирби, Р; Штраусберг, Р.Л.; Хейлз, Т.Г.; Стюарт, Дж. М.; Ли, Нью-Хэмпшир (2010). «Потенциал-управляемый Na+-канал SCN5A является ключевым регулятором сети транскрипции генов, которая контролирует инвазию рака толстой кишки» . Исследования рака . 70 (17): 6957–6967. дои : 10.1158/0008-5472.CAN-10-1169 . ПМЦ 2936697 . ПМИД 20651255 .
- ^ Перес-Нейт, М; Рао, В.Р.; Джентиле, С. (2016). «Активация HERG1/Kv11.1 стимулирует транскрипцию p21waf/cip в клетках рака молочной железы посредством кальциневрин-зависимого механизма» . Онкотаргет . 7 (37): 58893–58902. дои : 10.18632/oncotarget.3797 . ПМК 5312283 . ПМИД 25945833 .
- ^ Лансу, К; Джентиле, С. (2013). «Активация калиевых каналов подавляет пролиферацию клеток рака молочной железы, активируя программу старения» . Смерть клеток и болезни . 4 (6): е652. дои : 10.1038/cddis.2013.174 . ПМЦ 3698542 . ПМИД 23744352 .
- ^ Пей, Л; Мудрец, О; Славин А; Грязь; Пауэрс, С; Ян, Л.Ю.; Хоуи, Т. (2003). «Онкогенный потенциал TASK3 (Kcnk9) зависит от функции K+-канала» . Труды Национальной академии наук . 100 (13): 7803–7807. Бибкод : 2003PNAS..100.7803P . дои : 10.1073/pnas.1232448100 . ПМК 164668 . ПМИД 12782791 .
- ^ Сайто, Цуёси; Шлегель, Ричард; Андрессон, Тиркелл; Юге, Луи; Ямамото, Масао; Ямасаки, Хироши (1998). «Индукция клеточной трансформации мутированной вакуолярной H+-аттазой 16К (дуктином) сопровождается подавлением межклеточной коммуникации щелевых соединений и транслокацией коннексина 43 в клетках NIH3T3» . Онкоген . 17 (13): 1673–1680. дои : 10.1038/sj.onc.1202092 . ПМИД 9796696 .
- ^ Гупта, Н; Мартин, П.М.; Прасад, П.Д.; Ганапати, В. (2006). «SLC5A8 (SMCT1)-опосредованный транспорт бутирата лежит в основе опухолесупрессирующей функции транспортера». Науки о жизни . 78 (21): 2419–2425. дои : 10.1016/j.lfs.2005.10.028 . ПМИД 16375929 .
- ^ Репке, Т.К.; Пертелл, К; Кинг, EC; Ла Перль, К.М.; Лернер, диджей; Эбботт, GW (2010). «Направленное удаление Kcne2 вызывает глубокий кистозный гастрит и неоплазию желудка» . ПЛОС ОДИН . 5 (7): е11451. Бибкод : 2010PLoSO...511451R . дои : 10.1371/journal.pone.0011451 . ПМЦ 2897890 . ПМИД 20625512 .
- ^ Ли, член парламента; Ху, Р.Дж.; Джонсон, Лос-Анджелес; и др. (1997). «Человеческий ген KVLQT1 демонстрирует тканеспецифический импринтинг и включает хромосомные перестройки синдрома Беквита-Видемана». Природная генетика . 15 (2): 181–185. дои : 10.1038/ng0297-181 . ПМИД 9020845 . S2CID 24715509 .
- ^ Мартино, Джей Джей; Уолл, бакалавр искусств; Мастрантони, Э; Вилимчик, Б.Дж.; Ла Кава, С.Н.; Дегенхардт, К; Уайт, Э; Чен, С (2013). «Метаботропный глутаматный рецептор 1 (Grm1) является онкогеном в эпителиальных клетках» . Онкоген . 32 (37): 4366–4376. дои : 10.1038/onc.2012.471 . ПМЦ 3910169 . ПМИД 23085756 .
