силурийский
| силурийский | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
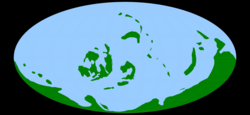 Карта Земли в раннем силуре, ок. 440 млн лет назад [ нужна ссылка ] | |||||||||||||
| Хронология | |||||||||||||
| |||||||||||||
| Этимология | |||||||||||||
| Формальность имени | Формальный | ||||||||||||
| Синоним(ы) | Готландский | ||||||||||||
| Информация об использовании | |||||||||||||
| Небесное тело | Земля | ||||||||||||
| Региональное использование | Глобальный ( ICS ) | ||||||||||||
| Используемая шкала времени | Временная шкала ICS | ||||||||||||
| Определение | |||||||||||||
| Хронологическая единица | Период | ||||||||||||
| Стратиграфическая единица | Система | ||||||||||||
| Впервые предложено | Родерик Мерчисон , 1835 г. | ||||||||||||
| Формальность временного интервала | Формальный | ||||||||||||
| Определение нижней границы | FAD граптолита ascensus Akidograptus | ||||||||||||
| Нижняя граница ГССП | Добс Линн , Моффат, Великобритания 55 ° 26'24 "N 3 ° 16'12" W / 55,4400 ° N 3,2700 ° W | ||||||||||||
| Нижний GSSP ратифицирован | 1984 [ 4 ] [ 5 ] | ||||||||||||
| Определение верхней границы | FAD граптолита Monograptus униформис | ||||||||||||
| Верхняя граница ГССП | Клонк , Чехия 49 ° 51'18 "N 13 ° 47'31" E / 49,8550 ° N 13,7920 ° E | ||||||||||||
| Верхний GSSP ратифицирован | 1972 [ 6 ] | ||||||||||||
| Атмосферные и климатические данные | |||||||||||||
| Уровень моря выше современного | Около 180 м, с кратковременными отрицательными экскурсиями. [ 7 ] | ||||||||||||

Силурийский i ( / s ɪ ˈ lj ʊər sih- . ən , s aɪ / ) LURE -ee-ən, sy- - [ 8 ] [ 9 ] [ 10 ] — это геологический период и система, охватывающая 24,6 миллиона лет от конца ордовикского периода, 443,8 миллиона лет назад ( млн лет назад ), до начала девонского периода, 419,2 млн лет назад. [ 11 ] Силурий — самый короткий период палеозойской эры. Как и в случае с другими геологическими периодами , пласты горных пород , определяющие начало и конец периода, хорошо идентифицированы, но точные даты неопределенны на несколько миллионов лет. Основа силурийского периода приходится на серию крупных событий ордовикско-силурийского вымирания, когда было уничтожено до 60% морских родов.
Одним из важных событий этого периода было первоначальное зарождение земной жизни в ходе так называемой силурийско-девонской земной революции : сосудистые растения возникли из более примитивных наземных растений, [ 12 ] [ 13 ] дикарийские грибы начали расширяться и диверсифицироваться вместе с гломеромикотановыми грибами, [ 14 ] а три группы членистоногих ( многоножки , паукообразные и шестиногие ) стали полностью наземными. [ 15 ]
Другой важной вехой эволюции в силурийском периоде стала диверсификация челюстных рыб , к которым относятся плакодермы , акантодии (которые дали начало хрящевым рыбам ) и остихтианы ( костистые рыбы , далее разделенные на лопастеперые и лучепёрые рыбы ), [ 16 ] хотя это соответствовало резкому сокращению численности бесчелюстных рыб , таких как конодонты и остракодермы .
История обучения
[ редактировать ]−4500 — – — – −4000 — – — – −3500 — – — – −3000 — – — – −2500 — – — – −2000 — – — – −1500 — – — – −1000 — – — – −500 — – — – 0 — |
| |||||||||||||||||||||||||||||||||||||||||||||
Силурийская система была впервые идентифицирована шотландским геологом Родериком Мерчисоном , который исследовал содержащие ископаемые осадочные породы на юге Уэльса в начале 1830-х годов. Он назвал последовательности кельтского племени Уэльса силуресами , вдохновленный своим другом Адамом Седжвиком , который назвал период своего исследования Кембрийским периодом , от латинского названия Уэльса. [ 17 ] Хотя британские породы, которые теперь идентифицируются как принадлежащие силурийской системе, и земли, которые, как теперь полагают, были заселены в древности силурами, обнаруживают слабую корреляцию ( ср . Геологическую карту Уэльса , Карту доримских племен Уэльса ), Мерчисон предположил, что их территория включала обнажения Каэр-Карадок и Венлок-Эдж , а в противном случае в других местах существовало множество силурийских скал, «чтобы оправдать предложенное название». [ 18 ] В 1835 году эти двое представили совместный доклад под названием « О силурийских и кембрийских системах, демонстрирующий порядок, в котором древние осадочные пласты сменяют друг друга в Англии и Уэльсе», который стал зародышем современной геологической шкалы времени . [ 19 ] Как было впервые выявлено, «силурийский» ряд, если его проследить дальше, быстро стал перекрывать «кембрийский» эпизод Седжвика, однако это спровоцировало яростные разногласия, которые положили конец дружбе.
Английский геолог Чарльз Лэпворт разрешил конфликт, определив новую ордовикскую систему, включая оспариваемые пласты. [ 20 ] Альтернативное название силура было «Готланд» по названию слоев балтийского острова Готланд . [ 21 ]
Французский геолог Иоахим Барранд , опираясь на работы Мерчисона, использовал термин силурийский период в более широком смысле, чем это было оправдано последующими знаниями. Он разделил силурийские породы Богемии на восемь этапов. [ 22 ] Его интерпретация была подвергнута сомнению в 1854 году Эдвардом Форбсом . [ 23 ] и более поздние стадии Барранда; С тех пор было показано, что F, G и H относятся к девонскому периоду. Несмотря на эти изменения в первоначальных группировках слоев, признано, что Барранд сделал Богемию классической территорией для изучения самых ранних силурийских окаменелостей.
Подразделения
[ редактировать ]| Эпоха | Возраст | Начинать (мья) |
Этимология Эпохи и этапы |
Примечания |
|---|---|---|---|---|
| Лландовери | Радданский | 443.8 | Ферма Сефн-Руддан, Лландовери в Кармартеншире , Уэльс | |
| Аэрониан | 440.8 | Ферма Cwm Coed-Aeron, Уэльс | Трек Трефор рядом с фермой - это место расположения GSSP. | |
| Теличский | 438.5 | Ферма Пен-лан-Телич, Лландовери, Уэльс | ||
| Венлок | Шейнвудский | 433.4 | Деревня Шейнвуд, Мач-Венлок и Венлок-Эдж , Шропшир , Англия | Во время Венлока появляются древнейшие из известных трахеофитов рода Cooksonia . Сложность немного более поздних Гондваны растений , таких как Baragwanathia , которые напоминали современный плаун, указывает на гораздо более длительную историю сосудистых растений, простирающуюся до раннего силура или даже ордовика . [ нужна ссылка ] В Венлоке появляются и первые наземные животные, представленные воздуходышащими многоножками из Шотландии . [ 24 ] |
| гомеровский | 430.5 | Гомер, Шропшир , Англия | ||
| Ладлоу | Горстиан | 427.4 | Gorsty village near Ludlow , Shropshire, England | |
| Лудфордиан | 425.6 | Ладфорд, Шропшир , Англия | ||
| Дополнения | — | 423.0 | Назван в честь местности в заповеднике Гомолка-а-Пржидоли недалеко от Праги пригорода Сливенец , Чехия . | Пржидоли – старое название кадастрового участка поля. [ 25 ] |
Палеогеография
[ редактировать ]
Поскольку суперконтинент Гондвана покрывает экватор и большую часть южного полушария, большой океан занимал большую часть северной половины земного шара. [ 26 ] Высокий уровень моря в силурийском периоде и относительно равнинная местность (с небольшим количеством значительных горных поясов) привели к образованию множества цепочек островов и, следовательно, к богатому разнообразию окружающей среды. [ 26 ]
В силурийском периоде Гондвана продолжала медленный дрейф на юг к высоким южным широтам, но есть свидетельства того, что силурийские ледяные шапки были менее обширными, чем ледяные шапки позднего ордовика. В этот период южные континенты оставались едиными. Таяние ледниковых шапок и ледников способствовало повышению уровня моря, о чем свидетельствует тот факт, что силурийские отложения перекрывают эродированные ордовикские отложения, образуя несогласие . Континенты Авалония , Балтика и Лаврентия слились вместе около экватора , положив начало формированию второго суперконтинента , известного как Еврамерика .
Когда протоЕвропа столкнулась с Северной Америкой, в результате столкновения прибрежные отложения, накапливавшиеся с кембрия, сложились у восточного побережья Северной Америки и западного побережья Европы. Это событие — Каледонская складчатость , волна горообразования, которая простиралась от штата Нью-Йорк через объединенную Европу и Гренландию до Норвегии. В конце силура уровень моря снова упал, оставив характерные бассейны эвапоритов, простирающиеся от Мичигана до Западной Вирджинии, а новые горные хребты быстро подверглись эрозии. Река Тейс , впадающая в мелководное среднеконтинентальное море, размыла пласты ордовикского периода, образовав отложения силурийских пластов в северном Огайо и Индиане.
Огромный океан Панталасса покрывал большую часть северного полушария. Другие второстепенные океаны включают две фазы Тетиса, Прото-Тетис и Палео-Тетис , Рейский океан , Океан Япета (узкий морской путь между Авалонией и Лаврентией) и недавно образовавшийся Уральский океан .

Климат и уровень моря
[ редактировать ]Когда-то считалось, что силурийский период отличался относительно стабильными и теплыми температурами, в отличие от экстремальных оледенений ордовика до него и сильной жары последующего девона; однако теперь известно, что глобальный климат претерпел множество резких колебаний на протяжении силурийского периода. [ 27 ] [ 28 ] о чем свидетельствуют многочисленные крупные отклонения изотопов углерода и кислорода в этот геологический период. [ 29 ] [ 30 ] [ 31 ] Уровень моря поднялся по сравнению с ирнантическим минимумом на протяжении первой половины силура; впоследствии они падали на протяжении остальной части периода, хотя на эту общую тенденцию накладываются модели меньшего масштаба; можно выделить пятнадцать высоких стоянок (периоды, когда уровень моря был выше края континентального шельфа), а самый высокий силурийский уровень моря, вероятно, был примерно на 140 метров (459 футов) выше, чем самый низкий достигнутый уровень. [ 26 ]
В этот период Земля вступила в теплую парниковую фазу, чему способствовали высокие уровни CO 2 (4500 частей на миллион), а теплые мелководные моря покрыли большую часть экваториальной суши. [ 32 ] В начале силура ледники отступили обратно к Южному полюсу , пока почти не исчезли в середине силура. [ 28 ] Слои разбитых раковин (так называемых ракушечников ) служат убедительным доказательством того, что в климате преобладали сильные штормы, порождаемые тогда, как и сейчас, теплыми морскими поверхностями. [ 33 ]
Возмущения
[ редактировать ]Климат и углеродный цикл кажутся довольно нестабильными в силурийский период, когда частота изотопных отклонений (указывающих на колебания климата) была выше, чем в любой другой период. [ 26 ] Событие Иревикен , событие Мулде и событие Лау представляют собой изотопные экскурсии после незначительного массового вымирания. [ 34 ] и связано с быстрым изменением уровня моря. Каждый из них оставляет в геологической летописи схожий след, как геохимический, так и биологический; Особенно сильно пострадали пелагические (свободно плавающие) организмы, а также брахиоподы , кораллы и трилобиты , и вымирание редко происходит в виде быстрой серии быстрых всплесков. [ 26 ] [ 31 ] Колебания климата лучше всего объясняются последовательностью оледенений, но отсутствие тиллитов в среднем и позднем силуре делает это объяснение проблематичным. [ 35 ]
Флора и фауна
[ редактировать ]Некоторые палеонтологи рассматривают силурийский период как продолжительный период восстановления после массового вымирания в позднем ордовике (LOME), которое прервало каскадный рост биоразнообразия, непрерывно продолжавшийся на протяжении кембрия и большей части ордовика. [ 36 ]
Силурий был первым периодом, когда были обнаружены мегаокаменелости обширной наземной биоты в виде моховых миниатюрных лесов вдоль озер и ручьев и сетей крупных микоризных нематофитов , что ознаменовало начало силурийско-девонской земной революции. [ 12 ] [ 13 ] [ 37 ] Однако наземная фауна не оказала серьезного влияния на Землю, пока не диверсифицировалась в девоне. [ 26 ]

Первые ископаемые находки сосудистых растений , то есть наземных растений с тканями, переносящими воду и пищу, появились во второй половине силурийского периода. [ 38 ] Самыми ранними известными представителями этой группы являются Cooksonia . Большинство отложений, содержащих Cooksonia, имеют морскую природу. Предпочтительные места обитания, вероятно, располагались вдоль рек и ручьев. Барагванатия , по-видимому, почти такого же возраста: она датируется ранним Ладлоу (420 миллионов лет). [ нужно обновить? ] имеет ветвящиеся стебли и игольчатые листья длиной 10–20 см (3,9–7,9 дюйма). Растение демонстрирует высокую степень развития по отношению к возрасту его ископаемых остатков. Окаменелости этого растения были обнаружены в Австралии. [ 39 ] [ 40 ] Канада, [ 41 ] и Китай. [ 42 ] Eohostimella heathana — раннее, вероятно, наземное «растение», известное по сжатым окаменелостям. [ 43 ] раннесилурийского (лландоверского) возраста. [ 44 ] Химический состав его окаменелостей похож на химический состав окаменелых сосудистых растений, а не водорослей. [ 43 ]