- ^ Шпейер, CL; Смит, Дж. С.; Банда, М; и др. (2012). «Метаботропный глутаматный рецептор-1: потенциальная терапевтическая мишень для лечения рака молочной железы» . Исследование и лечение рака молочной железы . 132 (2): 565–573. дои : 10.1007/s10549-011-1624-x . ПМЦ 3898178 . ПМИД 21681448 .
- ^ Чжан, Дж. Т; Цзян, X. H; Се, С; Ченг, Х; Да Донг, Дж; Ван, Ю; Фок, К.Л.; Чжан, XH; Солнце, Т.Т; Цанг, Л.Л; Чен, Х; Сан, XJ; Чанг, Ю.В.; Цай, З.М; Цзян, WG; Чан, ХК (2013). «Понижение уровня CFTR способствует эпителиально-мезенхимальному переходу и связано с плохим прогнозом рака молочной железы» . Biochimica et Biophysical Acta (BBA) - Исследования молекулярных клеток . 1833 (12): 2961–2969. дои : 10.1016/j.bbamcr.2013.07.021 . ПМИД 23916755 .
- ^ Се, С; Цзян, X. H; Чжан, Дж. Т; и др. (2013). «CFTR подавляет прогрессирование опухоли посредством миР-193b, воздействующих на активатор урокиназы плазминогена (uPA) при раке простаты». Онкоген . 32 (18): 2282–2291, 2291.e1–7. дои : 10.1038/onc.2012.251 . ПМИД 22797075 . S2CID 21255355 .
- ^ Сирнес, С.; Брюун, Дж.; Кольберг, М.; и др. (2012). «Коннексин43 действует как супрессор опухоли колоректального рака и предсказывает исход заболевания». Международный журнал рака . 131 (3): 570–581. дои : 10.1002/ijc.26392 . ПМИД 21866551 . S2CID 6293505 .
- ^ Шиклинг, Б.М.; Англия, Южная Каролина; Айкин-Бернс, Н.; и др. (2015). «Ингибитор каналов BKCa модулирует туморогенную способность гормон-независимых клеток рака молочной железы через путь Wnt» . Отчеты онкологии . 33 (2): 533–538. дои : 10.3892/или.2014.3617 . ПМК 4306270 . ПМИД 25422049 .
- ^ Фельдер, CC; Макартур, Л; Ма, А.Л; и др. (1993). «Опухолесупрессорная функция мускариновых рецепторов ацетилхолина связана с активацией рецептор-управляемого притока кальция» . Труды Национальной академии наук Соединенных Штатов Америки . 90 (5): 1706–1710. Бибкод : 1993PNAS...90.1706F . дои : 10.1073/pnas.90.5.1706 . ПМК 45948 . ПМИД 7680475 .
- ^ Резания, С; Каммерер, С; Ли, С; Штайнекер-Фронвизер, Б; Горишек, А; Девани, Т.Т; Верхейен, С; Пассеггер, Калифорния; Тебризи-Визси, Н.Г.; Хакл, Х; Платцер, Д; Зарнани, А.Х; Малле, Э; Ян, SW; Бауэрнхофер, Т; Шрайбмайер, В. (2016). «Сверхэкспрессия вариантов сплайсинга гена KCNJ3 оказывает противоположное влияние на жизненно важные параметры линии клеток злокачественного рака молочной железы MCF-7» . БМК Рак . 16 : 628. doi : 10.1186/s12885-016-2664-8 . ПМЦ 4983040 . ПМИД 27519272 .
- ^ Каммерер, С; Соколовский, А; Хакл, Х; и др. (2016). «KCNJ3 является новым независимым прогностическим маркером для пациентов с раком молочной железы с положительным рецептором эстрогена» . Онкотаргет . 7 (51): 84705–84717. дои : 10.18632/oncotarget.13224 . ПМЦ 5356693 . ПМИД 27835900 .
Внешние ссылки
[ редактировать ] Цитаты, связанные с развитием биоэлектричества в Wikiquote
Цитаты, связанные с развитием биоэлектричества в Wikiquote