Из силура известны также окаменелости, считающиеся наземными животными. Самая старая из находок многоножек когда-либо известных - это Kampecaris obanensis и Archidesmus sp. из позднего силура (425 миллионов лет назад) Керреры . [ 45 ] Есть также другие многоножки, многоножки и паукообразные тригонотарбиды , известные из Ладлоу (420 миллионов лет назад). [ 45 ] [ 46 ] [ 47 ] Хищные беспозвоночные указывают на то, что простые пищевые сети существуют , включающие нехищных животных-жертв. Экстраполируя данные биоты раннего девона , Эндрю Джерам и др. в 1990 году [ 48 ] предложил пищевую сеть, основанную на еще не обнаруженных детритофагах и травоядных микроорганизмах. [ 49 ] Многоножки из формации Коуи, такие как Cowiedesmus и Pneumodesmus, считались старейшими многоножками среднего силура, жившими 428–430 миллионов лет назад. [ 24 ] [ 50 ] [ 51 ] интерпретируют возраст этого образования как ранний девон . хотя позже некоторые исследователи [ 52 ] [ 53 ] Тем не менее, Pneumodesmus по- прежнему остается важной окаменелостью как старейшее окончательное свидетельство существования дыхалец, способных вдыхать воздух. [ 45 ]

Появились первые костные рыбы Osteichthyes , представленные акантодиями, покрытыми костной чешуей. Рыбы достигли значительного разнообразия и развили подвижные челюсти , приспособленные к опорам передних двух или трех жаберных дуг. Разнообразная фауна эвриптерид (морских скорпионов) — некоторые из них длиной несколько метров — бродила по мелководным силурийским морям и озерам Северной Америки; многие из их окаменелостей были найдены в штате Нью-Йорк . Брахиоподы были многочисленны и разнообразны, а таксономический состав, экология и биоразнообразие силурийских брахиопод отражали ордовикские. [ 54 ] Брахиоподы, пережившие LOME, разработали новые приспособления к стрессу окружающей среды. [ 55 ] и после массового вымирания они, как правило, были эндемиками одной палеопластики, но впоследствии расширили свой ареал. [ 56 ] Наиболее многочисленными брахиоподами были атрипиды и пентамериды; [ 57 ] атрипиды были первыми, кто восстановился и редиверсифицировался в Руддании после LOME, [ 58 ] в то время как восстановление пентамеридов было отложено до Аэрониана. [ 57 ] Мшанки демонстрировали значительную степень эндемизма к конкретному шельфу. [ 59 ] У них также сложились симбиотические отношения с книдариями. [ 60 ] и строматолиты. [ 61 ] Многие окаменелости двустворчатых моллюсков были также найдены в силурийских отложениях. [ 62 ] с этого периода известны первые глубокосверлящие двустворчатые моллюски. [ 63 ] Пик разнообразия хитонов пришелся на середину силура. [ 64 ] Гедереллоиды пользовались значительным успехом в силурийском периоде, при этом некоторые развивались в симбиозе с колониальным морщинистым кораллом Entelophyllum . [ 65 ] Силурийский период был периодом расцвета тентакулитоидов , [ 66 ] который испытал эволюционное излучение, сосредоточенное главным образом в Балтоскандии, [ 67 ] наряду с расширением их географического ареала в Лландовери и Венлоке. [ 68 ] Трилобиты начали восстанавливаться в Руддане, [ 69 ] и они продолжали добиваться успеха в силурии, как и в ордовике, несмотря на сокращение кладового разнообразия в результате LOME. [ 70 ] Ранний силурийский период был периодом хаотического развития криноидей , поскольку они диверсифицировались после LOME. [ 71 ] Члены Flexibilia, на которых LOME оказал минимальное воздействие, приобретали все большее экологическое значение в силурийских морях. [ 72 ] Монобатридные камерыты, как и гибкие, диверсифицировались в лландовери, тогда как циатокриниды и дендрокриниды диверсифицировались позже в силурийском периоде. [ 73 ] Сцифокриноидные лоболиты внезапно появились в конце силура, незадолго до границы силура и девона, и исчезли так же внезапно, как и появились вскоре после своего первого появления. [ 74 ] Эндобиотические симбионты были распространены у кораллов и строматопороидов. [ 75 ] [ 76 ] В частности, ругозные кораллы были заселены и покрыты разнообразными эпибионтами. [ 77 ] включая некоторые гедереллоиды, как указано выше. [ 65 ] Фотосимбиотические склерактинии впервые появились в среднем силуре. [ 78 ] Обилие рифов было неоднородным; иногда окаменелости встречаются часто, но в других случаях они практически отсутствуют в летописи горных пород. [ 26 ]
-
 Куксония , самое раннее сосудистое растение, средний силур.
Куксония , самое раннее сосудистое растение, средний силур. -
 Окаменелости силурийского морского дна, собранные в заповеднике «Гнездо Рена» , Дадли, Великобритания.
Окаменелости силурийского морского дна, собранные в заповеднике «Гнездо Рена» , Дадли, Великобритания. -
 Фрагменты криноидей в силурийском (Придоли) известняке ( Сааремаа , Эстония)
Фрагменты криноидей в силурийском (Придоли) известняке ( Сааремаа , Эстония) -
 Окаменелости силурийского морского дна, собранные в заповеднике «Гнездо Рена», Дадли, Великобритания.
Окаменелости силурийского морского дна, собранные в заповеднике «Гнездо Рена», Дадли, Великобритания. -
 Eurypterus — распространённый эвриптерид верхнего силура.
Eurypterus — распространённый эвриптерид верхнего силура. -
 Pterygotus был гигантским эвриптеридом, имевшим почти космополитическое распространение (здесь показана реконструкция девонского вида P. anglicus ).
Pterygotus был гигантским эвриптеридом, имевшим почти космополитическое распространение (здесь показана реконструкция девонского вида P. anglicus ). -
 Трилобиты еще были разнообразны и распространены в силурии. Окаменелости Calymene celbra чрезвычайно многочисленны в некоторых частях центральной части США.
Трилобиты еще были разнообразны и распространены в силурии. Окаменелости Calymene celbra чрезвычайно многочисленны в некоторых частях центральной части США. -
 Halysites — таблитчатый коралл , вымершая группа кораллов, жившая в палеозое.
Halysites — таблитчатый коралл , вымершая группа кораллов, жившая в палеозое. -
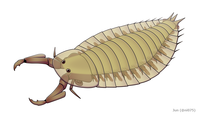 Париоскорпион был загадочным членистоногим из силура Висконсина.
Париоскорпион был загадочным членистоногим из силура Висконсина. -
 Образец Dalmanites limulurus из силурийских отложений Нью-Йорка.
Образец Dalmanites limulurus из силурийских отложений Нью-Йорка. -
 Камень, содержащий несколько геодезированных пентамерид брахиопод из слоев Индианы.
Камень, содержащий несколько геодезированных пентамерид брахиопод из слоев Индианы. -

-
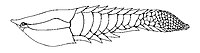
 Джамойтиус был загадочным позвоночным, возможно, родственным анаспиду . рыбе-
Джамойтиус был загадочным позвоночным, возможно, родственным анаспиду . рыбе- -
-
 Tujiaaspis — бесчелюстной галеаспис из раннего силура ( телиха ) Китая, демонстрирующий происхождение парных плавников.
Tujiaaspis — бесчелюстной галеаспис из раннего силура ( телиха ) Китая, демонстрирующий происхождение парных плавников. -

-
 Fanjingshania , климатоидная колючая акула из нижнего силура (аэрона), описанная по разрозненным элементам дермоскелета.
Fanjingshania , климатоидная колючая акула из нижнего силура (аэрона), описанная по разрозненным элементам дермоскелета. -
![Shenacanthus vermiformis[69] — род челюстно-стволовых хондрихтиев из раннего силура (телиха) Китая.](//upload.wikimedia.org/wikipedia/commons/thumb/6/6c/Shenacanthus_vermiformis.jpg/154px-Shenacanthus_vermiformis.jpg) Шенакантус [ 80 ] — род челюстно-стволовых хондрихтиев из раннего силура (телиха) Китая.
Шенакантус [ 80 ] — род челюстно-стволовых хондрихтиев из раннего силура (телиха) Китая. -
![Xiushanosteus[80] — старейшая известная плакодерма раннего силура (телиха) Китая.](//upload.wikimedia.org/wikipedia/commons/thumb/c/c5/Xiushanosteus.jpg/200px-Xiushanosteus.jpg) Ксиушаностей [ 80 ] является старейшей известной плакодермой раннего силура (телиха) Китая.
Ксиушаностей [ 80 ] является старейшей известной плакодермой раннего силура (телиха) Китая. -
 Entelognathus primordialis — рыба -плакодерма позднего силура.
Entelognathus primordialis — рыба -плакодерма позднего силура.
















![Shenacanthus vermiformis[69] — род челюстно-стволовых хондрихтиев из раннего силура (телиха) Китая.](http://upload.wikimedia.org/wikipedia/commons/thumb/6/6c/Shenacanthus_vermiformis.jpg/154px-Shenacanthus_vermiformis.jpg)
![Xiushanosteus[80] — старейшая известная плакодерма раннего силура (телиха) Китая.](http://upload.wikimedia.org/wikipedia/commons/thumb/c/c5/Xiushanosteus.jpg/200px-Xiushanosteus.jpg)

См. также
[ редактировать ]Примечания
[ редактировать ]- ^ Джеппссон, Л.; Калнер, М. (2007). «Силурийское событие Мулде и сценарий для секундо-секундных событий». Труды Королевского общества Эдинбурга по наукам о Земле и окружающей среде . 93 (02): 135–154. дои : 10.1017/S0263593300000377 .
- ^ Муннеке, А.; Самтлебен, К.; Бикерт, Т. (2003). «Событие Иревикен в нижнем силуре Готланда, Швеция, - связь с аналогичными палеозойскими и протерозойскими событиями». Палеогеография, Палеоклиматология, Палеоэкология . 195 (1): 99–124. дои : 10.1016/S0031-0182(03)00304-3 .
- ^ «Диаграмма/Шкала времени» . www.stratigraphy.org . Международная комиссия по стратиграфии.
- ^ Лукас, сентсер (6 ноября 2018 г.). «Метод хроностратиграфии GSSP: критический обзор» . Границы в науках о Земле . 6 : 191. Бибкод : 2018FrEaS...6..191L . дои : 10.3389/feart.2018.00191 .
- ^ Холланд, К. (июнь 1985 г.). «Серии и этапы силурийской системы» (PDF) . Эпизоды . 8 (2): 101–103. дои : 10.18814/epiiugs/1985/v8i2/005 . Проверено 11 декабря 2020 г.
- ^ Хлупач, Иво; Хладил, Индрих (январь 2000 г.). «Глобальный стратотипический разрез и точка границы силура и девона» . CFS Courier Forschungsinstitut Senckenberg . Проверено 7 декабря 2020 г.
- ^ Хак, Бу; Шуттер, СР (2008). «Хронология палеозойских изменений уровня моря». Наука . 322 (5898): 64–68. Бибкод : 2008Sci...322...64H . дои : 10.1126/science.1161648 . ПМИД 18832639 . S2CID 206514545 .
- ^ Уэллс, Джон (3 апреля 2008 г.). Словарь произношения Лонгмана (3-е изд.). Пирсон Лонгман . ISBN 978-1-4058-8118-0 .
- ^ «Силурийский» . Dictionary.com Полный (онлайн). nd
- ^ «Силурийский» . CollinsDictionary.com . ХарперКоллинз .
- ^ «Международная хроностратиграфическая карта, версия 2015/01» (PDF) . Международная комиссия по стратиграфии . Январь 2015.
- ^ Перейти обратно: а б Кэпел, Эллиот; Клил, Кристофер Дж.; Сюэ, Цзиньчжуан; Монне, Клод; Серве, Томас; Каскалес-Миньяна, Борха (август 2022 г.). «Силурийско-девонская наземная революция: модели разнообразия и систематическая ошибка выборки записей макроископаемых сосудистых растений» . Обзоры наук о Земле . 231 : 104085. Бибкод : 2022ESRv..23104085C . doi : 10.1016/j.earscirev.2022.104085 . hdl : 20.500.12210/76731 . S2CID 249616013 .
- ^ Перейти обратно: а б Сюэ, Цзиньчжуан; Хуан, Пу; Ван, Деминг; Сюн, Конхуэй; Лю, Ле; Бейсингер, Джеймс Ф. (май 2018 г.). «Силурийско-девонская наземная революция в Южном Китае: таксономия, разнообразие и эволюция характера сосудистых растений в палеогеографически изолированном низкоширотном регионе» . Обзоры наук о Земле . 180 : 92–125. Бибкод : 2018ESRv..180...92X . doi : 10.1016/j.earscirev.2018.03.004 . Проверено 8 декабря 2022 г.
- ^ Лутзони, Фрэнсис; Новак, Майкл Д.; Альфаро, Майкл Э.; Риб, Валери; Мядликовска, Иоланта; Круг, Майкл; Арнольд, А. Элизабет; Льюис, Луиза А.; Суоффорд, Дэвид Л.; Хиббетт, Дэвид; Хилу, Хидир; Джеймс, Тимоти Ю.; Квандт, Дитмар; Магаллон, Сьюзен (21 декабря 2018 г.). «Современные излучения грибов и растений, связанные с симбиозом» . Природные коммуникации . 9 (1): 5451. Бибкод : 2018NatCo...9.5451L . дои : 10.1038/ s41467-018-07849-9 ПМК 6303338 . ПМИД 30575731 . S2CID 56645104 .
- ^ Гарвуд, Рассел Дж.; Эджкомб, Грегори Д. (сентябрь 2011 г.). «Ранние наземные животные, эволюция и неопределенность» . Эволюция: образование и информационно-пропагандистская деятельность . 4 (3): 489–501. дои : 10.1007/s12052-011-0357-y .
- ^ Бразо, доктор медицины; Фридман, М. (2015). «Происхождение и ранняя филогенетическая история челюстных позвоночных» . Природа . 520 (7548): 490–497. Бибкод : 2015Natur.520..490B . дои : 10.1038/nature14438 . ПМЦ 4648279 . ПМИД 25903631 .
- ^ См.:
- Мерчисон, Родерик Импи (1835). «О силурийской системе пород» . Философский журнал . 3-я серия. 7 (37): 46–52. дои : 10.1080/14786443508648654 . Из стр. 48: «…смею предположить, что, поскольку огромная масса рассматриваемых пород, простирающаяся с юго-запада на северо-восток, пересекает царство наших предков силуров, следует принять термин «силурийская система»…»
- Уилмарт, Мэри Грейс (1925). Бюллетень 769: Классификация геологического времени Геологической службы США по сравнению с другими классификациями, сопровождаемая оригинальными определениями терминов эпохи, периода и эпохи . Вашингтон, округ Колумбия, США: Типография правительства США. п. 80.
- ^ Мерчисон, Родерик (1835). «О силурийской системе пород» . Лондонский и Эдинбургский философский журнал и научный журнал . Третья серия, Том. 7: 46–52 – через Библиотеку наследия биоразнообразия.
- ^ Седжвик; Мерчисон, Род-Айленд (1835 г.). «О силурийских и кембрийских системах, демонстрирующих порядок, в котором более древние осадочные пласты сменяют друг друга в Англии и Уэльсе» . Отчет пятого собрания Британской ассоциации содействия развитию науки. § Уведомления и рефераты различных сообщений в секции . 5 : 59–61.
- ^ Лэпворт, Чарльз (1879). «О тройственной классификации пород нижнего палеозоя» . Геологический журнал . 2-я серия. 6 (1): 1–15. Бибкод : 1879GeoM....6....1L . дои : 10.1017/s0016756800156560 . S2CID 129165105 . Со стр. 13–14: «Сам Северный Уэльс – во всяком случае, весь большой район Бала, где Седжвик впервые установил физическую последовательность между породами промежуточной или так называемой верхнекембрийской или нижнесилурийской системы; и во всех Вероятно, большая часть Шельфа и района Карадок, откуда Мерчисон впервые опубликовал свои отличительные окаменелости, находится на территории Ордовиков… Таким образом, здесь мы имеем намек на подходящее название для центральной системы нижнего палеозоя. называться Ордовикской системой, в честь этого древнего британского племени».
- ^ Готландская система была предложена в 1893 году французским геологом Альбертом Огюстом Кошоном де Лаппарентом (1839–1908): Лаппарент, А. де (1893). Трактат по геологии (на французском языке). Полет. 2 (3-е изд.). Париж, Франция: Ф. Сави. п. 748. Со с. 748: «В соответствии с этими подразделениями в силуре обычно выделяют три стадии: нижний или кембрийский ярус (1); средний или ордовикский ярус (2); верхний или готландский ярус (3)». (В соответствии с этими подразделениями в пределах силура обычно выделяют три этапа: нижний этап, или кембрий [1]; средний этап, или ордовик [2]; верхний этап, или готланд [3].)
- ^ Барранд, Иоахим (1852). Systême silurien du center de la Bohême (на французском языке). Париж, Франция и Прага (Чехия): (Собственное издание). стр. ix–x.
- ^ Форбс, Эдвард (1854). «Юбилейное послание президента» . Ежеквартальный журнал Лондонского геологического общества . 10 : XXII – LXXXI. См. стр. xxxiv.
- ^ Перейти обратно: а б Пол Селден и Хелен Рид (2008). «Самые старые наземные животные: силурийские многоножки из Шотландии» (PDF) . Бюллетень британской группы Myriapod & Isopod . 23 : 36–37.
- ^ Манда, Штепан; Фрида, Иржи (2010). «Пограничные события силура и девона и их влияние на эволюцию головоногих моллюсков: эволюционное значение размера яиц головоногих моллюсков во время массового вымирания» . Бюллетень геонаук . 85 (3): 513–40. дои : 10.3140/bull.geosci.1174 .
- ^ Перейти обратно: а б с д и ж г Муннеке, Аксель; Калнер, Майкл; Харпер, Дэвид А.Т .; Серве, Томас (2010). «Химический состав морской воды ордовика и силура, уровень моря и климат: краткий обзор». Палеогеография, Палеоклиматология, Палеоэкология . 296 (3–4): 389–413. Бибкод : 2010PPP...296..389M . дои : 10.1016/j.palaeo.2010.08.001 .
- ^ Ян, Гуанчжоу; Ленерт, Оливер; Мянник, Пип; Калнер, Микаэль; Луань, Сяокун; Гонг, Фаньи; Ли, Ликсия; Вэй, Синь; Ван, Гуансюй; Чжан, Ренбин; У, Жунчан (15 ноября 2022 г.). «Записи об изменениях климата в раннем силурийском периоде в Южном Китае и Балтии на основе комплексной биостратиграфии конодонтов и изотопной хемостратиграфии» . Палеогеография, Палеоклиматология, Палеоэкология . 606 : 111245. Бибкод : 2022PPP...60611245Y . дои : 10.1016/j.palaeo.2022.111245 . S2CID 252504361 . Проверено 8 декабря 2022 г.
- ^ Перейти обратно: а б Гамбакорта, Г.; Меникетти, Э.; Тринчианти, Э.; Торричелли, С. (март 2019 г.). «Силурийский климатический переход, зафиксированный в эпиконтинентальном Балтийском море». Палеогеография, Палеоклиматология, Палеоэкология . 517 : 16–29. Бибкод : 2019PPP...517...16G . дои : 10.1016/j.palaeo.2018.12.016 . S2CID 135118794 .
- ^ Янг, Сет А.; Бенаюн, Эмили; Козик, Невин П.; Подсказки, Олле; Мартма, Тыну; Бергстрем, Стиг М.; Оуэнс, Джереми Д. (15 сентября 2020 г.). «Морская окислительно-восстановительная изменчивость на Балтике во время вымираний в последнем ордовике – раннем силуре» . Палеогеография, Палеоклиматология, Палеоэкология . 554 : 109792. Бибкод : 2020PPP...55409792Y . дои : 10.1016/j.palaeo.2020.109792 . S2CID 218930512 .
- ^ Спросон, Адам Д. (15 февраля 2020 г.). «Темпы последнего ордовикского и силурийского углеродного цикла составляют орбитальный цикл ~ 4,5 млн лет» . Палеогеография, Палеоклиматология, Палеоэкология . 540 : 109543. Бибкод : 2020PPP...54009543S . дои : 10.1016/j.palaeo.2019.109543 . S2CID 213445668 . Проверено 8 декабря 2022 г.
- ^ Перейти обратно: а б Троттер, Джули А.; Уильямс, Ян С.; Барнс, Кристофер Р.; Мянник, Пип; Симпсон, Эндрю (февраль 2016 г.). «Новые записи конодонта δ18O об изменении климата в силурианском периоде: последствия для экологических и биологических событий». Палеогеография, Палеоклиматология, Палеоэкология . 443 : 34–48. Бибкод : 2016PPP...443...34T . дои : 10.1016/j.palaeo.2015.11.011 .
- ^ Чалонер, Уильям Г. (2003). «Роль углекислого газа в эволюции растений». Эволюция на планете Земля : 65–83. дои : 10.1016/B978-012598655-7/50032-X . ISBN 9780125986557 .
- ^ Нилон, Т.; Уильямс, Д. Майкл (30 апреля 2007 г.). «Шельфовые отложения силура Западной Ирландии, подвергшиеся воздействию штормов: новая интерпретация отложений глубокого бассейна». Геологический журнал . 23 (4): 311–320. дои : 10.1002/gj.3350230403 .
- ^ Самтлебен, К.; Муннеке, А.; Бикерт, Т. (2000). «Развитие фаций и изотопов C / O на разрезах через Ладлоу на Готланде: свидетельства глобального и локального влияния на мелководную морскую среду». Фации . 43 (1): 1–38. Бибкод : 2000Faci...43....1S . дои : 10.1007/BF02536983 . S2CID 130640332 .
- ^ Калнер, Микаэль (2008). «Силурийские глобальные события – переломный момент изменения климата». Массовое вымирание . стр. 21–57. дои : 10.1007/978-3-540-75916-4_4 . ISBN 978-3-540-75915-7 .
- ^ Расмуссен, Кристиан М. О.; Крёгер, Бьёрн; Нильсен, Мортен Л.; Кольменар, Хорхе (9 апреля 2019 г.). «Каскадный тренд раннепалеозойской морской радиации, приостановленный позднеордовикским вымиранием» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (15): 7207–7213. Бибкод : 2019PNAS..116.7207R . дои : 10.1073/pnas.1821123116 . ПМК 6462056 . ПМИД 30910963 .
- ^ Реталлак, Грегори Дж. (июнь 2022 г.). «Ордовик-девонские лишайниковые полога до появления древесных деревьев» . Исследования Гондваны . 106 : 211–223. Бибкод : 2022GondR.106..211R . дои : 10.1016/j.gr.2022.01.010 . S2CID 246320087 . Проверено 23 ноября 2022 г.
- ^ Риттнер, Дон (2009). Энциклопедия биологии . Издательство информационной базы. п. 338. ИСБН 9781438109992 .
- ^ Рикардс, РБ (1 марта 2000 г.). «Возраст самых ранних плаунов: силурийская флора барагванатии в Виктории, Австралия» . Геологический журнал . 137 (2): 207–209. Бибкод : 2000GeoM..137..207R . дои : 10.1017/S0016756800003800 . S2CID 131287538 . Проверено 8 декабря 2022 г.
- ^ Ланг, Вашингтон; Куксон, IC (1935). «О флоре, включая сосудистые наземные растения, связанные с Monograptus , в породах силурийского возраста из Виктории, Австралия» . Философские труды Лондонского королевского общества Б. 224 (517): 421–449. Бибкод : 1935РСТБ.224..421Л . дои : 10.1098/rstb.1935.0004 .
- ^ Хубер, FM (1983). «Новый вид Baragwanathia из секстантской формации (Эмс) Северного Онтарио, Канада». Ботанический журнал Линнеевского общества . 86 (1–2): 57–79. дои : 10.1111/j.1095-8339.1983.tb00717.x .
- ^ Бора, Лили (2010). Принципы палеоботаники . Публикации Миттала. стр. 36–37.
- ^ Перейти обратно: а б Никлас, Карл Дж. (1976). «Химические исследования некоторых несосудистых палеозойских растений». Бриттония . 28 (1): 113–137. Бибкод : 1976Britt..28..113N . дои : 10.2307/2805564 . JSTOR 2805564 . S2CID 21794174 .
- ^ Эдвардс, Д. и Веллман, К. (2001), «Эмбриофиты на суше: от ордовика до лочковского (нижнего девона) периода», в Гензеле, П. и Эдвардсе, Д. (ред.), Растения вторгаются в землю: эволюционные и экологические перспективы , Нью-Йорк: издательство Колумбийского университета, стр. 3–28, ISBN. 978-0-231-11161-4 , с. 4
- ^ Перейти обратно: а б с Брукфилд, Мэн; Катлос, Э.Дж.; Суарес, ЮВ (03 октября 2021 г.). «Время расхождения многоножек различается в зависимости от молекулярных часов и ископаемых свидетельств: U / Pb возраст циркона самых ранних ископаемых отложений, содержащих многоножек, и их значение» . Историческая биология . 33 (10): 2014–2018. Бибкод : 2021HBio...33.2014B . дои : 10.1080/08912963.2020.1762593 . ISSN 0891-2963 . S2CID 238220137 .
- ^ Данлоп, Джейсон А. (1996). «Тригонотарбидный паукообразный из верхнего силура Шропшира» . Палеонтология . 39 : 605–614. ISSN 0031-0239 .
- ^ Шир, Уильям А.; Джерам, Эндрю Дж.; Селден, Пол (1998). Многоногие ноги (Arthropoda, Chilopoda, Scutigeromorpha) из силура и девона Британии и девона Северной Америки . Нью-Йорк, штат Нью-Йорк: Американский музей естественной истории.
- ^ Джерам, Эндрю Дж.; Селден, Пол А.; Эдвардс, Дайанна (1990). «Наземные животные силура: паукообразные и многоножки из Шропшира, Англия». Наука . 250 (4981): 658–61. Бибкод : 1990Sci...250..658J . дои : 10.1126/science.250.4981.658 . ПМИД 17810866 .
- ^ ДиМишель, Уильям А; Крюк, Роберт В. (1992). «Силурийский» . Беренсмейер, Анна К. (ред.). Наземные экосистемы во времени: эволюционная палеоэкология наземных растений и животных . Издательство Чикагского университета. стр. 207–10. ISBN 978-0-226-04155-1 .
- ^ Уилсон, Хизер М.; Андерсон, Лайалл И. (2004). «Морфология и систематика палеозойских многоножек (Diplopoda: Chilognatha: Archipolypoda) из Шотландии» . Журнал палеонтологии . 78 (1): 169–184. Бибкод : 2004JPal...78..169W . doi : 10.1666/0022-3360(2004)078<0169:MATOPM>2.0.CO;2 . ISSN 0022-3360 . S2CID 131201588 .
- ^ Веллман, Швейцария; Лопес, Г.; МакКеллар, З.; Хартли, А. (2023). «Возраст базального слоя Стоунхейвенской группы« Нижнего старого красного песчаника » в Шотландии: самое старое зарегистрированное наземное животное, дышащее воздухом, имеет силурийский возраст (поздний Венлок)» . Журнал Геологического общества . 181 . Геологическое общество Лондона. дои : 10.1144/jgs2023-138 . hdl : 2164/22754 . ISSN 0016-7649 .
- ^ Суарес, Стефани Э.; Брукфилд, Майкл Э.; Катлос, Элизабет Дж.; Штёкли, Дэниел Ф. (28 июня 2017 г.). «U-Pb возраст циркона самого старого зарегистрированного наземного животного, дышащего воздухом» . ПЛОС ОДИН . 12 (6): e0179262. Бибкод : 2017PLoSO..1279262S . дои : 10.1371/journal.pone.0179262 . ISSN 1932-6203 . ПМЦ 5489152 . ПМИД 28658320 .
- ^ Брукфилд, Мэн; Катлос, Э.Дж.; Гарза, Х. (07 июля 2024 г.). «Самая старая ассоциация растений-многоножек? Возраст, палеосреда и источники отложений силурийских озер в Керрере, Аргайл и Бьют, Шотландия» . Историческая биология : 1–13. дои : 10.1080/08912963.2024.2367554 . ISSN 0891-2963 .
- ^ Ронг, Цзя-ю; Шен, Шу-чжун (1 декабря 2002 г.). «Сравнительный анализ массового вымирания и выживания брахиопод в конце перми и в конце ордовика в Южном Китае» . Палеогеография, Палеоклиматология, Палеоэкология . 188 (1–2): 25–38. Бибкод : 2002PPP...188...25R . дои : 10.1016/S0031-0182(02)00507-2 . Проверено 16 апреля 2023 г.
- ^ Ронг, Цзяюй; Чжан, Ренбин (октябрь 1999 г.). «Основные источники восстановления брахиопод после массового вымирания в конце ордовика со специальными ссылками на прародителей» . Наука в Китае. Серия D: Науки о Земле . 42 (5): 553–560. Бибкод : 1999ScChD..42..553R . дои : 10.1007/BF02875250 . S2CID 129323463 . Проверено 16 апреля 2023 г.
- ^ Хуан, Бинг; Ронг, Цзяюй; Кокс, Л. Робин М. (1 февраля 2012 г.). «Глобальные палеобиогеографические закономерности брахиопод от выживания до восстановления после массового вымирания в конце ордовика» . Палеогеография, Палеоклиматология, Палеоэкология . 317–318: 196–205. Бибкод : 2012PPP...317..196H . дои : 10.1016/j.palaeo.2012.01.009 . Проверено 16 апреля 2023 г.
- ^ Перейти обратно: а б Хуан, Бинг; Джин, Джисуо; Ронг, Цзя-Ю (15 марта 2018 г.). «Схемы диверсификации брахиопод после вымирания в раннем – среднем Лландовери, силурий» . Палеогеография, Палеоклиматология, Палеоэкология . 493 : 11–19. Бибкод : 2018PPP...493...11H . дои : 10.1016/j.palaeo.2017.12.025 . Проверено 23 ноября 2022 г.
- ^ Цзя-Ю, Ронг; Харпер, Дэвид А.Т. (31 января 2000 г.). «Выживание и восстановление брахиопод после последнего массового вымирания ордовика в Южном Китае» . Геологический журнал . 34 (4): 321–348. doi : 10.1002/(SICI)1099-1034(199911/12)34:4<321::AID-GJ809>3.0.CO;2-I . Проверено 16 апреля 2023 г.
- ^ Ансти, Роберт Л.; Пачут, Джозеф Ф.; Таки, Майкл Э. (8 апреля 2016 г.). «Особенности эндемизма мшанок в период ордовикско-силурийского перехода» . Палеобиология . 29 (3): 305–328. doi : 10.1666/0094-8373(2003)0292.0.CO;2 (неактивен 31 января 2024 г.) . Проверено 16 апреля 2023 г.
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на январь 2024 г. ( ссылка ) - ^ Запальский, Николай К.; Винн, Олев; Тоом, Урсула; Эрнст, Андрей; Уилсон, Марк А. (16 сентября 2022 г.). «Мшано-книдарийский мутуализм вызвал новую стратегию большей эксплуатации ресурсов еще в позднем силуре» . Научные отчеты . 12 (1): 15556. Бибкод : 2022NatSR..1215556Z . дои : 10.1038/s41598-022-19955-2 . ПМЦ 9481587 . ПМИД 36114227 .
- ^ Клауссен, Анна Лене; Муннеке, Аксель; Эрнст, Андрей (2 марта 2021 г.). «Богатые мшанками строматолиты (бриостроматолиты) из силура Готланда и их связь с климатическими возмущениями глобального углеродного цикла» . Седиментология . 69 (1): 162–198. дои : 10.1111/сед.12863 . S2CID 233820331 .
- ^ Лильедал, Луи (январь 1985 г.). «Экологические аспекты окремненной фауны двустворчатых моллюсков силура Готланда » Летайя 18 (1): 53–66. Бибкод : 1985Letter..18...53L . дои : 10.1111/j.1502-3931.1985.tb00684.x . Получено 16 апреля.
- ^ Клауссен, Анна Лене; Муннеке, Аксель; Уилсон, Марк А.; Освальд, Ирина (22 апреля 2019 г.). «Самые древние глубокосверлящие двустворчатые моллюски? Свидетельства из силура Готланда (Швеция)» . Фации . 65 (3): 26. Бибкод : 2019Faci...65...26C . дои : 10.1007/s10347-019-0570-7 . S2CID 149500698 . Проверено 16 апреля 2023 г.
- ^ Чернс, Лесли (2 января 2007 г.). «Раннепалеозойская диверсификация хитонов (Polyplacophora, Mollusca) на основе новых данных из силура Готланда, Швеция» . Летайя . 37 (4): 445–456. дои : 10.1080/00241160410002180 . Проверено 16 апреля 2023 г.
- ^ Перейти обратно: а б Винн, Олев; Уилсон, Марк А.; Мэдисон, Анна; Казанцева, Елизавета; Тоом, Урсула (26 июля 2022 г.). «Первая симбиотическая ассоциация гедереллоидов и ругозовых кораллов (последний силурийский период Сааремаа, Эстония)» . ПАЛЕОС . 37 (7): 368–373. Бибкод : 2022Палай..37..368В . дои : 10.2110/пало.2022.005 . S2CID 251122025 . Проверено 16 апреля 2023 г.
- ^ Винн, Олев (1 июня 2010 г.). «Адаптивные стратегии в эволюции корковых тентакулитоидных трубчатых червей» . Палеогеография, Палеоклиматология, Палеоэкология . 292 (1–2): 211–221. Бибкод : 2010PPP...292..211В . дои : 10.1016/j.palaeo.2010.03.046 . Проверено 16 апреля 2023 г.
- ^ Виттмер, Жакалин М.; Миллер, Арнольд И. (1 декабря 2011 г.). «Анализ траектории глобального разнообразия загадочной группы: палеогеографическая история тентакулитоидов» . Палеогеография, Палеоклиматология, Палеоэкология . 312 (1–2): 54–65. Бибкод : 2011PPP...312...54W . дои : 10.1016/j.palaeo.2011.09.009 . Проверено 16 апреля 2023 г.
- ^ Вэй, Фань; Гун, Имин; Ян, Хао (1 ноября 2012 г.). «Биогеография, экология и вымирание силурийских и девонских тентакулитоидов» . Палеогеография, Палеоклиматология, Палеоэкология . 358–360: 40–50. Бибкод : 2012PPP...358...40W . дои : 10.1016/j.palaeo.2012.07.019 . Проверено 16 апреля 2023 г.
- ^ Вэй, Синь; Жан, Рен-Бин (март 2018 г.). «Поздняя рудданская (ранняя лландоверская, силурийская) ассоциация трилобитов из Южного Китая и ее последствия» . Палеомир . 27 (1): 42–52. дои : 10.1016/j.palwor.2017.05.008 . Проверено 16 апреля 2023 г.
- ^ Адрайн, Джонатан М.; Вестроп, Стивен Р.; Чаттертон, Брайан Д.Э.; Рамшельд, Ларс (8 февраля 2016 г.). «Силурийское альфа-разнообразие трилобитов и массовое вымирание в конце ордовика» . Палеобиология . 26 (4): 625–646. doi : 10.1666/0094-8373(2000)0262.0.CO;2 (неактивен 31 января 2024 г.) . Проверено 16 апреля 2023 г.
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на январь 2024 г. ( ссылка ) - ^ Питерс, Шанан Э.; Аусич, Уильям И. (8 апреля 2016 г.). «Макроэволюционная история ордовикско-раннесилурийских криноидей с поправкой на выборку» . Палеобиология . 34 (1): 104–116. дои : 10.1666/07035.1 . S2CID 67838459 . Проверено 16 апреля 2023 г.
- ^ Райт, Дэвид Ф.; Тоом, Урсула (30 августа 2017 г.). «Новые криноидеи из Балтийского региона (Эстония): филогенетика датирования окаменелостей ограничивает происхождение и ордовикско-силурийскую диверсификацию Flexibilia (Echinodermata)» . Палеонтология . 60 (6): 893–910. Бибкод : 2017Palgy..60..893W . дои : 10.1111/пала.12324 . S2CID 134049045 .
- ^ Аусич, Уильям И.; Делайн, Брэдли (15 ноября 2012 г.). «Макроэволюционный переход криноидей после вымирания в позднем ордовике (от ордовика к раннему силуру)» . Палеогеография, Палеоклиматология, Палеоэкология . 361–362: 38–48. Бибкод : 2012PPP...361...38A . дои : 10.1016/j.palaeo.2012.07.022 . Проверено 16 апреля 2023 г.
- ^ Цзун, Жуй-вэнь; Гонг, И-мин (1 ноября 2020 г.). «Обнаружение сцифокриноидных лоболитов в западной Джунгарии, Синьцзяне, северо-западном Китае: значение для палеобиогеографии сцифокриноидов и идентификация границы силура и девона» . Палеогеография, Палеоклиматология, Палеоэкология . 557 : 109914. Бибкод : 2020PPP...55709914Z . дои : 10.1016/j.palaeo.2020.109914 . S2CID 224964119 . Проверено 16 апреля 2023 г.
- ^ Винн, О.; Уилсон, Массачусетс; Мытус, М.-А. (2014). «Симбиотические эндобионтные биофации силура Балтики» . Палеогеография, Палеоклиматология, Палеоэкология . 404 : 24–29. Бибкод : 2014PPP...404...24В . дои : 10.1016/j.palaeo.2014.03.041 . Проверено 11 июня 2014 г.
- ^ Винн, О.; Мытус, М.-А. (2008). «Самые ранние эндосимбиотические минерализованные трубчатые черви из силура Подолья, Украина» . Журнал палеонтологии . 82 (2): 409–414. Бибкод : 2008JPal...82..409V . дои : 10.1666/07-056.1 . S2CID 131651974 . Проверено 11 июня 2014 г.
- ^ Затонь, Михал; Врзолек, Томаш (15 октября 2020 г.). «Колонизация складчатых кораллов различными эпибионтами: доминирование и инкрустация Syn vivo в среде мягкого дна среднего девона (живетского периода) гор Святого Креста, Польша» . Палеогеография, Палеоклиматология, Палеоэкология . 556 : 109899. Бибкод : 2020PPP...55609899Z . дои : 10.1016/j.palaeo.2020.109899 . S2CID 224869051 . Проверено 16 апреля 2023 г.
- ^ Запальский, Николай К. (22 января 2014 г.). «Свидетельства фотосимбиоза в палеозойских таблитчатых кораллах» . Труды Королевского общества B: Биологические науки . 281 (1775): 20132663. doi : 10.1098/rspb.2013.2663 . ISSN 0962-8452 . ПМЦ 3866410 . ПМИД 24307674 .
- ^ Андреев, Пламен С.; Сансом, Иван Дж.; Ли, Цян; Чжао, Вэньцзинь; Ван, Цзяньхуа; Ван, Чун-Чье; Пэн, Лицзянь; Цзя, Ляньтао; Цяо, Туо; Чжу, Мин (сентябрь 2022 г.). «Самый древний гнатостомный зуб» . Природа . 609 (7929): 964–968. Бибкод : 2022Nature.609..964A . дои : 10.1038/s41586-022-05166-2 . ISSN 1476-4687 . ПМИД 36171375 . S2CID 252569771 .
- ^ Перейти обратно: а б Чжу, Ю-ан; Лу, Цзин; Ван, Цзяньхуа, Чжао, Вэньцзинь; Ю, Альберг, Чжу, Мин (2022). . раннего силура Китая» . Nature «Самые древние челюстные позвоночные 609 ): 954–958. : 2022Natur.609..954Z doi . : 10.1038 / Bibcode s41586-022-05136-8 ( 7929 4687. PMID 36171378. . S2CID 252569910 .
Ссылки
[ редактировать ]- Эмилиани, Чезаре . (1992). Планета Земля: космология, геология, эволюция жизни и окружающей среды . Издательство Кембриджского университета. (Издание в мягкой обложке ISBN 0-521-40949-7 )
- Микулич, Д.Г., Д.Г. Бриггс и Дж. Клюссендорф. 1985. Новая исключительно сохранившаяся биота из нижнего силура штата Висконсин, США. Философские труды Лондонского королевского общества, 311B:75-86.
- Мур, РА; Бриггс, DEG; Брэдди, С.Дж.; Андерсон, Л.И.; Микулич, Д.Г.; Клюссендорф, Дж (2005). «Новый синзифосурин (Chelicerata: Xiphosura) из позднего лландовери (силурийского периода) Уокеша Лагерштатт, Висконсин, США». Журнал палеонтологии . 79 (2): 242–250. Бибкод : 2005JPal...79..242M . doi : 10.1666/0022-3360(2005)079<0242:anscxf>2.0.co;2 . S2CID 56570105 .
Внешние ссылки
[ редактировать ]- Огг, Джим (июнь 2004 г.), Обзор разрезов и точек стратотипов глобальной границы (GSSP)
- Палеос: силурийский
- UCMP Беркли: силурианец
- Палеопортал: силурийские пласты в США, штат за штатом. Архивировано 15 марта 2021 г. в Wayback Machine.
- Геологическая служба США: силурийские и девонские породы (США).
- «Международная комиссия по стратиграфии (ICS)» . Геологическая шкала времени 2004 г. Проверено 19 сентября 2005 г.
- Примеры силурийских окаменелостей
- База данных GeoWhen для силура
- Силурий (хроностратографическая шкала)